
Nadia H. Ali
Department of Microbial Chemistry, National Research Center, El-Tahrir Street, Dokki, Cairo, Egypt
Latifa A. Mohamed
Department of Microbial Chemistry, National Research Center, El-Tahrir Street, Dokki, Cairo, Egypt
Asian Journal of Biochemistry
Year: 2008 | Volume: 3 | Issue: 1 | Page No.: 1-10







ABSTRACT
Five strains of Streptomyces were screened for the abilities of their extracts to catalyze the hydrolytic or deaminating activities of purine and pyrimidine ribonucleosides and their bases. These studies are rare in Streptomyces. No hydrolytic cleavage for N-glycosidic bond of nucleosides was observed in all screened strains. Hydrolytic deamination was the only degradative activity occurred with cytidine (as substrate) from the ribonucleosides and their bases tested. Streptomyces hygroscopicus NRRL B-1476 gave the highest level of the hydrolytic deamination of cytidine to uridine. Uridine was chromatographically identified in cell-free extracts. Optimum pH and temperature of the enzyme activity were determined at 7.0 and 50°C, respectively. Thermal stability experiments indicated that the enzyme completely restored its activity at 50°C for 30 min, however a complete loss in enzyme activity was recorded when the enzyme was incubated at 80 and 90°C for 20 and 5 min, respectively. Dialyzed extract caused an increase in enzyme activity of about 55%. Results obtained indicate the involvement of sulfhydryl group(s) in the catalytic site of the enzyme. HgCl2, CuSO4 and FeCl3 (10-2 M) caused a complete inhibition of enzyme activity, whereas, little enzyme activity was retained in presence of AgCl2, MgSO4, BaCl2 and NaCl. Inhibition by uridine was of the competitive type and the enzyme exhibited classic Michaelis Menten saturation kinetics. Its apparent Km and Ki values were found to be 4.16 and 21.9 mM, respectively.
PDF Abstract XML References Citation
How to cite this article
Nadia H. Ali and Latifa A. Mohamed, 2008. Nucleoside Degradation in Some Streptomyces Strains. Asian Journal of Biochemistry, 3: 1-10.
DOI: 10.3923/ajb.2008.1.10
URL: https://scialert.net/abstract/?doi=ajb.2008.1.10
DOI: 10.3923/ajb.2008.1.10
URL: https://scialert.net/abstract/?doi=ajb.2008.1.10
INTRODUCTION
Cytidine deaminase (CDA; EC 3.5.4.5) is an enzyme of the pyrimidine salvage pathways catalyzing the hydrolytic deamination of cytidine and deoxycytidine to the corresponding uridine and deoxyuridine. In bacteria, the enzyme is apparently involved in the salvage pathway of pyrimidines; the catabolic breakdown and recycling of nucleoside compounds. In higher eukaryotes the role and regulation of CDA is less well defined, but it has been extensively studied due to its ability to inactivate several important cytidine-based anticarcinogenic drugs, such as cytosine arabinoside and 5-azacytidine (Emmanuelle et al., 1999). Cytidine deaminase (CDA) purified from human placenta revealed the presence of five isoenzymatic forms that differ only in their isoelectric point. (Vincenzetti et al., 2004). The clinical interest of the human enzyme is due to its capability to deaminate several antitumoral and antiviral cytosine nucleoside analogs, such as the anti-leukemic agent 1-β-D-arabinofuranosylcytosine and the anti-cancer agent 50-azadeoxycytidine, leading to their pharmacological inactivation (De Clerq, 2001). Activation-induced cytidine deaminase (AID) is required for Ig class switch recombination, a process that introduces DNA double-strand breaks in B cells. AID catalyzes cytidine deamination that originates DNA double-strand breaks needed for recombination and it promotes DNA damage response and cell survival (Wu et al., 2005; Krause et al., 2006). The enzymatic mechanisms of deaminating cleavage of nucleosides were developed largely through studies on mammalian tissues, bacteria, yeast and fungi. Cytidine deaminase has been reported from different filamentous fungi namely Aspergillus fumigatus IFO 5840 (Kim and Ha, 1992), A. niger (Elzainy et al., 1989), A. niger NRRL3 (Ali, 1998), A. terricola (Mohamed, 1993), Penicillium citrinum (Allam et al., 1991), P. sizowi (Bezborodov et al., 1974). Ali and Elzainy (2000) reported that extracts of Aspergillus niger NRRL3 catalyzed the hydrolytic deamination of only cytidine from the tested ribonucleotides and ribonucleosides and bases, on the other hand do not catalyzed the cleavage of N-glycosidic linkage of AMP, GMP and UMP and their corresponding nucleosides. Recently Elshafei et al. (2005) found that extracts of Penicillium politans NRC-510 could catalyze the deamination of cytidine to uridine maximally at pH 6.5 and 80°C. Concerning the purification of enzyme many attempts have been made by many authors to purify and characterize cytidine deaminase from Bacillus subtilis ED213 (Park et al., 1994) and Arabidopsis thaliana (Vincenzetti et al., 1999). Pyrimidine base supplementation to the culture medium was found to increase curdlan production by Agrobacterium sp. ATCC 31749 (West, 2006). As far as the authors are aware similar studies were reported only by (Elawarmy and Elzainy, 1985), they studied deamination of cytidine and cytosine in cell-free extracts of Streptomyces viridiviolaceus, so the aim of the study presented was to examine the presence of the enzyme(s) responsible for the degradation of purine and pyrimidine ribonucleosides in some other Streptomyces strains.
MATERIALS AND METHODS
Organisms
Five Streptomyces strains were used throughout this study, namely Str. erythrus NRRL ISP 5517; Str. erythrus NRRL 2338; Str. griseus NRRL B-2682 and Str. hygroscopicus NRRL B-1476. These cultures were obtained from Northern Utilization Research and Development Division, US Department of Agriculture, Peoria, Illinois, USA) and Str. sp. NRC11 was obtained from the Department of Microbial Chemistry, National Research Center, El-Tahrir Street, Dokki, Cairo, Egypt.
Media
Organisms were maintained on solid Czapek-Dox`s medium. The following liquid medium was used throughout this study (g 100 mL-1): glucose 3.0 NaNO3 0.2, K2 HPO4 0.1, MgSO4 7H2O 0.05 and KCl 0.05. The pH value was adjusted at 6.0 before autoclaving.
Cultivation of the Organisms
Spores were scraped and suspended in sterile distilled water. Equal portions of spore suspensions were prepared to inoculate, under aseptic conditions, 250 mL Erlenmeyer flasks, each containing 50 mL of sterile medium. The inoculated flasks were then incubated statically at 28°C for 4 days.
Preparation of Cell-Free Extracts
Mycelia were ground with twice their weight of washed cold sand in a cold mortar. The obtained slurry was centrifuged at 5500 rpm for 10 min and the supernatant was used as the crude enzyme preparation.
Dialysis of Extracts
Dialysis of the extracts was made against 200 volumes of cold 0.02M Tris-HCl buffer pH 7.0 for 2 h at 7°C using dialysis bags (sacks). Dialysis sacks (cellulose tubing 21 mm dia.) were obtained from SIGMA Diagnostic, St Louis, MO 631 78 USA.
Chemical Methods
Ribose was determined by the method of Ashwell (1957). Ammonia was estimated by Nessler`s reagent. Protein was determined by the method of Sutherland et al. (1949).
Enzyme Activity Determination
Purine and pyrimidine ribonucleoside hydrolase was estimated by measuring the ribose formed from the nucleoside by the enzyme action. Aminohydrolase was assayed by measuring the amount of ammonia which appeared after incubation of nucleobases and their nucleosides with the enzyme. This was accompanied by chromatographic identification of the products formed.
Identification of Pyrimidine Nucleoside
Chromatographic identification of the pyrimidine nucleoside was made using chromatographic Whatman No. 3 MM filter paper and two solvent systems. Solvent 1: n-butanol-glacial acetic acid-water (120: 30: 50) and solvent 2: n-butanol-formic acid-water (154: 20: 26) (Simth and Seakins, 1976). The spots were located with an ultraviolet lamp (VL 215 LC, Vilber Lourmat, France).
RESULTS
Screening of Streptomyces Strains
Five strains of Streptomyces (Str. erythrus NRRL ISP 5517; Str. erythrus NRRL 2338; Str. griseus NRRL B-2682; Str. sp. NRC 11 and Str. hygroscopicus NRRL B-1476) were screened for the abilities of their extracts to catalyze the hydrolytic and or the deaminating activities towards some purine and pyrimidine nucleosides and bases. Ribose was not detected in the two sets of reaction mixtures (one set contained purine ribonucleosides, adenosine, guanosine or inosine and the other set contained pyrimidine ribonucleosides cytidine, uridine and pyrimidine deoxy ribonucleoside, thymidine plus the fresh extracts (at pH 4.0, 6.0 and 8.0) of five Streptomyces under study indicating the absence of purine ribonucleoside and pyrimidine ribonucleoside and pyrimidine deoxyribonucleoside hydrolase in these extracts (Table 1), whereas ammonia was detected only in the reaction mixtures that contained cytidine plus extracts of. Str. erythrus NRRL 2338; Str. sp. NRC11 and Str. Hygroscopicus NRRL B-1476 at the same pH values and at the same experimental conditions. On the other hand Str.griseus NRRL B-2682 and Str. erythrus NRRRL ISP5517 could not catalyze the hydrolytic deamination of any nucleosides or their bases (Table 1). On comparing the levels of cytidine deaminase activity in the previously mentioned five Streptomyces strains, it is noted that the enzyme was found in a relatively high amounts in extracts of Str. hygroscopicus NRRL B-1476 as compared with other two organisms (Str. erythrus NRRL 2338; Str. sp. NRC11) so it was selected for further studies.
| Table 1: | Occurrence of deaminating activities in reaction mixtures containing nucleosides or their bases at different pH values in extracts of some Streptomyces strains |
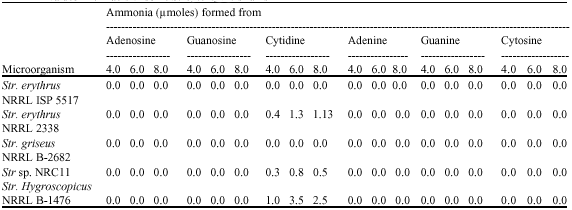 | |
| Reaction mixture contained: nucleoside or base, 5 μmoles; citrate buffer or Tris-HCl, 100 μmoles; extract protein, 1.2 mg; total volume, 1 mL; temperature, 40°C and reaction time, 60 min | |
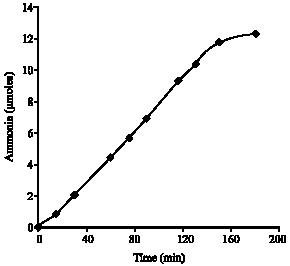 | |
| Fig. 1: | Cytidine deaminase as a function of time of the reaction by extracts of Str. hygroscopicus. Reaction mixture contained: cytidine 15 μmoles; phosphate buffer (pH 6.0), 300 μmoles; extract protein 1.9 mg; total volume, 3 mL; temperature, 40°C and reaction time as indicated |
Chromatographic Identification of the Products
Ammonia was chromatographically identified in reaction mixture that contained cytidine and the fresh cell-free extracts. The developed spots of the identified and authentic uridine had the same Rf values of 0.4 in solvent 1 and 0.25 in solvent 2. The results obtained indicate the presence of hydrolytic deaminating activity in extracts of Str. hygroscopicus NRRL B-1476.
Time of Reaction
Reaction mixtures were incubated for 180 min at 40°C at different incubation periods and after different time intervals aliquots were removed for ammonia determination. Figure 1 shows that there is a gradual increase in the amounts of ammonia formed by cytidine deaminase up to 150 min of incubation, after that there is a steady state occurred in which the increase in incubation time does not reflect noticeable increase in the amounts of ammonia formed from cytidine.
pH Optimum
To determine precisely the optimum pH value(s) at which optimum deamination of cytidine by cell-free extracts of Str. hygroscopicus NRRL B-1476, occurred, a reaction mixtures were made each contained the same amount of protein, the same amount of cytidine and the same amount of buffer. Citrate-phosphate and phosphate buffers were adjusted at pH 3.0 to 8.0. The reaction mixtures were then incubated at 40°C for 60 min after which determination of ammonia was carried out in all of them. Figure 2 shows that extracts of the experimental Str. hygroscopicus NRRL B-1476 strain could catalyze ammonia release from ribonucleoside cytidine over a wide range of pH values. Optimum pH value was recorded at pH 7.0 for the enzymatic deamination of cytidine, while as, enzyme activity at pH 4.0 and 8.0 were recorded about 50 and 83%, respectively after 60 min of incubation.
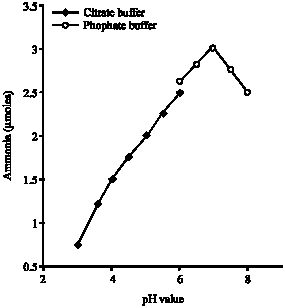 | |
| Fig. 2: | Cytidine deaminase activity as a function of pH value. Reaction mixture contained: cytidine 5.0 μmoles; buffer as indicated, 100 μmoles; extract protein 0.36 mg; total volume, 1 mL; temperature, 40°C and reaction time 60 min |
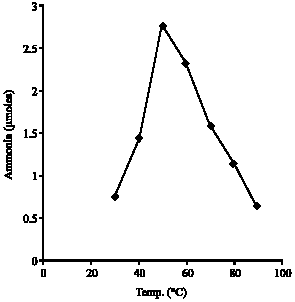 | |
| Fig. 3: | Activity for cytidine of Str. hygroscopicus as a function of temperature. Reaction mixture contained: cytidine 5.0 μmoles; phosphate buffer (pH 6.0), 100 μmoles; extract protein 0.36 mg; total volume, 1 mL; temperature, as indicated and reaction time 60 min |
Temperature Dependence of Cytidine Deaminase Activity
Reaction mixtures containing cytidine as substrate were incubated at different degrees of temperature ranged from 30-90°C for 60 min. Results obtained in Fig. 3 indicated that optimum temperature was obtained at 50°C and a considerable amount of enzyme activity was recorded at 40°C (52%) and 60°C (83.6%) as compared with that obtained at 50°C. Increasing the temperature above 50°C resulted to a gradual decrease in enzyme activity. On the other hand, incubating the enzyme at 80 and 90°C caused loss of cytidine deaminase activity (41.5 and 23.6%, respectively).
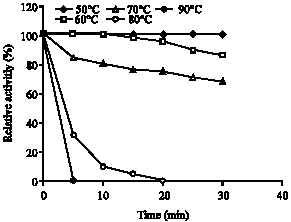 | |
| Fig. 4: | Thermal stability behavior of cytidine deaminase by extracts of Str. hygroscopicus. Reaction mixture contained: cytidine 5.0 μmoles; Tris-HCl buffer (pH 7.0), 100 μmoles; extract protein 1.1 mg; total volume, 1 mL; temperature 50°C and reaction time 60 min |
| Table 2: | Effect of different buffer systems on cytidine ribonucleoside deaminase activity of Str. hygroscopicus NRRL B-1476 |
 | |
| Reaction mixture contained: Cytidine, 5 μmoles; Buffer (pH 7.0), 100 μmoles; Extract protein, 1.43 mg; Total volume, 1 mL; Temperature, 50°C, Reaction time, 60 min | |
Heat Inactivation Kinetics
To test the stability of the enzyme as a function of exposure to 50-90°C in absence of the substrate, aliquots of the exposed crude enzyme preparations were withdrawn at different time intervals, cooled then incubated and assayed for cytidine deaminase activity. Results obtained are graphically represented in Fig. 4, which shows a complete stability of the enzyme activity when incubated at 50°C for 30 min. Gradual inactivation were noticed when the enzyme was incubated at 60 and 70°C, however a complete loss in enzyme activity was recorded when the enzyme was incubated at 80 and 90°C for 20 and 5 min, respectively.
Nature of Buffer
Four reaction mixtures were made at pH 7.0 and received equimolar amount of citrate-phosphate, Tris-acetate, Tris-HCl and phosphate buffers (0.2M). Results obtained in Table 2 indicate that the activity in Tris-HCl buffer is higher than the analogous activities obtained from the other three buffers.
Effect of Freezing and Thawing
In this experiment the crude extracts of cytidine deaminase from Str. hygroscopicus NRRL B-1476 was estimated periodically after three cycles of freezing and thawing during a period of three days at about -5°C, after which it was warmed and an aliquot was withdrawn for assay of activity under the same experimental conditions. It is clear from Table 3 that a slight decrease in enzyme activity about 8% after 1 cycle 24 h, on the other hand at the end of the second and third cycles about 35 and 72% of the activity were lost, respectively.
Dialyzing the Extracts
To find out whether or not cytidine deaminase requires a metal ion(s) in the process of catalysis, crude enzyme extract was prepared. The extract was dialyzed out against 200 volumes of cold 0.02 M tis-HCl buffer at pH 7.0 for 2 h at 7°C. Results obtained indicate that there is an increase of about 55% in enzyme activity upon dialysis. These results can be interpreted by the fact that some inhibitors in the crude extract that affect enzyme activity were dialyzed out.
| Table 3: | Effect of frequent freezing and thawing on cytidine deaminase activity of Str. hygroscopicus NRRL B-1476 |
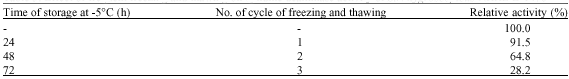 | |
| Reaction mixtures contained: cytidine, 5.0 μmoles; Tris-HCl buffer (pH 7.0); 100 μmoles; extract protein, 0.69 mg; total volume, 1.0 mL; temperature, 50°C and reaction time, 60 min | |
| Table 4: | Effect of some activators and inhibitors on cytidine deaminase activity of Str. hygroscopicus NRRL B-1476 |
 | |
| Reaction mixture contained: cytidine, 5.0 μmoles; Tris-HCl buffer, pH 7.0, 100 μmoles; extract protein, 1.86 mg; compound, 10 μmoles; total volume, 1.1 mL; temperature, 50°C and reaction time, 60 min | |
| Table 5: | Evidence for the involvement of SH group in cytidine deaminase activity |
 | |
Effect of Metal Ions
This experiment was carried out to find out whether or not cytidine deaminase requires a metal ion(s) in the process of catalysis. Different metal ion and other compounds were added to the reaction mixture containing dialyzed extracts each at final concentration of 10-2 M. A control reaction mixture that does not contain any of these salts was made. Results in Table 4 showed that CuSO4, FeCl3 and HgCl2 had a complete inhibitory effect on cytidine deaminase, while as MgSO4, BaCl2 and NaCl reduced activating effect by 50% and the remaining metal salts showed different degrees of inhibition depending upon the type of inhibitor and its final molarity in the reaction mixture. Addition of CoSO4 at 10 mM to the reaction mixture caused about 83% inhibition.
Effect of Sulfhydryl Compounds
To test whether or not SH groups are involved in the catalytic site of cytidine deaminase of Str. hygroscopicus NRRL B-1476 different reaction mixtures were prepared containing reduced glutathione, 2-mercaptoethanol or iodoacetate at a final concentration of 10 mM. Results obtained are cited as relative activities in Table 5 and indicated that the addition of iodoacetate caused a complete inhibition. The inhibitory effect of iodoacetate on cytidine deaminase activity can be interpreted by the fact that iodoacetate alkylates the sulfhydryl group(s). Addition of reduced glutathione and 2-mercaptoethanol at a final concentration of 10 mM to the reaction mixtures had no effect on cytidine deaminase suggesting that free sulfhydryl groups have no role in the activity of the enzyme.
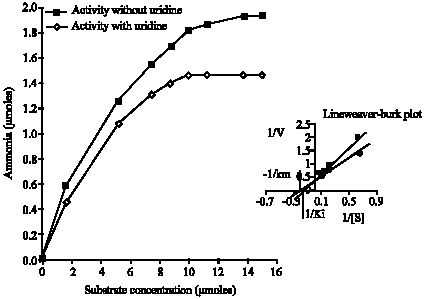 | |
| Fig. 5: | Effect of cytidine concentration on cytidine deaminase from Str. Hygroscopicous NRRL B-1476. Reaction mixture contained (in 1 mL/vol.) Substrate as indicated; extract protein 1.1 mg; Tris-HCl pH 7.0, 100 μmoles; temperature, 50°C and time of reaction 30 min |
Determination of the Apparent Km and Ki Values
Inhibition exerted by uridine seems to be of the competitive type Fig. 5, also demonstrates classic Michaels Menten saturation kinetics. The apparent of Km value for cytidine (under the experimental conditions) and the apparent Ki value for uridine were calculated from the Linweaver Burk plot and found to be 4.16 and 21.9 mM, respectively (Fig. 5). However, the competitive inhibition of cytidine deaminase by uridine leads to the suggestion of the existence of some sort of control mechanisms which does not allow the conversion of all the cytidine present into uridine and hence avoid depletion of the former Dixan and Webb (1964).
DISCUSSION
This study represents an effort to search for the degradation of purine and pyrimidine ribonucleosides and their bases in cell-free extracts of some Streptomyces strains. These studies are rare in Streptomyces strains. In order to obtain this goal, a screening study was made with five Streptomyces cultures for their abilities to catalyze the hydrolytic and or the deaminating activities of some purine and pyrimidine ribonucleosides and their bases. The previous results show that no hydrolytic cleavage of N-glycosidic bond of purine and pyrimidine ribonucleosides in all Streptomyces strains under study, whileas the only activity found in all Streptomyces strains was cytidine deamination, these finding were similar to studies obtained by Elawarmy and Elzainy (1985) in Streptomyces viridiviolaceus. On the other hand in filamentous fungi the enzymes responsible for the degradation of ribonucleosides were known, Ali and Elzainy (2000) reported cytidine deaminase in Aspergillus niger NRRL3, Allam et al. (1981) reported the presence of a pyrimidine ribonucleoside hydrolase that catalyzes of the N-glycosidic bond of cytidine and uridine in Penicillium chrysogenum. He also reported a broad spectrum intracellular constitutive purine and pyrimidine ribonucleoside hydrolase in cell-free extracts of Fusarium moniliforme. On comparing the levels of cytidine deaminase activity in the previously mentioned five Streptomyces strains it is noted that the enzyme was found in a relatively high amounts in extracts of Streptomyces hygroscopicus NRRL B-1476 as compared with the other testing Streptomyces so it was selected for further studies. Studies on the properties of cytidine deaminase of Streptomyces hygroscopicus NRRL B-1476 showed that optimum pH and temperature were found to be at 7.0 and 50°C, respectively, these results was semi similar to results obtained by Elawamry and Elzainy (1985) in Streptomyces viridiviolaceus that had optimal pH and temperature at 6.5-7.5 and 70°C, respectively and in fungi. Bezborodov et al. (1974) reported the presence of cytidine deaminase in P. sizowi was optimum activity at pH values ranging from 5.5-7.0 and optimum temperature of 75°C and Allam et al. (1991) reported that the optimum pH for P. citrinum is 6.5, In addition studies on thermal stability of cytidine deaminase of Streptomyces hygroscopicus NRRL B-1476 indicated that enzyme completely restored its activities at 50 and 60°C for 30 min, however a complete loss in enzyme activity was recorded when the enzyme was incubated at 80 and 90°C for 20 and 5 min, respectively. In contrast the corresponding enzyme reported in P. politans NRC-510 extracts is unique for its thermophilic nature as the enzyme restored its activity at 80°C for 60 min in absence of substrate (Elshafei et al., 2005), in accordance with our results. Dialyzing the extract caused an increase in enzyme activity of about 55%. These results can be explained by the fact that some inhibitors in the crude extracts were dialyzed out. Among the various metal salts tested, CuSO4 and HgCl2 at a concentrations 5x10-3 M and 10-2 M had a complete inhibitory effect on enzyme activity, while CoSO4, MnCl2 and MgCl2 caused a remarkable activating effect. HgCl2 evidenced complete inhibition of cytidine deaminase activity; this fact suggests the possible presence of thiol groups at the catalytic site of the enzyme. The enzyme was affected by sulfhydryl compounds such as reduced glutathione and 2-mercaptoethanol which cause a slight activating effect, whileas the addition of iodoacetate was found to be inhibitory on enzyme activity On the adverse, Allam et al. (1991) reported that SH group(s) might not be involved in catalytic site of that enzyme from P. citrinum. The apparent Km and Ki values for cytidine and uridine were calculated from the Linweaver Burk plot and found to be 4.16 and 21.9 mM, respectively. These results similar to result obtained by Allam et al. (1991). However, the competitive inhibition of cytidine deaminase by uridine leads to the suggestion of the existence of some sort of control mechanisms which does not allow the conversion of all the cytidine present into uridine and hence avoid depletion of the former.
REFERENCES
- Ali, T.H., 1998. Some kinetic studies on cytidine aminohydrolase activity from Aspergillus niger NRRL3. Acta Microbiol. Polonica., 47: 365-372.
Direct Link - Ali, T.A. and T.A. Elzainy, 2000. Hydrolysis of RNA monomers by extracts of Aspergillus niger NRRL. Antonie van Leeuwenhoek. 77: 229-234.
Direct Link - Allam, A.M., T.A. Elzainy, M.A. Elsayed and A.M. Elshafei, 1991. Some kinetic studied onribonucleosides degradation by extracts of Penicillium citrinum. Acta Biotechnol., 11: 227-233.
CrossRef - De Clerq, E., 2001. New development in anti-HIV chemotherapy. Curr. Med. Chem., 8: 1543-1572.
Direct Link - Elshafei, A.M., N.H. Ali and L.A. Mohamed, 2005. Cytidine deaminase activity in Penicillium politans NRC-510. J. Basic Microbiol., 45: 335-343.
CrossRefDirect Link - Emmanuelle, F.N.S., J.M. Grienenberger and J.M. Gualberto, 1999. A cytidine deaminase from Arabidopsis thaliana. Eur. J. Biochem., 236: 896-903.
Direct Link - Krause, K., K.B. Marcu and J. Greeve, 2006. The cytidine deaminases AID and APOBEC-1 exhibit distinct functional properties in a novel yeast selectable system. Mol. Immunol., 43: 295-307.
CrossRefDirect Link - Sutherland, E.W., C.R. Cori, R. Haynes and N.S. Olson, 1949. Purification of the hyperglycemic glycogenolytic factor from insulin and gastric mucosa. J. Boil. Chem., 180: 825-834.
PubMedDirect Link - Vincenzetti, S., A. Cambi, J. Neuhard, K. Schnorr, M. Grelloni and A. Vita, 1999. Cloning, expression and purification of cytidine deaminase from Arabidopsis thaliana. Expression and Purification, 15: 8-15.
PubMedDirect Link - Vincenzetti, S., P.L. Mariani, N. Cammertoni, V. Polzonetti, P. Natalini, B. Quadrini, R. Volpini and A. Vita, 2004. Isoenzymatic forms of human cytidine deaminase. Protein Eng. Design Select., 17: 871-877.
CrossRefDirect Link - West, T.P., 2006. Pyrimidine base supplementation effects curdlan production in Agrobacterium sp. ATCC 31749. J. Basic Microbiol., 46: 153-157.
CrossRefDirect Link - Wu, X., P. Geraldes, J.L. Platt and M. Cascalho, 2005. The double-edged sword of activation-induced cytidine deaminase. J. Immunol., 174: 935-941.
Direct Link