
Gamal El-din I. Harisa
Department of Pharmaceutics, College of Pharmacy, Kayyali Chair for Pharmaceutical Industry, King Saud University, P.O. Box 2457, Riyadh 11451, Saudi Arabia
Asian Journal of Biochemistry
Year: 2011 | Volume: 6 | Issue: 3 | Page No.: 263-272







ABSTRACT
The present study was conducted to investigate the effect of L-Arginine (Arg) supplementation on protein oxidation as well as Paraoxonase (PON) activities in rats received either normal or high cholesterol diet with or without 3% Arg in drinking water. Hypercholesterolemia (HC) was induced in rats by feeding diet containing cholesterol 5%, cholic acid 1% and propylthiouracil 0.5%. The spectrophotometric analysis methods were used for determination of lipid profiles as well as oxidative stress biomarkers in the plasma and liver tissues of rats. The results of current study revealed that the feeding of rats with Hypercholesterolemic Diet (HCD) resulted in significant increased of Atherogenic Index (AI) by 256% in respect to control rats. Arg supplementation to rats with HCD resulted in decrease of AI by 152% comparison with rats received HCD. Moreover, administration of Arg with normal or HCD attenuates the protein oxidation as well as lipids peroxidation in compared to intake of HCD alone. As a result of HCD feeding, plasma arylesterase (ARE) activity was decreased by 30% while in liver it decreased by 36%. Furthermore, paraoxonase activity (PON1) was decreased 42 and 74% for plasma and liver, respectively. On the other hand, the treatment with Arg ameliorates both activities in the plasma and liver in comparison with HCD. Arg preserve thiols and nitric oxide level, thereby reducing oxidative stress associated with increased cholesterol level. This may be related antioxidant effect of this amino acid. Therefore, Arg has beneficial effects in the treatment of HC thruogh decreases the proteins oxidation as well as preservation of high density lipoprotein function through maintaining PON activity.
PDF Abstract XML References Citation
Received: February 14, 2011;
Accepted: March 05, 2011;
Published: May 07, 2011
How to cite this article
Gamal El-din I. Harisa, 2011. L-Arginine Ameliorates Arylesterase/Paraoxonase Activity of Paraoxonase-1 in Hypercholesterolemic Rats. Asian Journal of Biochemistry, 6: 263-272.
DOI: 10.3923/ajb.2011.263.272
URL: https://scialert.net/abstract/?doi=ajb.2011.263.272
DOI: 10.3923/ajb.2011.263.272
URL: https://scialert.net/abstract/?doi=ajb.2011.263.272
INTRODUCTION
Increase of cholesterol in the blood is associated with increase its level in the tissues leading to oxidative stress (Kumar et al., 2006). Under these conditions xanthine oxidase (OX) (E.C.1.2.3.2) uses oxygen molecules as electron acceptors to produce superoxide radicals beside uric acid (Asai et al., 2007). Superoxide can be convert Nitric Oxide (NO) to peroxynitrite that mediate nitration of sulfur and aromatic residues of amino acids in polypeptide chains (Dalle-Donne et al., 2005). Moreover, superoxide itself can react with cysteine residues of sulfur containing amino acids as well as it can be converted to H2O2 and hydroxyl radical. This radical can reacts with either aliphatic, sulfur containing or aromatic amino acid, leading to activation or inhibition of the protein functions (Schoneich and Sharov, 2006). Additionally, such radicals attack the polyunsaturated fatty acids in the cells membrane particularly erythrocytes and vascular endothelium leading to lipids peroxidation expressed as malondialdehyde (Marjani et al., 2007). The cellular defense against oxidative stress is carried out by enzymes system such as superoxide dismutase, catalase and glutathione peroxidase. Also, cellular thiols like protein thiol, non-protein thiols as well as other none enzymatic antioxidants systems (Cimen, 2008).
Paraoxonase is aryldialkylphosphatase (E.C. 3.1.8.1) as one of antioxidant enzymes which catalyze the breakdown of paraoxon, paraoxonase activity (PON) and arylesters, arylesterase activity (ARE). Moreover, this enzyme has important role in lipid metabolism (Draganov and La Du, 2004). Paraoxonase-1 (PON1) is present in the serum on High-Density Lipoprotein (HDL) and plays an important role in the degradation of lipid peroxides. Furthermore, PON protects Low-Density Lipoprotein (LDL) and HDL from oxidation induced by free radicals (Serdar et al., 2006). The paraoxonase is member of antioxidant enzyme systems plays an important role in the protection of proteins as well as lipids against oxidative damage (Shetty et al., 2008). PON activity may be inhibited by increase of free radicals production (Rozenberg et al., 2003).
NO is vasodilator; such molecule plays an important role in regulation of cardiovascular, nervous and immune systems functions (McGrowder and Brown, 2007). The production of NO is decrease in response to oxidative stress; this may be one of the possible causes of complications associated with dyslipidemia (Honing et al., 1998). This decrease may be due to either deficiency of L-Arginine (Arg) or a deficiency of some cofactors required for NO production by NO synthase (El-Missiry et al., 2004). Arg decreases the superoxide production as well as improve endothelial function by restoration of NO levels (Kawano et al., 2002). Moreover, Arg has protective effects against reactive oxygen metabolites induced injury through interaction with these radicals; therefore, Arg supplementation attenuates the oxidative stress (Tsai et al., 2002).
Arg is used in treatment of Hypercholesterolemia (HC) but up till now the exact mechanism is still unknown. Therefore, the objective of current study was to investigate the effect of Arg supplementation on PON1, ARE activities of PON enzyme as well as proteins oxidation in rats supplemented with diet induced HC.
MATERIAL AND METHODS
Chemicals and instruments: Paraoxon, phenylacetate, cholic acid, L-Arginine, 5,5'-Dithiobis (2-nitrobenzoic acid), thiobarbituric acid, tetraethoxypropane, guanidine-HCl and Tris HCl, were supplied from (Sigma Chemical Co., St. Louis, MO, USA). All of the remaining chemicals are of analytical grade. Beckman XL-70 ultracentrifuge, 100,000 rpm (USA), JENWAY spectrophotometer model 6105 UV/VIS. Centrifuge Sigma 3K 20, up to 10,000 g (Germany), Homogenizer: Janke and Kunkel IKA 8,000-20.500 RPM (Germany).
Animals: Male wistar albino rats were obtained from animal facility. The animals were housed in animal house unit, in the Department of Pharmacology, College of Pharmacy, Al-Azhar University. The animals were received standard diet and water under standardized conditions away from stress condition. The standard diet was composed of 72.2% carbohydrate, 3.4% fats, 19.8% proteins, 3.6% cellulose, 0.5% vitamins and minerals and 0.5% salts. The animals were housed in metalic cages under standard laboratory conditions (12 h light/dark cycles at 25±2°C) with free access to standard rat pellet food and water. The rats were left for 2 weeks for acclimatization.
Experimental design: This study was carried out on 24 male wistar albino rats weighing 180±210 g and rats were divided into 4 groups, 6 rats in each group. Group 1; normal control (C) the rats were fed on standard diet pellets. Group 2; rats were fed on standard diet pellets plus 3% Arg in drinking water (White et al., 2004). Group 3; rats were supplemented with HCD in which the dietary chow was supplemented with 4% cholesterol, 1% cholic acid and 0.5% propylthiouracil (Deepa and Varalakshmi, 2006). Group 4; rats were received Arg in drinking water 3% in addition to HCD. The experimental protocol was carried out for 2 months. Experiments were conducted according to the guidelines of institutional animal ethical committee of College of Pharmacy, Al-Azhar University, Nasr City and Cairo, Egypt.
Preparation of blood and tissues specimens: All rats were allowed to fast for 12 h at the end of experiment and then sacrificed under diethyl ether anesthesia. Blood samples were drown by cardiac puncture and collected into heparinized centrifuge tubes and left to stand at room temperature for 10 min, then centrifuged at 3000 rpm for 10 min. The isolated plasma samples were stored at-20°C until analysis. The homogenate of liver tissue was prepared based on the previously described method of Feillet-Coudray et al. (2009).
Determination of plasma lipids profile, liver functions and kidney functions tests: The plasma level of Total Cholesterol (TC), High Density Lipoprotein Cholesterol (HDL-C), Triacylglycerol (TAG) and Low Density Lipoprotein Cholesterol (LDL-C) were measured and used for Atherogenic Index (AI) calculation. Uric acid, urea, creatinine, Alanine Transaminase (ALT), Aspartate Transaminase (AST), Alkaline Phosphatase (ALP) were determined using a commercially available kit (Biocon Diagnostic, Germany). Hepatic lipids were assayed after extraction by using the method of Folch et al. (1957).
Assay of oxidative stress biomarkers: Protein oxidation was assayed by measuring of Protein Carbonyl (PCO) content according to the method of Levine et al. (1994). Total protein content was determined by the method of Lowry et al. (1951) and method of Hu (1994) was used for determination of total thiols (TSH) by using Ellman’s reagent (5,5'-Dithiobis (2-nitrobenzoic acid)). Furthermore, Malondialdehyde (MDA) was determined spectrophotometrically as indicator of lipid peroxidation by the method of Ohkawa et al. (1979). Plasma nitrate levels were determined by a colorimetric method based on the Griess reaction (Moshage et al., 1995).
Plasma paraoxonase-1/arylesterase assay: The activity of PON1 toward paraoxon was determined by measuring the initial rate of substrate hydrolysis to p-nitrophenol. While plasma arylesterase activity of PON1 is measured using phenylacetate as substrate (Ayub et al., 1999).
Paraoxonase-1/arylesterase activity in liver tissues: PON1 activity toward phenylacetate in supernatants was assayed by the same method as in plasma. Whereas, PON1 activity toward paraoxon was measured as following. Ten microliters of liver homogenate were mixed with 300 μL of the reaction buffer containing 50 mM Tris HCl (pH 8.0), 2 mM paraoxon then 2 mM CaCl2 and the increase in absorbance at 405 nm. Enzyme activity in tissues was expressed in U mg-1 of protein.
Statistical analysis: Statistically differences between the studied groups is determined using one-way analysis of variance (ANOVA) followed by the Tukey-Kramer post analysis test to compare all groups. The data were expressed as Mean±Standard Deviation (SD), p≤0.05 were considered significant. GraphPad Prism® was used for statistical calculations (Version 5.00 for Windows, GraphPad Software, San Diego California USA).
RESULTS
In the present study, the feeding of cholesterol rich diet caused marked increase of TC, TAG and LDL-C and atherogenic index while HDL-C is decrease as compared with the rats received normal dietary chow (Table 1). Moreover the feeding of cholesterol rich diet caused marked increased of liver content of total lipids, TC as well as TAG (Table 2). Also, such diet caused a moderate increase in the activities of ALT, AST and ALP in comparison with intake of normal diet or normal diet with Arg 3% (Table 1).
| Table 1: | Plasma level of TC, HDL-C, TAG, LDL-C, AI, ALP, ALT, AST, CRE, BUN, PCO, TSH and MDA of rats groups fed a normal diet, a normal diet with Arg, a hypercholesterolemic diet or a hypercholesterolemic diet with Arg |
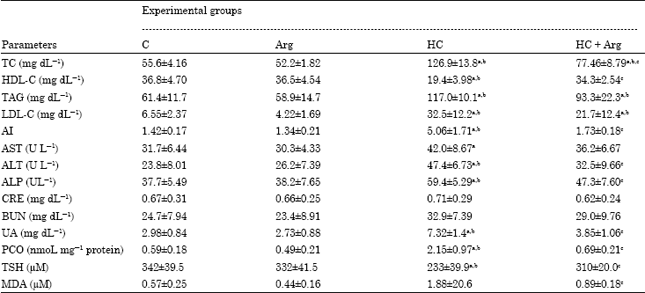 | |
| Data were tested by one-way analysis of variance and represented as Mean±SD. Six rats in each group (n = 6). Tukey’s post test was performed to determine differences between mean values. Within a row, statistically different values are marked with superscript letters when a significance was observed at p≤0.05. TC: Total cholesterol, HDL-C: High density lipoprotein cholesterol, TAG: Triacylglycerol, LDL-C: High density lipoprotein cholesterol, ALP: Alkaline phosphatase, ALT: Alanine aminotransferase, AST: Aspartate aminotransferase, CRE: Creatinine, BUN: Blood urea nitrogen, PCO: Protein carbonyl, TSH: Total thiols, MDA: Malondialdehyde | |
| Table 2: | Liver levels of Total Lipids (TL), TC and TAG, TSH and MDA expressed per (g) of liver tissues while PCO expressed per (mg) of protein of rats groups fed a normal diet with Arg, a hypercholesterolemic diet or a hypercholesterolemic diet with Arg |
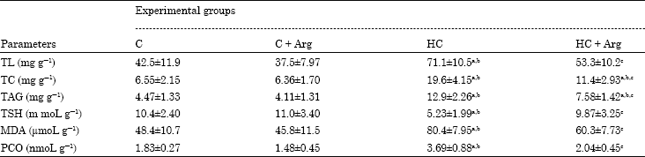 | |
| Data were tested by one-way analysis of variance and represented as Mean±SD. Six rats in each group (n = 6). Tukey’s post test was performed to determine differences between Mean values. Within a row, statistically different values are marked with a, significant from control at p≤0.05, b, significant from control plus Arg at p<0.001 and c, significant from HC at p<0.05 | |
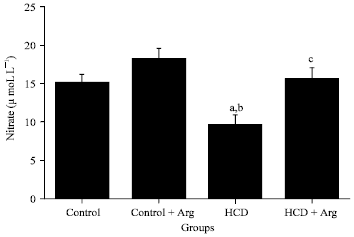 | |
| Fig. 1: | Plasma nitrate level of rats groups fed a normal diet, a normal diet supplemented with Arg, a hypercholesterolemic diet or a hypercholesterolemic diet supplemented with Arg. Data were tested by one-way analysis of variance and represented as Mean±SD. Six rats in each group (n = 6). Tukey’s post test was performed to determine differences between mean values. Within a row, statistically different values are marked with a, significant decrease from control at p<0.05, b, significant decrease from control plus Arg at p<0.001 and c, significant increase from HC at p<0.05 |
Conversely, the supplementation of Arg with hypercholesterolemic diet improves liver functions as compared with rats fed this diet alone. Furthermore, administration of Arg at dose 3% in drinking water or HCD not induced harmful effect on kidneys of rats as indicated by normal level of creatinine and urea in the investigated groups, Table 1 shows the statistical data of these investigations.
PCO and TSH were measured as a biomarker for protein oxidative damage. In the present study the additions of Arg to the normal or hypercholesterolemic diets significantly attenuate the level of PCO in plasma shown in Table 1 and in liver tissues shown in Table 2 at p<0.001. Also, Arg administration preserves TSH at value near that of normal, while the rats received HCD alone exert marked decrease of TSH level (p<0.001) for plasma and (p<0.01) for liver in respect to rat received normal diet. Regarding to MDA, the current results showed that HCD giving elicit a significant increase of this parameter in both hepatic tissues as well as plasma compared with normal diet (p<0.001). Arg treatment with HCD normalizes MDA level as compared with intake of HCD alone (p<0.001).
Intake of HCD induced significant decrease of plasma nitrate level in comparison with rats received normal dietary chow (p<0.01). On the other hand, Arg supplementation with standard diet or HCD significantly increases the nitrate level in comparison with HCD (p<0.01). The statistical data of these results is represented in Fig. 1.
ARE and PON1 activities were depicted in Table 3, the current investigations revealed that there is a significant decreased of both ARE and PON1 activities as a result of HCD feeding in comparison with normal dietary chow feeding in both plasma (p<0.001) and liver tissues (p<0.01). On the other side the supplementation of rats with 3% Arg in drinking water with standard diet or HCD markedly preserve both enzymatic activities (PON1 and ARE) in respect to the rats received HCD without any treatment (p<0.001).
| Table 3: | Paraoxonase-1(PON1) and Arylesterase (ARE) activities in plasma and liver of rats groups fed a normal diet, a normal diet supplemented with Arg, a hypercholesterolemic diet or a hypercholesterolemic diet supplemented with Arg |
 | |
| Data were tested by one-way analysis of variance and represented as Mean±SD. Six rats in each group (N = 6). Tukey’s post test was performed to determine differences between mean values. Within a row, statistically different values are marked with superscript when, significant value was observed at p≤0.05 | |
DISCUSSION
Exposure of biomolecules particularly proteins to reactive species was resulted in modification of amino acids residues, altering their structure and functions (Beal, 2002).
The feeding of rats with cholesterol rich diet resulted in an increased lipids profile and liver function tests. Arg supplementation to rats with such diet associated with amelioration of these parameters. These finding are in agreement with study of Sadeghi-Hashjin et al. (2009) reported that Arg administration improved the lipids abnormalities caused by high cholesterol feeding. Moreover, the observations reported by earlier scientists support the finding of the present study (Rossitch et al., 1991; Hurson et al., 1995). On the other hand Taboada et al. (2005) demonstrated that Arg treatment did not affect plasma lipids level in hypercholesterolemic animals model.
The uric acid is determined in this work as indicator for increase of XO activity while the creatinine and urea were determined to exclude the renal effect on plasma uric acid levels. The supplementation of rats with Arg plus hypercholesterolemic diet was associated with normalization of uric acid level. These finding are with the same observation of White et al. (2004) demonstrated that, Arg normalize the elevated XO activity induced by high blood cholesterol concentration. This effect of Arg may be due its ability to decrease hepatic injury along with decrease of elevated XO activity. This finding is supported by another study demonstrated that, Arg treatment decrease of both ALT and XO activities during liver injury (Vega et al., 2000). Moreover, it has been demonstrated that Arg administration reduces liver damage associated with increased oxidative stress (Angele et al., 2000).
Cellular thiol mostly GSH and other thiols play important role in protection of lipids and proteins against oxidative damage. Therefore, the decrease of thiols levels in biological systems, making them more susceptible to oxidative injury (Nagy et al., 2007). In the present study, supplementation of rats with Arg beside the normal or HCD preserves the thiols levels; also, Arg minimizes the protein oxidation as well as lipids peroxidation. The decreases of thiols by HCD feeding make the proteins more liable to damage by excessive free radicals produced by HC. As the results of increase oxidative stress, the protein carbonyl is increased. These findings are in agreement with the previous study demonstrated that there is an increase of protein oxidation in response to cholesterol induced oxidative stress (Ozdemirler et al., 2001). This may be attributed to increased proteins damaging at lysine, arginine, proline and threonine in the polypeptide chains (Amici et al., 1989). Administration of Arg minimizes proteins oxidation as indicated by attenuation of PCO formation in both plasma and liver tissues. These observations are similar to the study of Tripathi et al. (2009) reported that Arg administration improve lipids abnormality as well as minimizes the protein oxidation.
The decreased PON1 activity is due to either decreased HDL-C level or increased oxidative stress. This is in agreement with previous demonstration reported that the decreased of both ARE and PON1 activities due to impaired the antioxidant capacity (Dirican et al., 2004). Therefore, PON1 activity is decreased due to consumption of PON1 in the prevention of oxidative stress (Aviram et al., 1999). Free sulfhydryl at cysteine residue is necessary for the paraoxonase activity; the decrease of TSH in this investigation may be one of the causes for decrease of PON1 activity. Whereas another examination demonstrated that there is a positive correlation between TSH and PON1 activity (Shetty et al., 2007). This is supported by another study reported that oxidative stress inhibits PON activity while thiol containing antioxidants supplementation restores its activity (Costa et al., 2005). HC is associated inflammatory reactions; it well known that PON1 activity may decrease during the inflammatory condition (Baskol et al., 2006). Thus, it can also be suggested that increased utilization of PON1 for the prevention of inflammation induced by elevated blood cholesterol level. The preservation of PON1 activity by Arg supplementation may be either due to decrease of free radicals production, preservation of cellular thiols as well as maintenance of HDL-C level, whereas, many study demonstrated that Arg exert antioxidant effect (Lass et al., 2002).
Increased blood cholesterol level leads to decrease of Arg availability; consequently, the biological activity of NO is decreased. Furthermore, the NO is converted into peroxynitrite which mediated the cells damage associates with atherosclerosis (Subratty et al., 2007). In the present study nitrate level as marker for NO production is decreased by HCD feeding, these finding are corresponding to the observation of Boger et al. (2004). The same study reported that the NO synthesis is decreased in HC either due to the availability of Arg is impaired or increase of asymmetric dimethylarginine concentrations as NO synthase inhibitor.
CONCLUSION
Free radicals production is increased in HC, the thiol groups are attack by these radicals. PON1 contains free thiol groups at cysteine residues; therefore, such enzyme is inactivated by the free radicals. Arg intake improves the alteration of liver function in response to elevated cholesterol in the rat as well as it decreases the AI. Also, Arg treatment preserves NO level as well as total thiols contents, thereby reducing oxidative stress associated with HC. A reduction in XO activity by Arg treatment may be related antioxidant effect of this amino acid. These effects leading decrease the proteins oxidation as well as preservation of both ARE and PON1 of paraoxonase enzyme.
ACKNOWLEDGMENT
The author would like to thank Kayyali Research Chair for Pharmaceutical Industries, Department of Pharmaceutics, College of Pharmacy, King Saud University, Riyadh, Saudi Arabia for the assistance in completion of this study.
REFERENCES
- Amici, A., R.L. Levine, L. Tsai and E.R. Stadtman, 1989. Conversion of amino acid residues in proteins and amino acid homopolymers to carbonyl derivatives by metal-catalyzed oxidation reactions. J. Biol. Chem., 264: 3341-3346.
Direct Link - Angele, M.K., F. Fitzal, N. Smail, M.W. Knofrel and M.G. Schwacha et al., 2000. L-Arginine: Attenuates trauma-hemorrhage induced liver injury. Crit. Care Med., 28: 3242-3248.
PubMed - Aviram, M., M. Rosenblat, S. Billecke, J. Erogul and R. Sorenson et al., 1999. Human serum paraoxonase (PON 1) is inactivated by oxidized low density lipoprotein and preserved by antioxidants. Free Rad. Biol. Med., 26: 892-904.
CrossRef - Ayub, A., M.I. Mackness, S. Arrol, B. Mackness, J. Patel and P.N. Durrington, 1999. Serum paraoxonase after myocardial infarction. Arterioscler. Thromb. Vasc. Biol., 19: 330-335.
Direct Link - Baskol, G., M. Baskol, A. Yurci, O. Ozbakir and M. Yucesoy, 2006. Serum paraoxonase-1 activity and malondialdehyde levels in patient`s with ulcerative colitis. Cell Biochem. Funct., 24: 283-286.
PubMed - Beal, M.F., 2002. Oxidatively modified proteins in aging and disease. Free Radic. Biol. Med., 32: 797-803.
CrossRefDirect Link - Boger, R.H., D. Tsikas, S.M. Bode-boger, L. Phivthong-ngam, E. Schwedhelm and J.C. Frolich, 2004. Hypercholesterolemia impairs basal nitric oxide synthaseturnover rate: A study investigating the conversion of L-[guanidino-15N2]-arginine to 15N-labeled nitrate by gas chromatography-mass spectrometry. Nitric Oxide, 11: 1-8.
CrossRef - Cimen, M.Y.B., 2008. Free radical metabolism in human erythrocytes. Clinica Chimica Acta, 390: 1-11.
CrossRefDirect Link - Dalle-Donne, I., A. Scaloni, D. Giustarini, E. Cavarra and G. Tell et al., 2005. Proteins as biomarkers of oxidative/nitrosative stress in diseases: The contribution of redox proteomics. Mass Spectrometry Rev., 24: 55-99.
CrossRefDirect Link - Deepa, P.R. and P. Varalakshmi, 2006. Favorable modulation of the inflammatory changes in hypercholesterolemic atherogenesis by low-molecular-weight heparin derivative. Int. J. Cardiol., 106: 338-347.
PubMedDirect Link - Dirican, M., R. Akca, E. Sarandol and K. Dilek, 2004. Serum paraoxonase activity in uremic predialysis and hemodialysis patients. J. Nephrol., 17: 813-818.
PubMedDirect Link - Draganov, D.I. and B.N.L. Du, 2004. Pharmacogenetics of paraoxonase: A brief review. Naunyn-Schmiedeberg`s Arch. Pharmacol., 369: 78-88.
CrossRefDirect Link - El-Missiry, M.A., A.I. Othman and M.A. Amer, 2004. L-Arginine ameliorates oxidative stress in alloxan-induced experimental diabetes mellitus. J. Applied Toxicol., 24: 93-97.
CrossRefDirect Link - Feillet-Coudray, C., T. Sutra, G. Fouret, J. Ramos and C. Wrutniak-Cabello et al., 2009. Oxidative stress in rats fed a high-fat high-sucrose diet and preventive effect of polyphenols: Involvement of mitochondrial and NAD (P) H oxidase systems. Free Radic. Biol. Med., 46: 624-632.
PubMed - Folch, J., M. Lees and G.H.S. Stanley, 1957. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem., 226: 497-509.
CrossRefPubMedDirect Link - Ozdemirler, G., S. Kucuk, Y. Orhan, G. Aykac-Toker and M. Uysal, 2001. Lipid and protein oxidation in erythrocyte membranes of hypercholesterolemic subjects. Clin. Biochem., 34: 335-339.
PubMedDirect Link - Honing, M.L.H., P.J. Morrison, J.D. Banga, E.S.G. Stroes and T.J. Rabelink, 1998. Nitric oxide availability in diabetes mellitus. Diabetes Metab. Rev., 14: 241-249.
Direct Link - Hu, M.L., 1994. Measurement of protein thiol groups and glutathione in plasma. Methods Enzymol., 233: 380-385.
CrossRefPubMedDirect Link - Kawano, H., T. Motoyama, N. Hirai, K. Kugiyama, H. Yasue and H. Ogawa, 2002. Endothelial dysfunction in hypercholesterolemia is improved by L-Arginine administration: Possible role of oxidative stress. Atherosclerosis, 161: 375-380.
Direct Link - Kumar, S.A., V. Sudhahar and P. Varalakshmi, 2006. Oxidative renal injury and lipoprotein oxidation in hypercholesterolemic atherogenesis: Role of eicosapentaenoate-lipoate (EPA-LA) derivative. Prostaglandins Leukot. Essent. Fatty Acids., 75: 25-31.
CrossRef - Lass, A., A. Suessenbacher, G. Wolkart, B. Mayer and F. Brunner, 2002. Functional and analytical evidence for scavenging of oxygen radicals by L-Arginine. Mol. Pharmacol., 61: 1081-1088.
PubMedDirect Link - Levine, R.L., J.A. Williams, E.R. Stadtman and E. Shacter, 1994. Carbonyl assay for determination of oxidatively modified proteins. Methods Enzymol., 233: 346-357.
PubMedDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Marjani, A., A. Moradi and A.B. Ghourcaie, 2007. Alterations in plasma lipid peroxidation and erythrocyte superoxide dismutase and glutathione peroxidase enzyme activities during storage of blood. Asian J. Biochem., 2: 118-123.
CrossRefDirect Link - McGrowder, D. and P.D. Brown, 2007. Effects of nitric oxide on glucose transport: In vivo and in vitro studies. Asian J. Biochem., 2: 1-18.
CrossRefDirect Link - Moshage, H., B. Kok, J.R. Huizenga and P.L. Jansen, 1995. Nitrite and nitrate determinations in plasma: A critical evaluation. Clin. Chem., 41: 892-896.
PubMedDirect Link - Nagy, L., M. Nagata and S. Szabo, 2007. Protein and non-protein sulfhydryls and disulfides in gastric mucosa and liver after gastrotoxic chemicals and sucralfate: Possible new targets of pharmacologic agents. World J. Gastroenterol., 13: 2053-2060.
PubMedDirect Link - Ohkawa, H., N. Ohishi and K. Yagi, 1979. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem., 95: 351-358.
CrossRefPubMedDirect Link - Rossitch, Jr. E., E. Alexander 3rd, P.M. Black and J.P. Cooke, 1991. L-Arginine normalizes endothelial function in cerebral vessels from hypercholesterolemic rabbits. J. Clin. Invest., 87: 1295-1299.
PubMed - Rozenberg, O., M. Rosenblat, R. Coleman, D.M. Shih and M. Aviram, 2003. Paraoxonase (PON1) deficiency is associated with increased macrophage oxidative stress: studies in PON1-knockout mice. Free Radic. Biol. Med., 34: 774-784.
CrossRef - Sadeghi-Hashjin, G., M. Abuhosseini and S. Asri-Rezaei, 2009. Role of nitric oxide in the plasma lipid profile in the rabbits. Arch. Med. Sci., 5: 308-312.
Direct Link - Serdar, Z., K. Aslan, M. Dirican, E. Sarandol, D. Yesilbursa and A. Serdar, 2006. Lipid and protein oxidation and antioxidant status in patients with angiographically proven coronary artery disease. J. Clin. Biochem., 39: 794-803.
Direct Link - Shetty, J.K., M. Prakash, M. Verma, S. Tripathy and P. Vickram, 2008. Effect of atorvastatin on paraoxonase activity in patients with hyperlipidemia. Asian J. Biochemistry, 3: 139-142.
CrossRefDirect Link - Shetty, J.K., P. Mungli, T. Sudeshna, V. Manish, K. Nagaraj Shashidhar and S. Pampapathy, 2007. Serum paraoxonase activity and protein thiols in chronic renal failure patients. Asian J. Biochem., 2: 274-278.
CrossRefDirect Link - Subratty, A.H., L.H. Semfa and M.D. Manraj, 2007. TAME-esterase and oxidative stress contribute to dysmetabolic syndrome in dyslipidaemia. Asian J. Biochem., 2: 323-329.
CrossRefDirect Link - Taboada, M.C., B. Rodriguez, R. Millan and I. Miguez, 2005. Role of dietary L-Arginine supplementation on serum parameters and intestinal enzyme activities in rats fed an excess-fat diet. Biomed. Pharmacother., 60: 10-13.
CrossRef - Tripathi, P., M. Chandra and M.K. Misra, 2009. Oral administration of L-Arginine in patients with angina or following myocardial infarction may be protective by increasing plasma superoxide dismutase and total thiols with reduction in serum cholesterol and xanthine oxidase. Oxid. Med. Cell. Longev., 2: 231-237.
PubMedDirect Link - Tsai, H.J., H.F. Shang, C.L. Yeh and S.L. Yeh, 2002. Effects of Arginine supplementation on antioxidant enzyme activity and macrophage response in burned mice. Burns, 28: 258-263.
PubMedDirect Link - Vega, V.L., L. Mardones, M. Maldonado, S. Nicovani, V. Manriquez, J. Roa and P.H. Ward, 2000. Xanthine oxidase released from reperfused hind limbs mediate kupffer cell activation, neutrophils sequestration and hepatic oxidative stress in rats subjected to tourniquet shock. Shock, 14: 565-571.
PubMedDirect Link - White, C., D. Parks, R. Patel, J. Shelton, M. Tarpy, B. Freeman and V. Darley-Usmar, 2004. L-Arginine inhibits xanthine oxidase-dependent endothelial dysfunction in hypercholesterolemia. FEBS Lett., 561: 94-98.
Direct Link