
M.M. Soliman
Department of Biochemistry, Faculty of Veterinary Medicine, Moshtohor, P.O. Box 13736, Benha University, Egypt
Y.A. El-Fattah El-Senosi
Department of Biochemistry, Faculty of Veterinary Medicine, Moshtohor, P.O. Box 13736, Benha University, Egypt
O.M.A. El-Hamid
Department of Biochemistry, Faculty of Veterinary Medicine, Moshtohor, P.O. Box 13736, Benha University, Egypt
A.El-Desouki Abd El-Mageed
Department of Biochemistry, Faculty of Veterinary Medicine, Moshtohor, P.O. Box 13736, Benha University, Egypt
R.S. Ismaeil
Departments of Physiology, Faculty of Veterinary Medicine, Moshtohor, P.O. Box 13736, Benha University, Egypt
H.A. El-Maqsoud Ali
Department of Biochemistry, Faculty of Veterinary Medicine, Moshtohor, P.O. Box 13736, Benha University, Egypt
Asian Journal of Biochemistry
Year: 2009 | Volume: 4 | Issue: 2 | Page No.: 60-67







ABSTRACT
In this study, we examined the effect of either lipopolysaccharide (LPS, 1 μg mL–1), Nigella.sativa extract (NS, 5 μg mL–1), or co-treatment of both for 24 h on cytokines expression in mature bovine adipocytes using RT-PCR analysis. The results showed that separate treatment by LPS and NS stimulated the expression of interleukin-1 (IL-1β), IL-6, IL-8 and IL-10. Co-treatment of cells by N. sativa with LPS inhibited LPS induced IL-6 and TNF-α expression and induced additive stimulatory effect on LPS induced IL-8 and IL-10 expression. The results indicate that N. sativa extract has immuno-modulatory effect on bovine adipocytes by stimulating different cytokines expression that potentiate different inflammatory and anti-inflammatory functions in bovine adipocytes.
PDF Abstract XML References Citation
How to cite this article
M.M. Soliman, Y.A. El-Fattah El-Senosi, O.M.A. El-Hamid, A.El-Desouki Abd El-Mageed, R.S. Ismaeil and H.A. El-Maqsoud Ali, 2009. Nigella sativa Modulates Cytokines Expression in Mature Bovine Adipocytes. Asian Journal of Biochemistry, 4: 60-67.
DOI: 10.3923/ajb.2009.60.67
URL: https://scialert.net/abstract/?doi=ajb.2009.60.67
DOI: 10.3923/ajb.2009.60.67
URL: https://scialert.net/abstract/?doi=ajb.2009.60.67
INTRODUCTION
The seeds of Nigella sativa (NS) which are commonly known as black seed or black cumin are used in herbal medicine all over the world for treatment and prevention of a number of diseases and conditions that include asthma, hypertension, diabetes, inflammation, cough, bronchitis, headache, eczema, fever, dizziness and influenza (Salem, 2005). The seeds or its oil are used as a carminative, diuretic, lactagogue and vermifuge. Nigella sativa is used in food as a spice and a condiment (Ali and Blunden, 2003). NS has an antitussive effect and induces codeine like effect (Boskabady et al., 2004). It contains more than 30% fixed oil and 0.40-0.45% w/w volatile oil. The volatile oil has been shown to contain 18-24% thymoquinone and 46% comprises many monoterpenes such as p-cymene and (Pinene 3) (EL-Tahir et al., 1993). Nigella sativa was recently subjected to extensive pharmacological investigations that revealed its antioxidant activity in different organs and acted as anticancer drugs (Houghton et al., 1995; Worthen et al., 1998). However, the effects of NS extract on bovine adipocytes function and alteration are still unknown.
Cytokines are relatively low-molecular-weight proteins produced by many cell types (Feghali and Wright, 1997). They are pharmacologically active, exhibiting both beneficial and pathologic effects on the target cells. Imbalanced expression of cytokines has been implicated in the progression of many diseases (Arend and Gabay, 2004). TNF-α has been shown to enhance anti-tumor responses and to promote tumor regression (Peron et al., 1999). IL-6 is a cytokine produced by a number of normal and transformed cells (Bartold and Haynes, 1991) and promotes or inhibits the growth of tumor cells depending upon the cell type (Feghali and Wright, 1997).
White adipose tissue secrets different cytokines that are important in modulating body defensive mechanisms such as IL-1, IL-6, TNF, IL-10 and IL-8. Adipose tissue also secrets various proteins as leptin, adiponectin and haptoglobin which are involved in regulation of different body conditions (Havel, 2004).
Most of already established findings focused on the effect of NS on cytokines production from lymphocytes and peripheral blood mononuclear cells (El-Obeid et al., 2005). For our knowledge, there are no data available about the effect of NS on bovine adipocytes and that is the purpose of this study.
MATERIALS AND METHODS
Materials
Dulbecco’s Modified Eagles’s Medium (DMEM), Bovine Serum Albumin (BSA) and Hank’s balanced salt solution were bought from Sigma-Aldrich Fine Chemical (St. Louis, MO, USA). Fetal Calf Serum (FCS) was from Trace Scientific Ltd. (Melbourne, Australia), LPS and collagenase were purchased from Wako Pure Chemicals Co. (Osaka, Japan).
Isolation of Mature Adipocytes
The experimental procedure and care of animals were in accordance with the guidelines of the Animal Care and Use Committee of Benha University. Subcutaneous adipose tissue (S/C AT) was obtained from three non-pregnant and non-lactating healthy Holstein cows (4 years old). (S/C AT) was dissected into small pieces and immersed in Hank’s balanced salt solution containing 2 mg mL–1 collagenase and 0.1% BSA in sterile 50 mL plastic tube for digestion. After digestion at 37°C for 90 min with gentle shaking, the solution was filtered through sterile nylon mesh with 80 μm pores. The filtrate was centrifuged at 1000xg for 5 min at room temperature. The supernatant cells (mature adipocytes) were washed twice with DMEM containing 10% FCS, 100 U mL–1 penicillin and 100 μg mL–1 streptomycin. The cells were counted by haemocytometer and the percentage of live to dead cells was encountered by tryban blue staining. 5x106 cells were used for each milliliter media. The cells were cultured in DMEM (2.5 mM glucose) containing 10% FCS 6-well plates under different experimental conditions for 24 h.
Preparation of Nigella sativa Extract
As described before by Ibrahim et al. (2008), N. sativa seeds were purchased from the local Egyptian market. The seeds were washed, dried and crushed to a powder with an electric microniser. 20 g of the powder was dissolved in 200 mL–1 phosphate buffer saline (pH 6.4) and centrifuged at 10000 rpm for 30 min at 4°C to remove residual solid debris. The clear supernatant was collected as the soluble extract after removal of the oily layer and insoluble pellet. Protein concentration was determined by Lowry et al. (1951) method.
Experimental Procedures and RT-PCR Analysis
Cells were incubated with PBS as control or LPS (μg mL–1) and/or with NS extract (5 μg mL–1). The dose of both LPS and NS was determined after several preliminary traits and it was the optimal sublethal dose. Cells were incubated at 37°C in 5% CO2 for 24 h (optimal time to induce different gene expression in cattle adipocytes). Total cellular RNA was isolated from cultured adipocytes by the guanidine-isothiocyanate method using TRIzol reagent (Gibco BRL, Rochville, MD, USA). RNA (1 μg) was treated at 72°C for 5 min and reverse transcribed using 100 units of Moloney murine leukemia virus reverse transcriptase (Gibco), 50 pmol of poly (dT) primer and 20 nmol of dNTPs in a total volume of 10 μL at 37°C for 1 h. After heating at 94°C for 5 min, PCR amplification was performed with 2.5 units Taq polymerase (Perkin-Elmer, Foster City, CA, USA), 3 mM MgCl2 and 50 pmol of forward and reverse primers specific for respective genes in a total volume of 50 μL. The PCR conditions for different tested genes are shown in Table 1. The experiments were repeated 3 times on mature adipocytes of 3 cows. After electrophoresis in 1.5% agarose gel, the PCR products were stained with ethidium bromide and visualized under UV lamp.
| Table 1: | PCR primers |
 | |
| In the parenthesis, amplification and size of PCR product are shown. Annealing temperature, time and number of PCR cycle of respective genes are also shown, while temperature and time of denaturation and elongation steps of each PCR cycle are 94°C, 30 sec and 72°C, 60 sec, respectively | |
Statistical Analysis
All data were expressed as Means±SEM and analyzed by Dunnett's t-test for multiple comparisons with a single control group using Stat View program. Significance was reported as p<0.05.
RESULTS
Effect of NS Extract on IL-6, TNF-α andIL-1β mRNA Expression
In Fig. 1 and 2, treatment of bovine adipocytes with LPS up-regulated significantly (p<0.05) mRNA expression of TNF-α and IL-6. LPS is known to be the potent stimulator of different cytokines during acute phase inflammation and/or diseases. NS extract up-regulated IL-6 mRNA expression but did not affect TNF-α expression. Co-treatment of NS extract and LPS more potently down-regulated mRNA expression of TNF-α and IL-6 than that induced by LPS treatment alone.
Regarding IL-1 β expression, as shown in Fig. 3, LPS induced 2 fold increase (p<0.05) in IL-1β expression while treatment of cells with NS extract induced IL-1β expression (about 1.8 fold increase) but co-treatment of LPS with NS extract did not show any changes in IL-1β expression than that induced by either LPS or NS extract although very small additive stimulatory effect is seen but not significant. These findings clearly and collectively show the role of NS extract in inhibiting the proinflammatory mediators, which are known as acute phase cytokines.
Effect of NS Extract on IL-8 and IL-10 mRNA ExpressionIL-8 is a cytokine produced by adipocytes, monocytes, neutrophils and NK cells and acts as a chemoattractant factor for neutrophils, basophils and T-cells. Treatment of mature adipocytes by LPS did not affect IL-8 mRNA expression as shown in Fig. 4 but NS extract stimulated IL-8 mRNA expression. Combination of both LPS and NS extract partially induced additive effect on IL-8 expression. IL-10 is anti-inflammatory cytokine that inhibits pro-inflammatory cytokines. On the same line, treatment of bovine adipocytes by LPS (Fig. 5), induced 6 fold increase in IL-10 mRNA expression (p<0.05). NS extract treatment slightly induced IL-10 expression (1 fold increase). Combination of both LPS and NS extract showed an additive up-regulation in mRNA expression of IL-10. Those findings clarify the role of N. Sativa as chemoattractant and regenerative factor to potentiate different body defensive mechanisms.
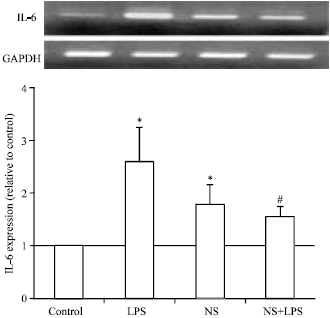 | |
| Fig. 1: | Effect of Nigella sativa on LPS induced IL-6 expression in bovine adipocytes. Mature bovine adipocytes were cultured in 6-well plated at 5x106 per well with or without 5 μg mL–1 in presence or absence of LPS (1 μg mL–1). Two microgram RNA was extracted and reverse transcribed using RT-PCR analysis for IL-6 expression. The bands of both genes were visualized using 1.5% gel stained with ethedium bromide. Intensity of expression was normalized with that of GAPDH (as internal standard) using NIH image for densitometric analysis. Values are expressed as Mean±SEM obtained from 3 experiments. *p<0.05 vs. control and #p<0.05 vs. LPS |
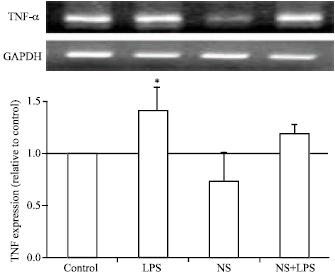 | |
| Fig. 2: | Effect of N. sativa on LPS induced TNF-α expression in bovine adipocytes. Mature bovine adipocytes were cultured as described in Fig. 1. Two microgram RNA was reverse transcribed and subjected to RT-PCR analysis using specific primers for TNF-α and GAPDH as internal standard. Values are expressed as Mean±SEM obtained from 3 experiments. *p<0.05 vs. control |
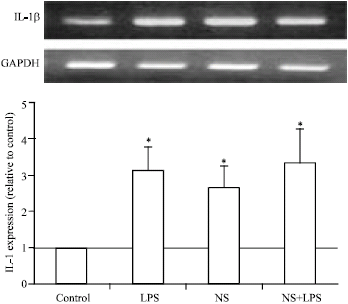 | |
| Fig. 3: | Effect of N. sativa on LPS induced IL-1-β expression in bovine adipocytes. Mature bovine adipocytes were cultured as described in Fig. 1. Two microgram RNA was reverse transcribed and subjected to RT-PCR analysis using specific primers for IL1-β and GAPDH as internal standard. Values are expressed as Mean±SEM obtained from 3 experiments. *p<0.05 vs. control |
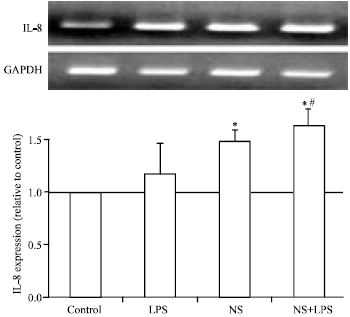 | |
| Fig. 4: | Effect of N. sativa on LPS induced IL-8 expression in bovine adipocytes. Mature bovine adipocytes were cultured as described in Fig. 1. Two microgram RNA was reverse transcribed and subjected to RT-PCR analysis using specific primers for IL-8 and GAPDH as internal standard. Values are expressed as Mean±SEM obtained from 3 experiments. *p<0.05 vs. control and #p<0.05 vs. LPS |
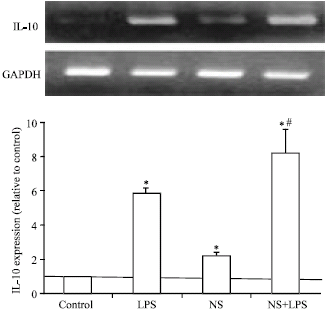 | |
| Fig. 5: | Effect of N. sativa on LPS induced IL-10 expression in bovine adipocytes. Mature bovine adipocytes were cultured as described in Fig. 1. Two microgram RNA was reverse transcribed and subjected to RT-PCR analysis using specific primers for IL-10 and GAPDH as internal standard. Values are expressed as Mean±SEM obtained from 3 experiments. *p<0.05 vs. control and #p<0.05 vs. LPS |
DISCUSSION
The present findings showed the immuno-modulatory effect of N. sativa extract on LPS induced different inflammatory cytokines. N. Sativa extract inhibited LPS-induced TNF-α and IL-6 expression and induced additive effect on LPS stimulated IL-8 and IL-10 expression. The exact mechanism by which NS extract induce its action is unclear. Ibrahim et al. (2008) showed that NS inhibited hepatotoxicity induced by carbon tetra chloride by down regulation of CYP2E and CYP3A protein expressions due to reduction of nitrous oxide through downregulation of inducible Nitrous Oxide Synthase (iNOS) expression. Salem (2005) reported that N. sativa inhibited the inflammatory cytokines IL-1 and TNF-α and enhanced IL-8 expression in human Peripheral Blood Mono-nuclear Cells (PBMC) through an alteration of trafficking of inflammatory cells via., modulating the expression of chemokines and or other adhesion molecules. Therefore, N. sativa helps to balance the inflammatory response during endotoxemia as reported by present findings and that of Howard et al. (1993).
IL-8 is a chemokine produced by a wide variety of cell types in response to LPS (Yoshimura et al., 1987) by investigating the effect of N. sativa seed proteins on cytokine production in humans PBMC, it enhanced the production of IL-3 and IL-1 by lymphocytes when cultured with or without allogeneic cells (Haq et al., 1995). Present findings showed the additive stimulatory effect of NS extract on LPS stimulated IL-1 expression and its role as an initiator of inflammation in cattle. Of note, stimulatory effect of whole N. sativa and their fractionated proteins was also noticed on the production of TNF-α by either non-activated or mitogen-activated PBMC (Haq et al., 1999) and that are parallel to present results regardless the difference in cell types and species.
Interleukin-10 (IL-10) is a potent anti-inflammatory cytokine. It inhibits IL-1 and IL-6 production from macrophages (Fiorentino et al., 1991). This cytokine plays a critical role in shaping the immune response by blocking class II major histocompatibility complex expression and decreasing pro-inflammatory cytokine expression (Donnelly et al., 1999; Moore et al., 2001) and that is similar to present findings.
In conclusion, aqueous extract of NS showed immune-modulatory effect on bovine adipocytes through down-regulation of IL-6 and TNF and up-regulation of IL-10 and IL-8 expression induced by LPS treatment. Moreover, bovine adipocytes play an important role in balancing different inflammatory responses that happened during inflammation and or disease like blood mono-nuclear cells.
REFERENCES
- Ali, B.H. and G. Blunden, 2003. Pharmacological and toxicological properties of Nigella sativa. Phytother. Res., 17: 299-305.
CrossRefPubMedDirect Link - Arend, W. and C. Gabay, 2004. Cytokines in the rheumatic diseases. Rev. Rheum. Dis. Clin. North Am., 30: 41-67.
PubMed - Bartold, P.M. and D.R. Haynes, 1991. Interleukin-6 production by human gingival fibroblasts. J. Periodontal Res., 26: 339-345.
CrossRefPubMedDirect Link - Boskabady, M.H., S. Kiani, P. Jandaghi, T. Ziaei and A. Zarei, 2004. Antitussive effect of Nigella sativa in Guinea pigs. Pak. J. Med. Sci., 3: 224-228.
Direct Link - El-Obeid, A., S. Al-Harbi, S.N. Al-Jomaha and A. Hassib, 2005. Herbal melanin modulates tumor necrosis factor alpha (TNF-a), interleukin 6 (IL-6) and vascular endothelial growth factor (VEGF) production. Phytomedicine, 13: 324-333.
PubMed - El Tahir, K.E.H., M.M.S. Ashour and M.M. Al-Harbi, 1993. The respiratory effects of the volatile oil of the black seed (Nigella sativa) in guinea-pigs: Elucidation of the mechanism(s) of action. Gen. Pharmacol.: Vasc. Syst., 24: 1115-1122.
CrossRefPubMedDirect Link - Donnelly, R.P., H. Dickensheets and D.S. Finbloom, 1999. The interleukin-10 signal transduction pathway and regulation of gene expression in mononuclear phagocytes. J. Interferon Cytokine Res., 19: 563-573.
PubMed - Feghali, C.A. and T.M. Wright, 1997. Cytokines in acute and chronic inflammation. Front. Biosci., 2: 12-26.
CrossRefPubMedDirect Link - Fiorentino, D.F., A. Zlotnik, T.R. Mosmann, M. Howard and A. O'Garra, 1991. IL-10 inhibits cytokine production by activated macrophages. J. Immunol., 147: 3815-3822.
PubMed - Havel, J.P., 2004. Updates on adipocytes hormones: Regulation of energy balance and carbohydrate∕ lipid metabolism. Diabetes, 53: S143-S151.
PubMed - Haq, A., P.I. Lobo, M. Al-Tufail, N.R. Rama and S. Al-Sedairy, 1999. Immunomodulatory effect of Nigella sativa proteins fractionated by ion exchange chromatography. Int. J. Immunopharmacol., 21: 283-295.
CrossRefPubMedDirect Link - Houghton, P.J., R. Zarka, B. de las Heras and J.R.S. Hoult, 1995. Fixed oil of Nigella sativa and derived thymoquinone inhibit eicosanoid generation in leukocytes and membrane lipid peroxidation. Planta Med., 61: 33-36.
CrossRefPubMedDirect Link - Howard, M., T. Muchamuel, S. Andrade and S. Menon, 1993. Interleukin 10 protects mice from lethal endotoxemia. J. Exp. Med., 177: 1205-1208.
CrossRefPubMedDirect Link - Ibrahim, S., M. Ishizuka, M. Soliman, K. El-Bohi and K. Sobhy et al., 2008. Protection by Nigella sativa against carbon tetrachloride induced downregulation of hepatic cytochrome P450 isozymes in rats. Japanese J. Vet. Res., 56: 119-128.
PubMed - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Moore, K.W., R. de Waal Malefyt, R.L. Coffman and A. O'Garra, 2001. Interleukin-10 and the interleukin-10 receptor. Annu. Rev. Immunol., 19: 683-765.
CrossRefPubMedDirect Link - Salem, M.L., 2005. Immunomodulatory and therapeutic properties of the Nigella sativa L. seed. Int. Immunopharmacol., 5: 1749-1770.
CrossRefPubMedDirect Link - Yoshimura, T., K. Matsushima, S. Tanaka, A. Robinson, E. Appella, J. Oppenheim and J. Leonard, 1987. Purification of a human monocyte-derived neutrophil chemotactic factor that has peptide sequence similarity to other host defense cytokines. Proc. Natl. Acad. Sci., 84: 9233-9237.
PubMed - Worthen, D., O. Ghoshen and P. Crooks, 1998. The in vitro anti-tumor activity of some crude and purified components of black seed, Nigella sativa. Anticancer Res., 18: 1527-1533.
PubMed