
Om Prakash
Department of Biochemistry, Faculty of Sciences, Banaras Hindu University, Varanasi-221005, U.P., India
Nivedita Jaiswal
Department of Biochemistry, Faculty of Sciences, Banaras Hindu University, Varanasi-221005, U.P., India
Rajesh Kumar Pandey
Department of Biochemistry, Faculty of Sciences, Banaras Hindu University, Varanasi-221005, U.P., India
Asian Journal of Biochemistry
Year: 2011 | Volume: 6 | Issue: 3 | Page No.: 282-290







ABSTRACT
Effect of some metal ions, EDTA and sulfhydryl reagents on the activity of partially purified amylase (Sp. Activity 1213 U mg-1 protein) of soybean seeds were studied. Cobalt (II) and Manganese (II) exhibited marked activating effects on the activity, enhancing up to 200% of the initial activity at 2 mM concentration while Mercury (II) ions severely inhibited. Inhibition by mercury and activation by cobalt and manganese were concentration-dependent. However, other metal ions (K+, Ca2+, Mg2+, Al3+, Cu2+, Zn2+ and Fe3+) moderately increased the enzyme activity to a certain extent and then suppressed. Sodium, Cadmium and Nickel had no detectable influence on the activity. EDTA (12.5 mM) was found to be ineffective even for 1 h of incubation at 27°C suggesting that no metal ion is present in the enzyme. No marked inhibition of amylase activity with the sulfhydryl reagents was found. The stability of the enzyme to metal ions as well as EDTA suggests its promising potential for use in detergent industries.
PDF Abstract XML References Citation
Received: January 13, 2011;
Accepted: February 28, 2011;
Published: May 07, 2011
How to cite this article
Om Prakash, Nivedita Jaiswal and Rajesh Kumar Pandey, 2011. Effect of Metal Ions, EDTA and Sulfhydryl Reagents on Soybean Amylase Activity. Asian Journal of Biochemistry, 6: 282-290.
DOI: 10.3923/ajb.2011.282.290
URL: https://scialert.net/abstract/?doi=ajb.2011.282.290
DOI: 10.3923/ajb.2011.282.290
URL: https://scialert.net/abstract/?doi=ajb.2011.282.290
INTRODUCTION
Amylases are among the most important enzymes used for several biotechnological applications particularly employed in starch processing industries for the hydrolysis of polysaccharides such as starch into simpler sugar constituents (Ikram-ul-Haq, et al., 2002) and this is the basis for various industrial processes like preparation of glucose syrups, bread making, brewing, etc. These enzymes account for about 30% of the world’s enzyme production (Van der Maarel et al., 2002). Besides, it also finds use in detergents, desizing of textiles, modified starches, hydrolysis of oil-field drilling fluids and paper recycling (Richardson et al., 2002).
Many substances alter the activity of an enzyme by combining it in a way that influences the binding of the substrate (Dogan et al., 2007). These substances can be called as effectors either being activators or inhibitors. Metal ions can be considered as good examples, different metals exhibiting different behaviors in their ability to act as effectors. Li et al. (2005) also stated that metal binding to enzymes plays an important role in their activation and stabilization. Most amylases are known to be metal ion dependent enzymes (Pandey et al., 2000; Gupta et al., 2003; Ramachandran et al., 2004). According to Leveque et al. (2000), a metal ion present at high concentrations might compete with another metal present at a lower concentration and replace it at a metal-binding site, even if its affinity for the binding site is lower.
An accepted property of amylases is their content of Ca2+ as an integral part of the enzyme molecule and their consequent activation by Ca2+ and inhibition by chelating reagents (Mar et al., 2003; Kiran and Chandra, 2008). Besides, amylases have been reported to be inhibited by heavy metal ions such as Ag+, Hg2+, Cu2+, Fe2+, etc., suggesting the involvement of cysteine residue in enzyme activity. The importance of cysteine residues for catalysis has been described for amylases independent of their origin (Pandey et al., 2000; Lo et al., 2001; Diaz et al., 2003). The thiol of a cysteine residue occasionally acts as a ligand of metal ions (Tatara et al., 2005). Amylases are also found sensitive to various sulfhydryl modifying reagents as free thiol or sulfhydryl groups are considered to be useful in stability, redox behaviour, metal binding, acidity, nucleophilicity and catalytic activity (Bardwell, 2005; Giles et al., 2003).
In order to obtain an insight into these, an attempt has been made to partially purify amylase of soybean seeds and the effect of various metal ions and the chelating reagent; EDTA on the amylase activity was determined. Besides, the possible effects of sulfhydryl reagents (p-hydroxymercurybnzoic acid, N-ethylmaleimide and iodoacetic acid) on amylase activity have also been investigated.
MATERIALS AND METHODS
Materials: Soybean seeds were procured from the local market. Sephadex G-75 was from Pharmacia Fine Chemicals Uppsala, Sweden. Bovine serum albumin and maltose were obtained from Sigma Chemical Co., USA. Soluble starch and ammonium sulphate were from Qualigens Fine Chemicals, Mumbai. All other reagents were analytical grade chemicals either from BDH or E. Merck, India. The study was conducted during January 2009 to December 2009.
Enzyme extraction and preparation of crude extract: Soybean seeds (50 g) were soaked in 25 mM sodium acetate buffer (pH 5.5) for 8 h at 4°C. The soaked seeds were homogenized along with the buffer using a kitchen blender, filtered through muslin cloth and centrifuged at 15,000 rpm (21,633 g) for 30 min. The supernatant so obtained (crude preparation) was subjected to further purification.
Enzyme and protein assays: The amylase activity was determined by the method of Bernfeld (1955) with slight modifications. The assay system contained 1.0 mL of suitably diluted enzyme solution, 0.5 mL of 2% soluble starch (in assay buffer) and 0.5 mL of assay buffer (Tris acetate, 150 mM, pH 5.5). This reaction mixture was incubated at 27°C for 3 min. The reaction was terminated by the addition of 2.0 mL of DNS (3,5-dinitrosalicylic acid) reagent solution. Colour, due to the reducing sugar liberated was developed by heating the reactants in a boiling water bath for 5 min and then cooling down to room temperature. After addition of 20 mL of double distilled water, the absorbance was determined at 540 nm. A blank was prepared as above but without the enzyme. A calibration curve of maltose was also prepared.
One unit of amylase activity was defined as the amount of enzyme required to produce 1 μmol of maltose per minute under the assay conditions.
The alkaline 3,5-dinitrosalicylic acid reagent occasionally gives anomalous results, due to the effect of various ions on the reducing value (Robyt and Whelan, 1965).
Protein was estimated by the method of Lowry et al. (1951) with Folin-ciocalteau reagent calibrated with crystalline bovine serum albumin.
Enzyme purification
Ammonium sulphate precipitation: The crude amylase preparation (50 mL) was precipitated with solid ammonium sulphate. The sample was separated into three fractions based on the saturation percent of ammonium sulphate (0-30, 30-80 and 80-100%). The precipitation was carried out at 4°C under constant stirring and the precipitated proteins were centrifuged at 10,000 rpm (9615 g) for 15 min. The precipitate obtained was dissolved in the extraction buffer (25 mM sodium acetate, pH 5.5). Specific activity of the sample was estimated in all the three fractions. The selected fraction having high specific activity was further processed by dialysis (against the same extraction buffer) and centrifugation (9615 g, 15 min) prior to gel filtration chromatography.
Sephadex G-75 gel filtration chromatography: The enzyme obtained from the previous step was applied (1.0 mL) to a Sephadex G-75 column (1.4x32 cm) pre-equilibrated with 25 mM sodium acetate buffer (pH 5.5) and eluted with the same buffer. The eluate collected in various 2 mL fractions were analyzed for both protein and amylase activity. The active fractions were pooled and concentrated against solid sucrose. The enzyme preparation was kept frozen (at-20°C) in small aliquots until further use.
Electrophoresis: 12% Native polyacrylamide gel electrophoresis was carried out according to the method of Laemmli (1970). The bands were visualized by coomassie blue and silver staining in order to check the purity of the enzyme preparation.
Effect of metal ions, EDTA and sulfhydryl reagents on amylase activity: A stock solution of each of the desired reagent was prepared in double distilled water. The residual amylase activity of suitably diluted enzyme was determined in the presence of varying concentration of these reagents added in the standard assay mixture (comprising of 2% starch, 150 mM Tris acetate buffer, pH 5.5 at 27°C). In order to explore the stability of the enzyme to the chelating reagent EDTA, enzyme was incubated with various concentrations of EDTA (direct exposure) for 1 h at 27°C at pH 5.5 and then the residual activity of the exposed enzyme was determined.
RESULTS AND DISCUSSION
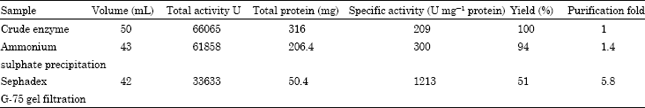
Partial purification of the enzyme: Among the three fractions selected for the study, protein precipitated in the range of 30-80% saturation of ammonium sulphate (having a specific activity of 300 U mg-1 protein) was selected for further separation on Sephadex G-75 column. Taking into account the amylase obtained in a single symmetrical peak from fractions 8-22, 5.8 fold purification was achieved with recovery of 51% (Table 1 and Fig. 1). Recently, Mohamed et al. (2009) partially purified amylase from a wheat local variety with 1.41 fold purification and 2.3% recovery.
Effect of metal ions on soybean amylase activity: Various metal ions such as K+, Ca2+, Mg2+, Al3+, Cu2+, Zn2+, Fe3+, Na+, Cd2+, Ni2+, Hg2+, Pb2+, CO2+, Mn2+ at various concentrations (ranging from 0.0005 to 5 mM) were tested for activation/inhibition effects.
| Table 1: | Partial purification of α-amylase from soybean seeds |
 | |
 | |
| Fig. 1: | Purification profile of soybean amylase in Sephadex G-75 gel filtration chromatography |
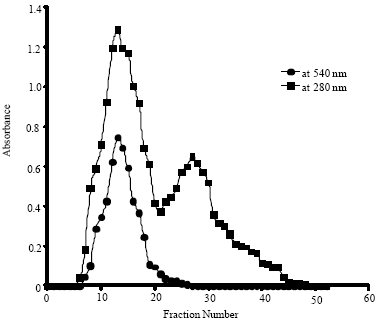
Na+, Cd2+ and Ni2+ had negligible effect on amylase activity (data not shown). K+, Mg2+, Al3+ and Fe3+ showed an increase in amylase activity upto 0.005 mM and then showed a gradual decline with increasing concentrations (Fig. 2). Similarly, the activating effect of Ca2+, Cu2+ and Zn2+ on amylase was dependent on the concentration of the metals upto the limiting value of 0.25 mM, beyond which there was a decrease in activity. Usually, the role of Ca2+ in maintaining the stability and structure of the amylase is well documented (Parkin, 1993). Ca2+ was found to increase the activity of α-amylase from Bacillus subtilis (El-Banna et al., 2007). Cu2+ showed strong inhibition on the activity of the enzyme from Bacillus subtilis JS-2004 (Asgher et al., 2007). The effect of Zn2+ varied between amylases. For instance, it had a potent inhibitory effect on Schwanniomyces alluvius α-amylase (Moranelli et al., 1987).
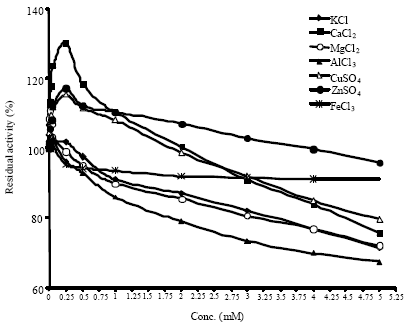
A concentration-dependent and time-independent inhibition in enzyme activity was observed in case of heavy metal ion Hg2+ (observed from 0.001 to 5 mM, Fig. 3). A similar trend with respect to concentration-dependence was observed in case of Pb2+ which showed complete loss in activity at 50 mM (data not shown). Hg2+, Pb2+ and Cu2+ have been reported to inhibit microbial amylase (Lin et al., 1998). Hg2+ was observed to be a potent inhibitor in α-amylase from mung beans (Tripathi et al., 2007). The inhibitory effect of Hg2+ on amylase may indicate the importance of indole amino acid residues in enzyme function as has been demonstrated for other microbial amylases (Gupta et al., 2003).
A significant stimulation in amylase activity was observed with increasing concentration of CO2+ and Mn2+ (observed from .0005 to 2 mM, Fig. 4) which are of wide occurrence in biological systems. Dahot et al. (2001) also found an increase in amylase activity with CO2+ and Mn2+ with Moringa oleifera seeds. Similarly, Dutta et al. (2006) obtained an enhanced amylase activity in Heliodiaptomus viduus with these ions (at 5 mM). In contrast, an inhibitory effect of CO2+ on amylase from B. subtilis JS-2004 was observed (Asgher et al., 2007). Penicillium olsonii α-amylase showed an enhanced enzyme activity with Mn2+ (1 mM) while no effects in enzyme activity was found with CO2+ (Afifi et al., 2008).
 | |
| Fig. 2: | Effect of metal ions on the activity of amylase of soybean seeds |
 | |
| Fig. 3: | Effect of the heavy metal ion Hg2+ on soybean amylase activity |
Reports have been made that metallic chlorides are potential activators of amylases (Vega-Villasante et al., 1993; Oboh and Ajele, 1997; Mohapatra et al., 1998). If this is the case, then the metallic chlorides used (viz. KCl, CaCl2, MgCl2 and FeCl3) should show activation virtually at all the concentrations tested. Moreover, if it were responsible alone, one would expect the degree of activation to be the same for all the metallic chlorides tested. However, results as already described are contrary to this. This argument is in line with the findings of Wakim et al. (1969) which showed that halides activate amylase activity but they are not mandatory for the activity of the enzyme. Florkin and Mason (1962) also stated that amylases are either activated by this anion or are indifferent to its presence. Thus, it can be said that the effects observed on the enzyme activity might be the contribution from cations.
 | |
| Fig. 4: | Effect of cobalt and manganese on soybean amylase activity |
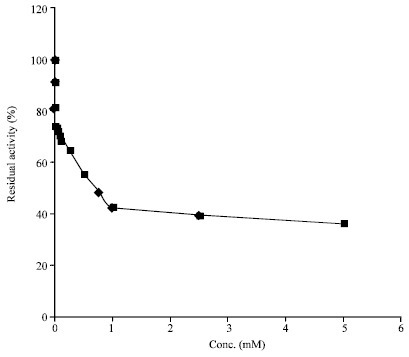
Effect of EDTA on soybean amylase activity: It is well known that amylases contain Ca2+ as an essential component of the enzyme molecule and are often inhibited by the chelating reagent EDTA (Mar et al., 2003; Kiran and Chandra, 2008) presumably because of its chelating properties. Interestingly, soybean amylase retained almost full activity with the varying concentrations of EDTA (from 0.0005 to 12.5 mM) used in the study. Besides, the enzyme maintained about 90% of its initial activity even after 1 h incubation with 12.5 mM EDTA (Fig. 5). Utong et al. (2006) also found continued production of extracellular amylase activity suggesting that the amylase enzyme produced was not a metalloenzyme. In contrast, Femi-Ola and Olowe (2011) found that EDTA inhibited the enzyme activity suggesting that EDTA acted by chelating Ca2+. These results indicate that soybean amylase is strongly resistant to the chelating reagent which is very interesting in that no metal ion is essential for the manifestation of soybean amylase activity. Moreover, its stability to incubation with chelating reagents (one of the indispensable ingredients in detergent formulations) is of great importance from the viewpoint of its high performance even in detergent formulations and the physiological significance of which calls for study in future works.
Effect of sulfhydryl reagents on soybean amylase activity: The effect of the sulfhydryl reagents (p-hydroxymercurybenzoic acid, N-ethylmaleimide and Iodoacetic acid; observed from 0.5 to 12.5 mM) on amylase activity are presented in Fig. 6. The minimum amount of these reagents inducing a detectable inhibition of activity in soybean amylase was found to be 1 mM. Iodoacetic acid and N-ethylmaleimide showed decline in enzyme activity with increasing concentrations while pHMB had only marginal inhibitory effects on the activity of soybean amylase. This pattern of sulfhydryl reagent selectivity is not like that reported for other plant amylases which are largely inhibited by low concentrations of any sulfhydryl reagent (Marshall, 1975). Besides, all sulfhydryl reagents used in this study should have totally inhibited activity if cysteinyl sulfhydryls were necessary for catalysis. However, the data indicate that soybean amylase sulfhydryls are not directly involved in catalysis. Soybean amylase inhibition by the sulfhydryl reagents could be due to binding of noncatalytic cysteinyl sulfhydryls, causing changes in the alignment of catalytic amino acids, as is proposed for sweet potato α-amylase (Thoma et al., 1971) or by causing steric hindrance.
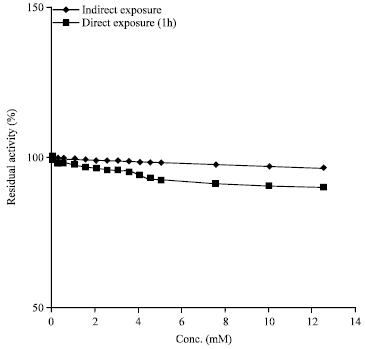 | |
| Fig. 5: | Effect of indirect exposure (in the presence of substrate) and direct exposure of soybean amylase (1 h) to EDTA |
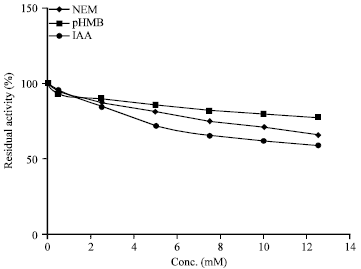 | |
| Fig. 6: | Effect of sulfhydryl reagents on activity of soybean amylase |
CONCLUSION
The soybean amylase was partially purified by ammonium sulphate fractionation and gel filtration chromatography with a purification factor of 5.8 and a recovery of 51%. The enzyme retained more than 65% activity even in the presence of metal ions at a concentration of 5 mM. It was also found quite stable to the effect of chelating reagent, EDTA. No significant inhibition with the sulfhydryl reagents was found indicating that sulfhydryls of soybean amylase are intact and not free to be directly involved in catalysis. The stability of the enzyme to the metal ions and chelating reagent establishes its potential for use in detergent formulations.
REFERENCES
- Ikram-ul-Haq, R. Abdullah, H. Ashraf and A.H. Shah, 2002. Isolation and screening of fungi for the biosynthesis of alpha amylase. Biotechnology, 2: 61-66.
CrossRefDirect Link - Van der Maarel, M.J.E.C., B. van der Veen, J.C.M. Uitdehaag, H. Leemhuis and L. Dijkhuizen, 2002. Properties and applications of starch-converting enzymes of the α-amylase family. J. Biotechnol., 94: 137-155.
CrossRefPubMedDirect Link - Richardson, T.H., X. Tan, G. Frey, W. Callen and M. Cabell et al., 2002. A novel, high performance enzyme for starch liquefaction: Discovery and optimization of a low pH, thermostable α-amylase. J. Biol. Chem., 277: 26501-26507.
CrossRefPubMedDirect Link - Dogan, S., P. Turan, M. Dogan, M. Alkan and O. Arslan, 2007. Inhibition kinetics of polyphenol oxidase by glutamic acid. Eur. Food Res. Technol., 225: 67-73.
CrossRef - Li, W.F., X.X. Zhou and P. Lu, 2005. Structural features of thermozymes. Biotechnol. Adv., 23: 271-281.
CrossRefPubMedDirect Link - Pandey, A., P. Nigam, C.R. Soccol, V.T. Soccol, D. Singh and R. Mohan, 2000. Advances in microbial amylases. Biotechnol. Applied Biochem., 31: 135-152.
PubMedDirect Link - Gupta, R., P. Gigras, H. Mohapatra, V.K. Goswami and B. Chauhan, 2003. Microbial α-amylases: A biotechnological perspective. Process Biochem., 38: 1599-1616.
CrossRefDirect Link - Ramachandran, S., A.K. Patel., K.M. Nampoothiri, S. Chandran, G. Szakacs, C.R. Soccol and A. Pandey, 2004. α-amylase from a fungal culture grown on oil cakes and its properties. Braz. Arch. Biol. Technol., 47: 309-317.
Direct Link - Mar, S.S., H. Mori, H.J. Lee, K. Fukuda and A. Kimura et al., 2003. Purification, characterization and sequence analysis of two alpha-amylase isoforms from azuki bean, Vigna angularis, showing different affinity towards beta-cyclodextrin sepharose. Biosci. Biotechnol. Biochem., 67: 1080-1093.
PubMed - Kiran, K.K. and T.S. Chandra, 2008. Production of surfactant and detergent-stable, halophilic and alkalitolerant alpha-amylase by a moderately halophilic Bacillus sp. Strain TSCVKK. Applied Microbiol. Biotechnol., 77: 1023-1031.
Direct Link - Femi-Ola, T.O. and B.M. Olowe, 2011. Characterization of α amylase from Bacillus subtilis BS5 isolated from Amitermes evuncifer silvestri. Res. J. Microbiol., 6: 140-146.
CrossRefDirect Link - Lo, H.F., L.L. Lin, H.L. Chen, H.H. Hsu and C.T. Chang, 2001. Enzymatic properties of a SDS-resistant Bacillus sp. TS-23 α-amylase produced by recombinant Escherichia coli. Process Biochem., 36: 743-750.
CrossRef - Diaz, A., C. Sieiro and T.G. Villa, 2003. Production and partial characterization of a β-amylase by Xanthophyllomyces dendrorhous. Lett. Applied Microbiol., 36: 203-207.
CrossRef - Tatara, Y., T. Yoshida and E. Ichishima, 2005. A single free cysteine residue and disulphide bond contribute to the thermostability of Aspergillus saitoi 1,2-α-omannosidase. Biosci. Biotechnol. Biochem., 69: 2101-2108.
Direct Link - Marshall, J.J., 1975. Inhibition of plant and bacterial β-amylases. Mol. Cell. Biochem., 7: 127-129.
CrossRef - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Mohamed, S.A., A.L. Al-Malki and T.A. Kumosani, 2009. Partial purification and characterization of five α-amylase from a wheat l0cal variety (Balady) during germination. Aust. J. Basic Applied Sci., 3: 1740-1748.
Direct Link - El-Banna, T.E., A.A. Abd-Aziz, M.I. Abou-Dobara and R.I. Ibrahim, 2007. Production and immobilization of α-amylase from Bacillus subtilis. Pak. J. Biol. Sci., 10: 2039-2047.
CrossRefPubMedDirect Link - Asgher, M., M.J. Asad, S.U. Rahman and R.L. Legge, 2007. A thermostable α-amylase from a moderately thermophilic Bacillus subtilis strain for starch processing. J. Food Eng., 79: 950-955.
CrossRef - Lin, L.L., C.C. Chyau and W.H. Hsu, 1998. Production and properties of a raw-starch-degrading amylase from the thermophilic and alkaliphilic Bacillus sp. TS-23. Biotechnol. Applied Biochem., 28: 61-68.
PubMedDirect Link - Tripathi, P., L.L. Leggio, J. Mansfeld, R. Ulbrich-Hofmann and A.M. Kayastha, 2007. α-Amylase from mung beans (Vigna radiata)-correlation of biochemical properties and tertiary structure by homology modelling. Phytochemistry, 68: 1623-1631.
CrossRef - Vega-Villasante, F., H. Nolasco and R. Civera, 1993. The digestive enzymes of the pacific brown shrimp Penaeus californiensis: I-properties of amylase activity in the digestive tract. Compre. Biochem. Physiol., 106: 547-550.
CrossRef - Mohapatra, M.A., U.C. Banerjee and M. Bapuji, 1998. Characterization of a fungal amylase from Mucor sp. associated with the marine sponge Spirastrella sp. J. Biotechnol., 60: 113-117.
CrossRefDirect Link - Leveque, E., B. Haye and A. Belarbi, 2000. Cloning and expression of an α-amylase encoding gene from the hyperthermophilic archaebacterium Thermococcus hydrothermalis and biochemical characterization of the recombinant enzyme. FEMS Microbiol. Lett., 186: 67-71.
PubMed - Utong, J., F. Al-Quadan and H. Akel, 2006. Effect of various growth conditions on production of extracellular amylase from thermotolerant Bacillus species isolated from hot springs in Jordan. J. Boil. Sci., 6: 621-625.
CrossRefDirect Link - Giles, N.M., G.I. Giles and C. Jacob, 2003. Multiple roles of cysteine residue in biocatalysis. Biochem. Biophys. Res. Commun., 300: 1-4.
Direct Link - Dutta, T.K., M. Jana, P.R. Pahari and T. Bhattacharya, 2006. The effect of temperature, pH and salt on amylase in Heliodiaptomus viduus (Gurney) (Crustacea: Copepoda: Calanoida). Turk. J. Zool., 30: 187-195.
Direct Link - Wakim, J., M. Robinson and J.A. Thoma, 1969. The active site of porcine-pancreatic alpha-amylase: Factors contributing to catalysis. Carbohydrate Res., 10: 487-503.
CrossRef - Dahot, M.U., A.A. Saboury, S. Ghobadi and A.A. Moosavi-Movahedi, 2001. Properties of the alpha amylase from Moringa oleifera seeds. J. Biol. Sci., 1: 747-749.
CrossRefDirect Link