
Ozlem Yildirim
Department of Biology, Faculty of Science, University of Ankara,
06100 Tandogan-Ankara, Turkey
Asian Journal of Biochemistry
Year: 2007 | Volume: 2 | Issue: 6 | Page No.: 395-401







ABSTRACT
The glutathione contents, ascorbic acid, nitric oxide, lipid peroxide levels and the activities of free radical scavenging enzymes were examined using five different concentrations of ascorbic acid treated Razaki cultivar and 1616C rootstock. Reduced glutathione was increased with ascorbic acid treatment after seven day in both samples with respect to controls. Similar results were shown in ascorbic acid, nitric oxide and thiobarbituric acid reactive substance levels, while some of these changes were dose dependent. The catalase activity was very low and it increased with increasing ascorbic acid concentration almost in Razaki cultivar and 1616C rootstock. Maximum glutathione peroxidase activity was observed at the end of second day and 10 mg L-1 ascorbic acid treatment for Razaki cultivar. The glutathione peroxidase activity was decreased in 1616C rootstock with respect to controls. Superoxide dismutase activities in these samples were significantly decreased at all concentrations of ascorbic acid and studied times.
PDF Abstract XML References Citation
How to cite this article
Ozlem Yildirim, 2007. Effects of Ascorbic Acid Treatment to Grapevine Rootstock-1616C and Vitis vinifera L. cvs. Razaki. Asian Journal of Biochemistry, 2: 395-401.
DOI: 10.3923/ajb.2007.395.401
URL: https://scialert.net/abstract/?doi=ajb.2007.395.401
DOI: 10.3923/ajb.2007.395.401
URL: https://scialert.net/abstract/?doi=ajb.2007.395.401
INTRODUCTION
It is well defined that free radicals such are superoxide radical, hydroxyl radical, hydrogen peroxide and singlet oxygen have a wide potential for causing cell injury (Lamb and Dixon, 1997). Thus physiological antioxidant enzymes (CAT, GSPOD, SOD etc.) and nonenzymatic (glutathione, vitamin C, vitamin E etc.) scavenger components have been shown to prevent cell injury mediated by free radicals under a variety of conditions in biological systems (Apel and Hirt, 2004; Shao et al., 2005). On the other hand, free radicals also participate in plant growth, development and defense mechanisms (Lamb and Dixon, 1997). Ascorbic acid (vitamin C) scavenges free radicals (reactive oxygen and nitrogen species) and prevent oxidative damage to DNA, lipids and proteins.
Alterations in superoxide dismutase, peroxidase and catalase activities and tissue glutathione concentrations have been reported in plants. For example, in York Imperial apple activity of superoxide dismutase and catalase increases during bud swelling and then decreases during bud break (Wang et al., 1991). The most important reactants in free radical biochemistry in aerobic cells are oxygen and its radical derivatives (superoxide, hydroxyl radical and hydrogen peroxide). Their accumulation is enhanced by several factors as environmental stresses, microbial factors, chemicals, etc. Oxygen radicals are normal metabolites that are inactivated by antioxidant defences. Inactivation and removal of reactive oxygen species depend on reactions involving the antioxidative defence system. Vitamins A, E and C, carotene and glutathione provide antioxidant defences by their ability to exist in reversible oxidized and reduced forms. The enzyme superoxide dismutase (SOD, EC 1.15.1.1) which detoxifies superoxide radical, catalase (CAT, EC 1.11.1.6) and glutathione peroxidase (GSPOD, EC 1.11.1.9); which act on H2O2 and hydroperoxide, respectively, serve as the endogenous antioxidants. Previous experiments have been shown that the oxidative stress reduce the regeneration of grapevine (Vitis vinifera L.) protoplasts (de Marco and Roubelakis-Angelakis, 1996). Also, wounding of leaf tissue and contribution of maceration enzymes were studied to identify the mechanisms of reactive oxygen species generation (Papadakis and Roubelakis-Angelakis, 1999).
Ascorbic acid (vitamin C) is one of the most powerful natural antioxidants. It is water soluble vitamin and generally considered as one of the main activated oxygen species detoxification systems in plant cells. In aqueous solutions, vitamin C also scavenges reactive nitrogen oxide species efficiently, preventing the nitrosation of target molecules. Ascorbate is capable of regenerating tocopherol from the tocopheroxyl radical that is formed upon inhibition of lipid peroxidation by vitamin E. In the presence of free transition metal ions (iron and copper) and ascorbate, the hydroxyl radical can be generated and initiation of lipid peroxidation may occur.
As ascorbate has been shown to be effective against free radicals, present study was to determine whether vitamin C effects the defensive team whose combined purpose is to protect cells from active oxygen damage. For this purpose Razaki cultivar and 1616C rootstock were treated with five different concentrations (1, 5, 10, 50 and 250 mg L-1 ascorbic acid) of ascorbic acid. Glutathione, ascorbic acid, nitric oxide, lipid peroxidation levels and antioxidant enzyme activities (catalase, glutahione peroxidase and superoxide dismutase) were measured in rootstock and cultivar leaves during development. These results were analyzed and the relationship between ascorbate and activated oxygen scavenging enzymes in developing leaves is discussed.
MATERIALS AND METHODS
Materials
All chemicals used were of analytical grade.
Plant Material
This experiment was carried out between March 2002-August 2002 under controlled conditions in the greenhouse at the Department of Horticulture, Faculty of Agriculture, University of Ankara. Plastic pots were used and filled with peat + perlite + air-dry soil with a ratio of 2:1:2, respectively. Four-bud cuttings from 1616C rootstock and Razaki were used and before planting, tree buds from the bottom were removed and planted in plastic pots. When the length of homogen shoots reached to 70 cm ascorbic acid application was started. Plants were watered with 1, 5, 10, 50 and 250 mg L-1 ascorbic acid concentration at the day 0. Tissue collection was performed at the days 2, 4 and 7 from all plants. After the samples were taken, they were frozen in liquid nitrogen and kept at -80°C until use.
Protein Determination and Measurement of Glutathione, Ascorbic Acid, Nitric Oxide and Lipid Peroxidation Levels
The protein content of the samples was measured by the method of Lowry et al. (1951) using bovine serum albumin as a standard.
Glutathione content (GSH) was determined by the procedure of Owens and Belcher (1965) with slight modifications. Freshly collected sample was ground to a fine powder in liquid nitrogen and then freed from proteins by mixing with an equal volume of 3% (w/v) metaphosphoric acid and 30% (w/v) NaCl solution. Deproteinized sample was centrifuged at 3,500 g for 10 min at 4°C. This spectrophotometric procedure measures the change in absorbance at 412 nm occuring when glutathione reduces 5,5-dithio-bis-(2-nitrobenzoic acid).
Tissue Ascorbic Acid (AA) contents were measured in deproteinezed homogenates by the method of Roe and Kuether (1967) in which the colored complex formed was measured spectrophotometrically.
Nitric oxide, as an unstable molecule, that reacts with oxygen and biological molecules to form several end products (e.g., nitrite, nitrate and S-nitrosothiols) that can be determined by different methods. The most preferred method is based on Griess reaction which can be easily applied in the laboratory. The leaf sample was ground to a fine powder in liquid nitrogen and homogenized in 3 volumes of 1.15% KCl at 4°C for 30 sec at 4000 rpm using Ultra Turrax T-25 homogenizer. Homogenized sample (0.5 mL) was incubated with nitrate reductase (EC 1.6.6.2) from Aspergillus sp. (50 mU 100 μL of sample) with NADPH (final concentration, 80 μmol L-1) diluted in 20 mmol L-1 Tris buffer (pH 7.6) for 30 min at room temperature for nitrate reduction. The control sample was analyzed daily with an exogeneous standard (sodium nitrate, 50 μmol L-1). After the reduction, 5% (w/v) ZnSO4 was added for deproteinization. Then this mixture was cetrifuged at 5000xg for 10 min. The samples nitrite levels were measured by Griess reaction (Green et al., 1982). The absorbance at 540 nm was measured and compared to a standard curve constructed using known nitrite concentrations.
Substances that react with thiobarbituric acid were measured as described by Yildirim and Büyükbingöl (2002) using a molar extinction coefficient of 1.56x104 and the results were expressed as nmol TBARS per mg protein.
Enzyme Extraction and Measurement of Free Radical Scavenging Enzyme Activities:
All enzyme extraction operations were performed at 4°C. A 5 g fresh weight samples were ground to a fine powder in liquid nitrogen and homogenized in 15 mL of ice-cold 1M Tris-HCl buffer (pH 8.0) containing 5 mM leupeptin, 1 mM PMSF, 500 mM EDTA, 100 mM MgCl2, 20% (v/v) Triton X-100, 14 μM β-mercaptoethanol and 30% (w/v) insoluble PVP (3x30 sec with 5 min cooling interval) at 8000-10000 rpm. using Ultra Turrax T-25 homogenizer. The homogenate was strained through three layers of miracloth and centrifuged at 14,000xr.p.m for 25 min at 4°C. The resultant supernatant was used as a source of free radical scavenging enzyme activity. Glutathione peroxidase (GSPOD), catalase (CAT) and Superoxide Dismutase (SOD) activities were measured as described (Yildirim and Büyükbingöl, 2002).
Statistical Analysis
All experiments were performed using at least 3 independent experiments and the values were expressed as mean±SD. Data were analyzed using Duncan’s multiple range test in order to detect inter-group differences. A p-value of <0.05 was considered significant.
RESULTS
In freshly harvested samples, glutathione content was significantly (p<0.05) increased at indicated times as compared with respective controls (Table 1). Almost, no significant difference observed in glutathione level between treatments at the end of the fourth and seventh week. In addition, there was no significant difference between rootstock and cultivar at different treatment concentrations (p>0.05). The ascorbic acid content in grape leaves were significantly increase with respective controls at all studied times (p<0.05). Ascorbic acid level increased from 0.19±0.006 to 0.3±0.007 mg g-1 fresh weight in cultivar and from 0.15±0.007 to 0.35±0.009 mg g-1 fresh weight in rootstock at the end of 7th day of the treatment.
As can be seen from Table 2, in the cultivar nitrite levels were increased from 46.6±3.1 to 75.2±3.9 and 48.6±2.6 to 70.3±3.3 mM mg-1 protein at the end of the 2nd and 7th day, respectively. Nitrite levels of 1616C rootstock changed from 44.1±4.3 to 76.6±3.2 and 55.7±3.1 to 75.2±2.5 mM mg-1 protein at the same time as indicated above. As shown in Table 1 and 2, ascorbic acid administration increased nitrite level in 1616C rootstock and Razaki cv.
Levels of TBARS was significantly increased in Razaki cv. and 1616C rootstock samples with respective controls (Table 1 and 2). The TBARS level was significantly increased from approximatelly 13.9±3.1 to 25.4±3.9, 16.1±3.4 to 37.4±2.5 and 15.0±2.6 to 21.2±3.3 nmol mg-1 protein in the Razaki cv. and 17.6±4.3 to 67.5±3.2, 19.2±3.1 to 65.6±3.7 and 13.9±3.4 to 39.7±2.5 nmol mg-1 protein, respectively in 1616 C rootstock (p<0.05).
| Table 1: | Changes in reduced glutathione (nmol g-1 fresh weight), ascorbic acid (mg g-1 fresh weight), nitrite (mM mg-1 protein) and TBARS (nmol mg-1 protein) levels of control and ascorbic acid treated 1616C Rootstock |
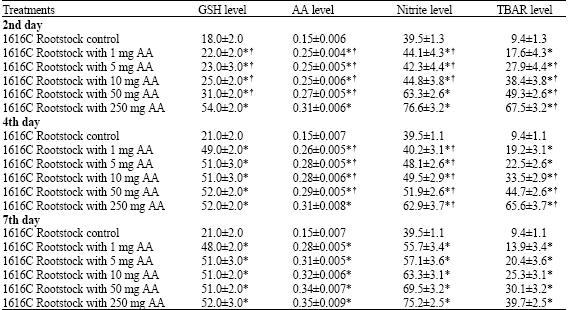 | |
| Results are expressed as means±SD (n = 2 to 4 independent experiments) p-values are shown as *p<0.05 versus respective control groups. †p<0.05 versus seventh day. Other details are given in Materials and Methods | |
| Table 2: | Changes in reduced glutathione (nmol g-1 fresh weight), ascorbic acid (mg g-1 fresh weight), nitrite (mM mg-1 protein) and TBARS (nmol mg-1 protein) levels of control and ascorbic acid treated Razaki cultivars |
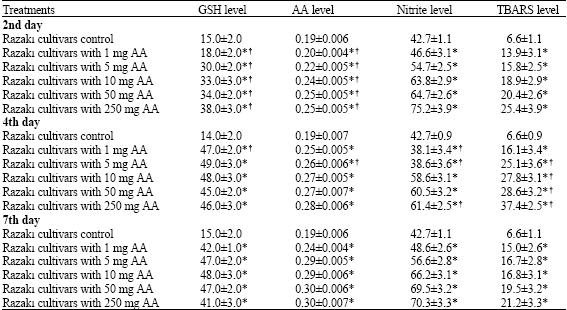 | |
| Results are expressed as means±SD (n = 2 to 4 independent experiments) p-values are shown as * p<0.05 versus respective control groups. †p<0.05 versus 7th day | |
Table 1 and 2 also shows that the TBARS levels of control leaves remained almost unchanged (6.6±1.1 nmol mg-1 protein for Razaki cv. and 9.4±1.2 nmol mg-1 protein for 1616C rootstock). In general, TBARS levels in all leaves increased with increasing concentration of ascorbic acid at the end of the fourth day, then decreased significantly at the end of seventh day (p<0.05).
| Table 3: | Changes in catalase, glutathione peroxidase and superoxide dismutase activities of control and ascorbic acid treated Razaki cultivar and 1616C Rootstock |
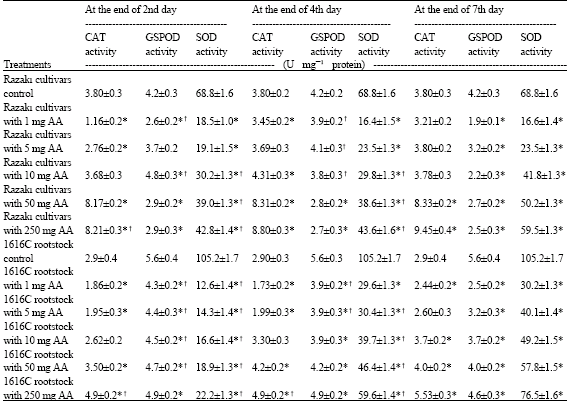 | |
| Results are expressed as means±SD. (n = 2 to 4 independent experiments). p-values are shown as * p<0.05 versus respective control groups. †p<0.05 versus seventh day | |
Although catalase activity was low in these leave of Razaki cv. And 1616C rootstock samples, its activity increases significantly with increasing ascorbic acid concentration (Table 3). The catalase activity was significantly increased from approximatelly 1.16±0.2 to 8.21±0.3, 3.45±0.2 to 8.8±0.2 and 3.21±0.2 to 9.45±0.4x10-4 k-1 sec-1 mg-1 protein at the end of second, fourth and seventh day, respectively in Razaki cultivar (p<0.05). The enzyme activity both in Razaki cv. and 1616C rootstock control leaves remained almost unchanged (3.8±0.3 and 2.9±0.4, respectively). Catalase activity in 1616C rootstock was significantly different at 1 mg and 250 mg ascorbic acid treatment (p<0.05).
The control values for glutathione peroxidase in the grape leaves were 4.2±0.3, 5.6±0.4 U mg-1 protein (Table 3). In 1616C rootstock, GSPOD activity decreased significantly at the end of 7th day as compared to 2nd day up to 250 mg AA treatment. In addition, there were decreases in GSPOD activity in the rootstock at all concentrations of ascorbic acid treatment as compared with controls. On the other hand, treatment with low concentrations of ascorbic acid (1, 5, 10 mgAA L-1) increased GSPOD activity, then the enzyme activity decreased significantly at high concentrations of treatment as compared to control in Razaki cv.
As compared with control samples it was seen that ascorbic acid treatment decreased superoxide dismutase activities from 68.8±1.6 to 18.5±1.0 for 1 mg L-1AA; 19.1±1.5 for 5 mg L-1AA; 30.2±1.3 for 10 mg L-1AA; 39.0±1.3 for 50 mg L-1AA and 42.8±1.4 U mg-1 protein for 250 mg L-1AA at the end of second day in Razaki cv. leaves. Similar results were observed at the end of the fourth and seventh day. At the same time, the SOD activities of 1616C rootstock were 105.2±1.7 U mg-1 protein in control group, 30.2±1.3 for 1 mg L-1AA treatment; 40.1±1.4 for 5 mg L-1 AA treatment; 49.2±1.5 for 10 mg L-1AA treatment; 57.8±1.5 for 50 mg L-1AA treatment and 76.5±1.6 U mg-1 protein for 250 mg L-1AA treatment at the end of 7th day in 1616C rootstock leaves. In general, there was an significant increase (p<0.05) in the activities of SOD when the ascorbic acid concentrations were increased in all studied samples.
DISCUSSION
Based upon the results of the present study, we conclude that Razaki cv. and 1616C rootstock exhibit increased levels of nonenzymatic scavenger components in ascorbic acid treated leaves. Ascorbic acid and H2O2 concentrations reached a maximum, CAT activity declined and SOD activity increased in salicylic acid treated and heat acclimated leaves (Wang and Li, 2006). Ascorbate and glutathione considered as one of the main radical scavenging system in plant cell (Kocsya et al., 2001; Wang and Li, 2006). The changes in glutathione level showed the lowest concentration at the end of second day at the low concentrations (especially at 1, 5 and 10 mg L-1) of ascorbic acid treatment. The elevated GSH content observed in Razaki cv. and 1616C rootstock leaves could maintained protein thiol groups in the reduced state and thereby facilitates repair of oxidative stress induced damage. Accumulation of GSH has been reported during stress conditions may serve as signals to activate transcription factors and gene expression (Kocsya et al., 2001). In glyxysomes and peroxisomes H2O2 detoxified to H2O mainly by catalase, while in other subcellular compartments, ascorbate helps to water conversion of hydrogen peroxide. As expected, ascorbic acid level in leaves were increased with the treatment. However, as the applied ascorbic acid doses increase, no significant increase in ascorbic acid level was observed. The explanation of this situation might be the reduction of H2O2 by ascorbate. Endogenous lipid peroxide level was measured as TBARS which includes end products of lipid peroxidation such as malondialdehyde. We observed that in vivo levels of TBARS were significantly higher in ascorbic acid treated leaves than control groups. It has been reported that glucose oxidation through the pentose phosphate pathway leads to the excessive formation of NADPH, which can promote lipid peroxidation, thus the higher TBARS is not solely the result of peroxidative damage (Pfeifer and McCay, 1976). It was also reported that malondialdehyde content was one main parameter for evaluating membrane oxidation extent (Apel and Hirt, 2004; Shao et al., 2005). Nitric oxide is a small, highly diffusible and short lived molecule. Many reports showed that it has been proven to inhibit foliage expansion, protect plants from oxidative stress (Beligni and Lamattina, 2001). In present study we observed that ascorbic acid administration increased nitric oxide level in 1616C rootstock and Razaki cv.
Present results also show a general tendency of antioxidant enzyme activities to be decreased in ascorbic acid treated Razaki cv. and 1616C rootstock leaves, a pattern that might be indicative of conditions of decreased oxidative stress by treatment. In nature, the catalase activity is very low to detect in grape (Okuda and Yokotsuka, 1999). In the present investigation, it was observed that CAT activities were increased by ascorbic acid treatment but it still very low. O2•– radical can be generated by the plasma membrane NADPH oxidase and it can then form H2O2 by SOD mediated dismutation (Ogawa et al., 1997). GSPOD is one of the defence enzyme that acts on peroxides to remove them. The increase in the activity of CAT and GSPOD in ascorbic acid treated samples may be an adaptive to the changes in the treatment or compensatory mechanism developed to deal with the increased generation of free radicals. The increase membrane permeability and lipid peroxidation and decrease SOD activity was reported for the senescence of tobacco leaves (Dhindsa et al., 1981). Results of this study show a reduction in SOD activity during ascorbic acid treatment.
Taken as a whole, in this study, glutathione, ascorbic acid, nitric oxide, lipid peroxidation levels and three antioxidant enzyme activities were determined in Razaki cv. and 1616C rootstock during seven day in order to examine the relationship between ascorbic acid and free radical scavenging systems. In vivo studies demonstrated that ascorbic acid had an important antioxidant effect. Therefore, it is possible to postulate that antioxidant ascorbic acid can be utilized to protect leaves against oxidative stress.
REFERENCES
- Apel, K. and H. Hirt, 2004. Reactive oxygen species: Metabolism, oxidative stress and signal transduction. Annu. Rev. Plant Biol., 55: 373-399.
CrossRefPubMedDirect Link - Beligni, M.V. and L. Lamattina, 2001. Nitric oxide in plants: The history is just beginning. Plant Cell. Environ., 24: 267-278.
CrossRefDirect Link - De Marco, A. and K.A. Roubelakis-Angelakis, 1996. The complexity of enzymic control of hydrogen peroxide concentration may affect the regeneration potential of plant protoplasts. Plant Physiol., 110: 137-145.
Direct Link - Dhindsa, R.S., P. Plumb-Dhindsa and T.A. Thorpe, 1981. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation and decreased levels of superoxide dismutase and catalase. J. Exp. Bot., 32: 93-101.
CrossRefDirect Link - Green, L.C., D.A. Wagner, J. Glogowski, P.L. Skipper, J.S. Wishnok and S.R. Tannenbaum, 1982. Analysis of nitrate and [15N]in biological fluids. Anal. Biochem., 126: 131-138.
PubMed - Kocsya, G., G. Galibaa and C. Brunoldh, 2001. Role of glutathione in adaptation and signaling during chilling and cold acclimation in plants. Pyhsiol. Plant, 113: 158-164.
Direct Link - Lamb, L. and R.A. Dixon, 1997. The oxidative burst in plant disease resistance. Annu. Rev. Plant Physiol. Plant Mol. Biol., 48: 251-275.
CrossRefPubMedDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Papadakis, A.K. and K.A. Roubelakis-Angelakis, 1999. The generation of active oxygen species differs in tobacco and grapevine mesophyll protoplasts. Plant Physiol., 121: 197-206.
Direct Link - Ogawa, K., S. Kanematsu and K. Asada, 1997. Generation of superoxide anion and localization of CuZn-superoxide dismutase in the vascular tissue of spinach hypocotyls: Their association with lignification. Plant Cell Physiol., 38: 1118-1126.
Direct Link - Okuda, T. and K. Yokotsuka, 1999. Levels of glutathione and activities of related enzymes during ripening of Koshu and Cabernet Sauvignon grapes and during winemaking. Am. J. Enol. Viticul., 50: 264-270.
Direct Link - Owens, C.W.I. and R.V. Belcher, 1965. A colorimetric micro-method for the determination of glutathione. Biochem. J., 94: 705-711.
CrossRefDirect Link - Shao, H.B., Z.S. Liang and M.A. Shao, 2005. Changes of anti-oxidative enzymes and MDA content under soil water deficits among 10 wheat (Triticum aestivum L.) genotypes at maturation stage. Colloids Surf. B: Biointerfaces 45: 7-13.
CrossRefDirect Link - Shao, H.B., L.Y. Chu, G. Wu, J.B. Zang, Z.H. Lu and Y.C. Hu, 2007. Changes of some anti-oxidative physiological indices under soil water deficits among 10 wheat (Triticum aestivum L.) genotypes at tillering stage. Coloids Surf. B. Biointerfaces, 54: 143-149.
CrossRefPubMedDirect Link - Wang, S.Y., H.J. Jiao and M. Faust, 1991. Changes in ascorbate, glutathione and related enzyme activities during thidiazuron-induced bud break apple. Physiol. Plant., 82: 231-236.
Direct Link - Wang, L.J. and S.H. Li, 2006. Thermotolerance and related antioxidant enzyme activities induced by heat acclimation and salicylic acid in grape (Vitis vinifera L.) leaves. Plant Growth Regul., 48: 137-144.
Direct Link - Yildirim, O. and Z. Buyukbingol, 2002. Effects of supplementation with a combination of cobalt and ascorbic acid on antioxidant enzymes and lipid peroxidation levels in streptozotocin-diabetic rat liver. Biol. Trace Elem. Res., 90: 143-154.
Direct Link