
K. Cheng
Science Research, 205 Hana Road, Edison NJ 08817, USA
C.H. Zou
Science Research, 205 Hana Road, Edison NJ 08817, USA
Asian Journal of Biochemistry
Year: 2007 | Volume: 2 | Issue: 6 | Page No.: 432-436







ABSTRACT
We propose informatics models, using concepts of informative intensity, informative response intensity and informative flux, with different expressions and multiple layers, to describe informative recognitions between a large ribosomal subunit and a complex constructed with a small ribosomal subunit and an initiator tRNA bound on an initiation codon on an mRNA and between base-pairs of a triplet nucleotides (codon) at A site in a ribosome and on an mRNA and a sequence of three complementary nucleotides (anticodon) of a particular tRNA, beyond lengths of chemical bonds, for a natural protein synthesis. In a special expression of (quasi) electrostatics, our model shows the large ribosomal subunit and the complex have a negative informative recognition because they have an attractive force, it is the informative response Intensity to control their relative movements and spatial positions and to make a final decision, for the large ribosomal subunit, to hug, to leave or to ignore the complex. In a special expression of quantum mechanics, our model demonstrates normal complementary base-pairs of the codon and the anticodon have negative Informative Recognitions because their interactive forces are attractive; the other base-pairs have positive informative recognitions because their interactive forces are repulsive. A previous study reported a probability of wrong base-pairs is about 1/10000. With this probability, we estimate a constant barrier height of a potential energy function, the height is about 0.28 eV at a temperature of 37°C.
PDF Abstract XML References Citation
How to cite this article
K. Cheng and C.H. Zou, 2007. Informatics Models of Recognitions of Protein Synthesis. Asian Journal of Biochemistry, 2: 432-436.
DOI: 10.3923/ajb.2007.432.436
URL: https://scialert.net/abstract/?doi=ajb.2007.432.436
DOI: 10.3923/ajb.2007.432.436
URL: https://scialert.net/abstract/?doi=ajb.2007.432.436
INTRODUCTION
Recent years, Bulik et al. (2005) quantified the contribution of defective ribosomal products to antigen production by a model-based computational analysis. Malonga et al. (2006) proposed a model for protein-RNA interaction to study transfer RNA binding to human serum albumin. Whitehead et al. (2006) constructed a physiological response mode to understand cellular responses to gamma radiation, based on integrated analysis of temporal changes in global mRNA and protein abundance along with protein-DNA interactions and evolutionarily conserved functional associations. By analyzing the interactions between micro RNAs and a human cellular signaling network, Cui et al. (2006) found that microRNAs predominantly target positive regulatory motifs, but less frequently target negative regulatory motifs.
However, to our knowledge, questions how a large ribosomal subunit recognize a complex constructed with a small ribosomal subunit and an initiator tRNA bound on an initiation codon on an mRNA and how a triplet nucleotides (codon) at A site in a ribosome and on an mRNA base-pairs a sequence of three complementary nucleotides (anticodon) of a particular tRNA, beyond lengths of chemical bonds, have not been answered in a perspective of informatics.
Present objectives of this study are to develop and propose informatics models of recognitions between a large ribosomal subunit and a complex constructed with a small ribosomal subunit and an initiator tRNA bound on an initiation codon on an mRNA and between base-pairs of a triplet nucleotides (codon) at A site in a ribosome and on an mRNA and a sequence of three complementary nucleotides (anticodon) of a particular tRNA, beyond lengths of chemical bonds, for a natural protein synthesis, based on published molecular biological data (Riddle and Carbon, 1973; Stöffler and Whittman, 1977; Lake, 1985; Alberts et al., 2002) and our published concepts and methods (Cheng and Zou, 2003, 2006, 2007). Therefore, we theoretically answer the above questions in a view of informatics.
MATERIALS AND METHODS
Modeling Informative Recognition (IR) Between a Large Ribosomal Subunit and a Complex Constructed with a Small Ribosomal Subunit and an Initiator tRNA Bound on an Initiation Codon on an mRNA
The protoplasm has been considered as an electrolyte with an uneven charge distribution and the natural cellular electric field has been considered as quasi static in our previous studies (Cheng and Zou, 2003, 2006, 2007). This study was conducted in our institute from 2006 to 2007. We use the same consideration to propose our new models of an Informative Recognition between a complex (an informative sender) constructed with a small ribosomal subunit and an initiator tRNA bound on an initiation codon on an mRNA and a large ribosomal subunit (an informative receiver) beyond lengths of chemical bonds, for a natural protein synthesis (Fig. 1). We estimate a low layer of Informative Intensity (II), at point P (x, y, z) in a Cartesian coordinate system, with a Coulomb's Electric Field Intensity (EFI) that is a multiplication of charge unit dqs (rs), at a point Ps (xs, ys, zs), of a sender, a medium function MEF (r-rs) and a transmitting function TEF (r-rs), where, r is a position vector from the origin to P, rs is a position vector from the origin to Ps in or on the sender. |r-rs| is a distance between P and Ps and ε (r-rs) is a permitivity function of vector r-rs:
| (1) |
| (2) |
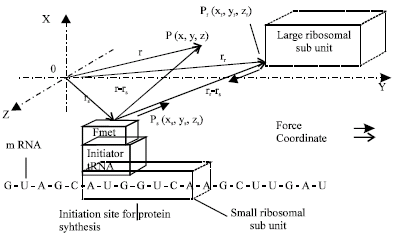 | |
| Fig. 1: | An informative recognition between a large ribosomal subunit and a complex constructed with a small ribosomal subunit and an initiator tRNA bound on an initiation codon on an assumed mRNA in a Cartesian coordinate system. The complex is considered as an informative sender with an assumed net charge Qsn (<0) and the large ribosomal subunit is considered as an informative receiver with an assumed net charge Qrn (>0), respectively. The draw is not in a real scale |
| (3) |
Then, we estimate a high layer of Informative Intensity (II) with a convolution of a production of a medium and a transmitting functions and a total charge distribution Qs (rs) on or in the complex,
| (4) |
Where * means a convolution mathematically. Obviously, an integration of Eq. 4 means an Informative Flux (IF) as well as an Electric Field Flux (EFF).
We estimate a middle layer of Informative Response Intensity (IRI) with an electric field force Fdq (rr), based on Coulomb's law,
| (5) |
Where, dqr (rr) is an integral unit of a receiver's total charge distribution Qr (rr). rr is a position vector from the origin to a point Pr (xr, yr, zr) on or in a large ribosomal subunit (Fig. 1).
Finally, we estimate a high layer of Informative Response Intensity (IRI) in terms of a total electric field force Ft,
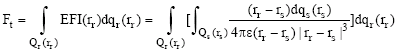 | (6) |
where, |rr-rs| is a distance between Ps and Pr, the distance is greater than a distance of any chemical bond.
Modeling Informative Recognition (IR) Between Codon and Anticodon Base-pairs at A Site in a Ribosome and on an mRNA:
Figure 2 illustrates our quantum mechanics model of Informative Recognition, in a Cartesian coordinate system, when an incoming aminoacyl tRNA (anticodon: AAC) is base-pairing with a codon (UUG) at A site bound with a ribosome that moves along an assumed mRNA template strand in the elongation (+y direction) of a protein synthesis. An amino acid of a peptidyl tRNA at P site has been linked in the peptide chain. We assume the incoming aminoacyl tRNA (an informative receiver) has a quantum wave function Ψ with a free state, the complex (an informative sender) constructed with a codon at A site and a ribosome forms a potential energy function to attract a complementary aminoacyl tRNA and to repulse other aminoacyl tRNA. V and L were defined respectively, for a convenient calculation, as an effective constant height and length of a potential energy function in our previous studies (Cheng and Zou, 2003). We assume L is about a size of a ribosome in x direction (greater than a distance of any chemical bond). When the height is a barrier (V>0), well (V<0) or plain (V = 0), the interactive force F between the incoming aminoacyl tRNA and the complex is repulsive, attractive or zero, respectively and its absolute value is proportional to the height, based on Ehrenfest theorem in quantum mechanics:
| (7) |
Where, V (x) is a potential energy function.
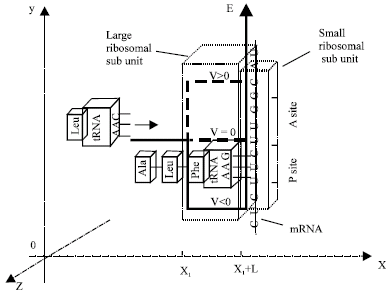 | |
| Fig. 2: | In a cartesian coordinate system, a ribosome and a codon at A site on an assumed mRNA constructs a complex (an informative sender) and a potential energy function V(x) that, respectively attracts (V<0) or repulses (V>0) a complementary or the other aminoacyl tRNA (an informative receiver). Assumed effective constant heights of a barrier (V>0), a plain (V = 0) and a well (V<0) are plotted for a comparison. The drawing is not in a real scale |
RESULTS AND DISCUSSION
We defined a positive, negative or zero Informative Recognition when the interactive force is repulsive, attractive, or zero, respectively (Cheng and Zou, 2006, 2007). Based on the concepts, our modeling results in this paper show a large ribosomal subunit and a complex constructed with a small ribosomal subunit and an initiator tRNA bound on an initiation codon on an mRNA have a negative Informative Recognition, Eq. (5) and (6) involve Newtonian dynamic movements, Eq. (6) makes a final decision, for a large ribosomal subunit, to hug, to leave or to ignore the complex.
According to our quantum mechanics model in this article (Fig. 2 and Eq. 7), normal complementary base-pairs of the codon and the anticodon have negative Informative Recognitions because their interactive forces are attractive; the other base-pairs (not shown in the figure) have positive Informative Recognitions because their interactive forces are repulsive. A previous study reported a probability of wrong base-pairs is about 1/10000 (Kirkwood et al., 1986), We estimate a constant barrier height of a potential energy function of our quantum mechanics model using our published method (Cheng and Zou, 2003), the height is about 0.28 eV at a temperature of 37°C. Therefore, we correlate a constant barrier height of a potential energy function and a probability of a wrong base-pair.
The significant findings of this research are using Coulomb's law to estimate an informative recognition between a large ribosomal subunit and a complex constructed with a small ribosomal subunit and an initiator tRNA bound on an initiation codon on an mRNA and using Ehrenfest theorem to approximately represent informative recognition between base-pairs of the codon and the anticodon, for protein translation. Our modeling results complete our original objectives.
In the special expression of (quasi) electrostatics, we think it is more informative to present an intensity or a force of Coulomb's electric field with a convolution, Eq. 4 and 6, than with a common integration, because it is very convenient to apply a Fourier Transform to obtain some information in frequency domain and it is very elegant to perform signal (informative) processing in future. In the special expression of quantum mechanics, we, respectively consider the wave function Ψ and the gradient of the wave function Ψ of Schrödinger equation as Potential Intensity and Field Intensity of information and matter and their integrations as corresponding Fluxes. We also believe, the principles of our models have general meanings because can be translated in other fields, such as magnetism, acoustics or ultrasonics and hydrodynamics. Moreover, our models in this study combine biochemistry, informatics and physics more organically than our previous works as well as they support from each other.
The limitation of the models is to obtain the distributions of electric charges and permittivities or to measure the data of quantum mechanics in real world environments today.
ACKNOWLEDGMENT
We thank Miss Vivien Cheng for helpful suggestions and comments for this publication.
REFERENCES
- Bulik, S., B. Peters and H.G. Holzhutter, 2005. Quantifying the contribution of defective ribosomal products to antigen production: A model-based computational analysis. J. Immunol., 175: 7957-7964.
Direct Link - Cheng, K. and C.H. Zou, 2003. A three dimensional (3 D) physical model of ribosome movement in protein synthesis. Biomed. Sci. Instrum., 39: 77-82.
Direct Link - Cheng, K. and C.H. Zou, 2006. Informatics and physics models of recognitions of DNA replication and their biomedical applications. Am. J. Applied Sci., 3: 2059-2062.
CrossRefDirect Link - Cheng, K. and C.H. Zou, 2007. Biomedicine and informatics model of Alzheimer`s disease. Am. J. Biochem. Biotechnol., 3: 145-149.
CrossRefDirect Link - Cui, Q., Z. Yu, E.O. Purisima and E. Wang, 2006. Principles of microRNA regulation of a human cellular signaling network. Mol. Syst. Biol., 2: 46-46.
Direct Link - Malonga, H., J.F. Neault and H.A. Tajmir-Riahi, 2006. Transfer RNA binding to human serum albumin: A model for protein-RNA interaction. DNA Cell Biol., 25: 393-398.
Direct Link - Whitehead, K., A. Kish, M. Pan, A. Kaur, D.J. Reiss, N. King, L. Hohmann, J. DiRuggiero and N.S. Baliga, 2006. An integrated systems approach for understanding cellular responses to gamma radiation. Mol. Syst. Biol., 2: 47-47.
Direct Link