
H.O.T. Iyawe
Department of Biochemistry, Ambrose Alli University, Ekpoma, Nigeria
A.O. Onigbinde
Department of Biochemistry, Ambrose Alli University, Ekpoma, Nigeria
Asian Journal of Biochemistry
Year: 2009 | Volume: 4 | Issue: 1 | Page No.: 30-35







ABSTRACT
The effect of malaria parasites and chloroquine in mice was examined. The importance of this study derives from the prevalence of malaria in the tropical and subtropical regions, as well as the declining therapeutic efficacy of chloroquine as a first line treatment against malaria infection in these endemic areas. This study aimed to determine the pattern of possible alterations in some haematological and antioxidant molecules in mice treated with either Plasmodium or chloroquine. Three groups of ten mice each categorized as control, non parasitized chloroquine treated (NPcqT) and Parasitized non treated (PnT) were used in this study. Observations from the work show that parasites in mice significantly (p<0.05) increased plasma total protein, globulin, erythrocyte fragility, total bilirubin, oxidative stress, glucose-6-phosphate dehydrogenase (G6PD), liver superoxide dismutase (SOD) and catalase (CAT) enzyme activities. Also the study showed that there is a significant (p>0.05) decrease in plasma SOD, CAT, reduced glutathione (GSH), liver G6PD and GSH. Parasitemia also reduced significantly (p<0.05) mice packed cell volume (PCV). Chloroquine treatment of Non Parasitized (NP) mice increased significantly (p<0.05) erythrocyte fragility, plasma total bilirubin, oxidative stress, but reduced (p<0.05) mice PCV, plasma SOD, CAT, G6PD, GSH but increased (p<0.05) liver SOD, CAT and reduced GSH significantly (p<0.05). The results obtained from the statistical analysis of data suggest that both malaria parasites increase oxidative stress in mice and chloroquine increases SOD and CAT activity in hepatic tissue of mice.
PDF Abstract XML References Citation
How to cite this article
H.O.T. Iyawe and A.O. Onigbinde, 2009. Impact of Plasmodium berghei and Chloroquine on Haematological and Antioxidants Indices in Mice. Asian Journal of Biochemistry, 4: 30-35.
DOI: 10.3923/ajb.2009.30.35
URL: https://scialert.net/abstract/?doi=ajb.2009.30.35
DOI: 10.3923/ajb.2009.30.35
URL: https://scialert.net/abstract/?doi=ajb.2009.30.35
INTRODUCTION
The World Health Organization (2000b) estimates that there are 270 million clinical cases of malaria disease annually resulting in at least 1.5-2.7 million deaths a year. The persistence of malaria as a public health problem is partly as a result of the emergence of Plasmodium species resistant to antimalarial drugs and insecticides, leading to increased morbidity and mortality (Lege-Oguntoye et al., 1989). The most vulnerable group for malaria infection are children and pregnant women (WHO, 2000a).
The parasite poses severe challenge to infected individuals mainly through the development of anaemia in association with other haematological changes. One of the reasons for the development of malarial anaemia seems to be oxidative stress resulting from the destruction of infected erythrocytes (Das and Nanda, 1999; Kremsner et al., 2000). Also, infectious diseases including malarial are known to activate the body phagocytes causing release of reactive oxygen species (ROS) as an antimicrobial action. Phagocytes generated ROS are non-specific effector molecules in their mode of defense mechanism thus, damaging parasitized and non-parasitized red cells (Kulkarni et al., 2003).
In most endemic areas, chloroquine use to be the main first line therapy for malaria (Olanrewaju and Johnson, 2001) until recently when WHO succeeded in promoting the combination treatment for malaria infection (Nosten and Brasseur, 2002).
A recent study (Umar et al., 2008) conducted in Nigeria has rated chloroquine efficacy in children under the age of five years with uncomplicated malaria parasites 72% in terms of adequate clinical and parasitological responses. Failure rate was 23%. It has been reported that during the development stages of Plasmodia, the parasite deplete the erythrocytes of its defence molecules namely, superoxide dismutase (SOD); catalase; NADH+; NADPH+; glutathione reductase and glutathione peroxidase. Also that oxidative stress experienced by Red Blood Cell (RBC) during Plasmodium vivax infection is accompanied with changes in the antioxidant defence system of the host which are restore to near normal level after treatment with chloroquine (Mishra et al., 1994; Sarin et al., 1993; Agomo et al., 1998).
Chloroquine therapy has been relegated in the treatment of malaria infection (Nosten and Brasseur, 2002), but implicated in the management of oxidative stress occasioned by malaria parasite (Mishra et al., 1994; Sarin et al., 1993); it therefore becomes necessary to determine the specific role chloroquine could play within the milieu of ROS produced during normal respiratory functions.
MATERIALS AND METHODS
Animals
Thirty albino male mice of eight weeks, bred at the animal house unit of College of Medicine, Ambrose Alli University, Ekpoma were used as subjects for this study. The animals were observed for seven days for any sign of ill health. At the end of the observation period, the animals were divided into eight groups of ten mice each.
Procedures
Three groups of animals respectively categorized as control and non parasitized chloroquine treated (NPcqT) and parasitized and non treated (PnT) was used. The animals were allowed free access to feed on Grower’s mash (from Bendel feeds and flourmills Ltd. Ewu Edo State Nigeria) and water. At the end of the experiment, the mice were anaesthetized with chloroform and blood collected by cardiac puncture into sample tubes from where serum used for assay was harvested after clotting and centrifugation.
Parasites
ANKA strain of Plasmodium berghei was maintained in the laboratory by syringe passage of parasitized blood into mice. A standard dose of 104 red blood cells mL-1 was inoculated intraperitoneally and parasitemia was assessed from Giemsa stained thin blood films (WHO, 2000).
Drug Preparation and Administration
Five hundred milligram (500 mg) Chloroquine phosphate containing 300 mg chloroquine base (Swiss Pharma Nigeria Ltd.) certified by National Agency for Food Drug Administration and Control (NAFDAC) was used. Each tablet was dissolved in 100 mL of distilled water and the resulting mixture centrifuged to obtain clear chloroquine solution. This preparation brought chloroquine concentration to 3 mg mL-1, which was then administered intraperitoneally (25 mg kg-1 b.wt.) for three days.
Tissue Extracts Preparation
Kidney, liver and heart tissues of subjects were obtained. The tissues were respectively defatted and half gram of each tissue was cut into tiny pieces followed by homogenization with ground glass using mortal and pestle. The mixtures were then centrifuge at 2,000 rpm for five minutes. The clear supernatants obtained from the process were filtered through Whatman No. 1filter paper and used for the assay.
Assays
Lipid peroxidation was determined as described by Gutteridge and Wilkins (1982), Superoxide dismutase assay by the method of Misra and Fridovich (1972), Catalase activity assay (Sinha, 1972), Assay of glutathione (Moron et al., 1979), the method of Keller (1970) was used to determine glucose-6-phosphate dehydrogenase activity (G6PD), gamma glutamyltransferase activity (GGT) was quantified using the method of Szaz (1976). Alanine aminotransferase (ALT) and aspartate aminotransferase (AST) assay were determined as described by Reitman and Frankel (1957), serum bilirubin (total, conjugated and unconjugated) were determined by the diazo reaction method (Noslin, 1960). Total serum albumin and protein were estimated according to the Dumas et al. (1971) and Gornall et al. (1949), respectively. Globulin concentration was obtained by difference between total protein and albumin.
Erythrocyte fragility was determined by modifying the standard cyanomethaemoglobin method of Fairbanks and Klee (1987). Briefly, A 1000 μL whole blood was pipette into a test tube and 6 mL of Drabkins solution was added and the mixture was allow to stand for 5 min after which, the absorbance reading was taken at 540 m. A standard solution 1 mg mL-1 of bovine haemoglobin was prepared. Serial dilutions were made from the stock solution and absorbance of the corresponding concentrations of haemoglobin determined at 540 m. Erythrocyte fragility was then calculated as percentage with the given expression:
RESULTS
Parasites in mice led to significant (p<0.05) increase in total serum protein and globulin compared to control and non-parasitized and chloroquine treated (NPcq) mice (Table 1). Erythrocyte fragility was observed to be higher (p<0.05) in PnT mice group compared to the other groups. In PnT and NpcqT groups, packed cell volumes (PCV) were significantly (p<0.05) reduced compared to control and the total and direct bilirubin in NPcqT and PnT groups increased (p<0.05) compared to control (Table 2). Treatment of non-parasitized mice with chloroquine and PnT groups increased (p<0.05) oxidative stress. Superoxide dismutase (SOD) and catalase (CAT) activities in both groups decreased (p<0.05) in comparison to control group and glucose-6-phoshate dehydrogenase (G6PD) activity in PnT mice increased (p<0.05) compared to control and NPcqT groups (Table 3). Chloroquine treatment in non-parasitized (NpnT) mice and parasitized non treated (PnT) groups does not appear to have any significant effect on the antioxidant parameters of mice kidney and heart organs. Parasites in non- treated groups reduced (p<0.05) liver SOD, CAT and G6PD activities (Table 4).
| Table 1: | Effect of Plasmodium berghei and chloroquine on plasma proteins and liver function enzymes of mice |
 | |
Mean±SD triplicate determinations (n = 10). Values in same row with different signs are significantly different (p<0.05) | |
| Table 2: | Effect of Plasmodium berghei and chloroquine on haematological parameters of mice |
 | |
Mean±SD triplicate determinations (n = 10). Values in same row with different signs are significantly different (p<0.05) | |
| Table 3: | Effects of Plasmodium berghei and chloroquine on MDA, glutathione and antioxidant enzymes of mice |
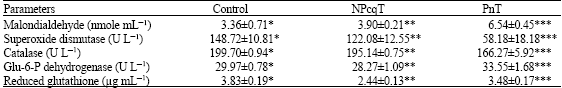 | |
Mean±SD triplicate determinations (n = 10). Values in same row with different signs are significantly different (p<0.05) | |
| Table 4: | Effects of Plasmodium berghei and chloroquine on MDA, glutathione and some antioxidant enzymes activities in mice tissue |
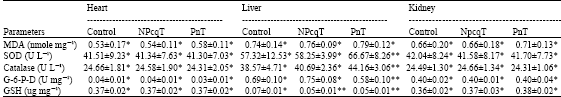 | |
| Mean±SD triplicate determinations (n = 10). Values in same row for a particular organ with different signs are significantly different (p<0.05) | |
DISCUSSION
The presence of parasitemia in mice was found to increase serum globulin and led to increase in mice total serum protein. The increase in globulin can be attributed to cellular mobilization of T-cells and its complements to mount immunity against parasite activity through synthesis and secretion of antibody molecules that forms part of immunoglobulin proteins.
Erythrocyte fragility is a measure of erythrocyte reduction through destruction of haemoglobin. In this study, erythrocyte fragility was monitored as decrease in haemoglobin content in blood. A low value of haemoglobin is indicative of high erythrocyte fragility. Fragility of erythrocyte is observed to be higher in PnT mice group. This is certainly as a result of increased haemolysis in PnT as the parasites derive their nutrients for metabolic activities from erythrocyte haemoglobin, in the process haem and globin molecules are produced. Haemoglobin destruction in parasitized cells is confirmed in this study, by the decrease in infected mice Packed Cell Volumes (PCV) levels compared to control.
The oxidative stress observed in PnT mice may not be excluded from the destruction of haemoglobin and the digestion of haem in the parasite acidic food vacuole to release excess free iron II atom into erythrocyte cytoplasm. In the presence of free iron and oxygen a Fenton reaction may have occurred to produce superoxide anion that will require the use of cellular superoxide dismutase to convert to hydrogen peroxide consequently, cellular catalase is depleted (Mishra et al., 1994), due to the utilization of the enzyme in catalyzing hydrogen peroxide to molecular oxygen and water. This depletion of the primary defence apparatus in erythrocyte may have permitted the formation of hydroxyl radicals that perturbed the erythrocyte membrane to increase the formation of malondialdehyde that indicated oxidative stress in mice. The reductions observed in antioxidant enzymes and molecules in non-parasitized mice treated with chloroquine also indicate oxidative stress. This observed marginal stress compared to PnT mice group, could probably have resulted from the activity of the microsomal cytochrome P450 reductase involved in drugs and xenobiotic metabolism in addition to the activity of other superoxide and hydrogen generating systems that operate during normal cellular metabolism.
A previous study failed to reveal any significant changes in biochemical indices examined in the sera of chloroquine treated mice. Histopathological examination however showed cytolysis in hepatocytes in mice group treated with chloroquine by the 12th week of drug administration (Okonkwo et al., 1997). Without observed oxidative stress in liver of non parasitized-chloroquine treated (NPcqT) mice, there are significant (p<0.05) increases in the activity of liver SOD, CAT and reduced G6PD activity compared to PnT mice. These observed increases were not significant (p>0.05) in comparison with control mice group. It may therefore be logical to attribute the non significant nature of the statistical data obtained between control and NPcqT mice to the short treatment duration of 72 h.
The observed significant (p<0.05) increases in liver SOD and CAT of parasitized mice compared to NPcqT and control may perhaps be tied to the fact that the presence of parasites in liver induces hepatic cells superoxide dismutase and catalase activities. This observation may probably be the feasible mechanism for the detoxification of microsomal and the electron transport chain produced superoxides. The mechanism by which parasitemia suppresses the Pentose Phosphate Pathway (PPP) as observed in the reduced activity of G6PD in this study is not known.
A recent report has demonstrated that chloroquine induces oxidative stress in blood at least within six hours after administration (Ogunbayo et al., 2006). This report is contrary to that earlier one reported by Okonkwo et al. (1997) and the finding of this study. This report is therefore unique in that it presents an empirical fact that parasitemia in mice increases oxidative stress in plasma and increases liver SOD and CAT activities while chloroquine does not appear to adversely affect oxidative stress status in mice treated with standard dose.
REFERENCES
- Das, B.S. and N.K. Nanda, 1999. Evidence for erythnocyte lipid peroxidation in acute Falciparum Malaria. J. Trans. Roy. Soc. Trop. Med. Hyg., 93: 58-62.
PubMed - Doumas, B.T., W.A. Watson and H.G. Biggs, 1971. Albumin standards and the measurement of serum albumin with bromcresol green. Clin. Chim. Acta, 31: 87-96.
CrossRefPubMedDirect Link - Gornall, A.G., C.J. Bardawill and M.M. David, 1949. Determination of serum proteins by means of the biuret reaction. J. Biol. Chem., 177: 751-766.
CrossRefPubMedDirect Link - Keller, D.F. and R.W. Kellermeyer, 1970. Erthrocyte glucose-6-phosphate dehydrogenase deficiency a pharmacogenetic prototype. CRC Crit. Rev. Clin. Lab. Sci., 1: 247-302.
CrossRefDirect Link - Kremsner, P.G., B. Greve, B. Lell, D. Luckner and D. Schmid, 2000. Malarial anemia in African children associated with high oxygen-radical production. Lancet, 355: 40-41.
CrossRefDirect Link - Kulkarni, A.G., A.N. Suryakar, A.S. Sardeshmukh and D.B. Rathi, 2003. Studies on Biochemical changes with special reference to oxidant and antioxidant in malaria patients. Indian J. Clin. Biochem., 18: 136-149.
Direct Link - Lege-Oguntoye, L., J.U. Abua, B. Werblinska, W.N. Ogala, A.B. Slotboom and P.F. Olurinola, 1989. Chloroquine resistance of Plasmodium falciparum in semi-immune children in Zaria, Northern Nigeria. Trans. R. Soc. Trop. Med. Hyg., 83: 599-601.
Direct Link - Misra, H.P. and I. Fridovich, 1972. The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. J. Biol. Chem., 247: 3170-3175.
CrossRefPubMedDirect Link - Mishra, N.C., L. Kabilan and A. Sharma, 1994. Oxidative stress and malaria-infected erythrocytes. Indian J. Malariol., 31: 77-87.
PubMedDirect Link - Moron, M.S., J.W. Depierre and B. Mannervik, 1979. Levels of glutathione, glutathione reductase and glutathione S-transferase activities in rat lung and liver. Biochim. Biophys. Gen. Subj., 582: 67-78.
CrossRefPubMedDirect Link - Nosten, F. and P. Brasseur, 2002. Combination therapy for malaria: The way forward? Drug, 62: 1315-1329.
Direct Link - Ogunbayo, O.A., R.A. Adisa, O.G. Ademowo and O.O. Olorunsogo, 2006. Incidence of chloroquine induced oxidative stress in the blood of rabbit. Int. J. Pharmacol., 2: 121-125.
CrossRefDirect Link - Olanrewaju, W.I. and A.W.B.R. Johnson, 2001. Chloroquine-resistance Plasmodium falciparum malaria in Ilorin, Nigeria: Prevalence and risk factors for treatment failure. Afr. J. Med. Sci., 30: 165-169.
Direct Link - Reitman, S. and S. Frankel, 1957. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol., 28: 56-63.
CrossRefPubMedDirect Link - Sarin, K., A. Kumar, A. Prskash and A. Sharma, 1993. Oxidative stress and antioxidant defence mechanism in Plasmodium vivax malaria before and after chloroquine treatment. Indian J. Malariol., 30: 127-133.
PubMed - Sinha, A.K., 1972. Colorimetric assay of catalase. Anal. Biochem., 47: 389-394.
CrossRefPubMedDirect Link - Szaz, G., 1976. A kinetic photometric method for serum gamma-glutamyl transpeptidase. Clin. Chem., 15: 124-136.
PubMedDirect Link - Umar, R.A., S.W. Hassan, M.J. Ladan and M.N. Jiya et al., 2008. Therapeutic efficacy of chloroquine for uncomplicated Plasmodium falciparum malaria in nigeria children at the time of transition to artemisinin based combination therapy. Res. J. Parasitol., 3: 32-39.
CrossRefDirect Link - WHO., 2000. Severe falciparum malaria. Trans. R. Soc. Trop. Med. Hyg., 94: S1-S90.
CrossRefPubMedDirect Link