
Manju Rawat
Institute of Pharmacy, Pt Ravishankar Shukla University, Raipur (C.G.)-492010, India
Deependra Singh
Institute of Pharmacy, Pt Ravishankar Shukla University, Raipur (C.G.)-492010, India
S. Saraf
Institute of Pharmacy, Pt Ravishankar Shukla University, Raipur (C.G.)-492010, India
Swarnlata Saraf
Institute of Pharmacy, Pt Ravishankar Shukla University, Raipur (C.G.)-492010, India
Asian Journal of Biochemistry
Year: 2007 | Volume: 2 | Issue: 4 | Page No.: 208-223







ABSTRACT
DNA vaccines, also known as genetic or nucleic acid vaccines involve the transfection of cells in vivo by introducing DNA plasmid carrying an antigen-coding gene resulting in an immune response. The purpose of this article is to describe DNA vaccination, to discuss its importance, to review different technologies for improving expression of antigens and to illustrate how it could serve as an alternative for conventional vaccines. DNA vaccines have been prepared against bacterial and viral infections, tumors and allergies and were found to be efficacious in animal studies. These offer number of advantages in terms of ease of administration, manipulation, amplitude and duration of immune response, chemical and biological stabilities and simplicity of manufacture with cost consideration. But, this futuristic alternative needs an elaborate research on its efficacy and associated risks in order to establish the clinical utility of DNA vaccines. This article will explore various aspects of DNA vaccines including its associated advantages, disadvantages, mechanism of action, experimental models studied with a brief overview of second generation vaccines.
PDF Abstract XML References Citation
How to cite this article
Manju Rawat, Deependra Singh, S. Saraf and Swarnlata Saraf, 2007. An Overview of Biochemical Aspects of DNA Vaccines. Asian Journal of Biochemistry, 2: 208-223.
DOI: 10.3923/ajb.2007.208.223
URL: https://scialert.net/abstract/?doi=ajb.2007.208.223
DOI: 10.3923/ajb.2007.208.223
URL: https://scialert.net/abstract/?doi=ajb.2007.208.223
INTRODUCTION
Vaccines are the biological preparations used to induce substantial immune response involving both humoral and cell mediated immunity to provide immune protection against most infective agents like viruses and bacteria. Vaccines usually include killed or attenuated form of the infectious agents, a modified product of infectious agents (toxoid) or a constituent of an infectious agent (such as the capsule) (Weiner and Kennedy, 1999). But, an urgent requirement is to develop vaccines that would be effective for intracellular parasites as well as safer to use. This has led to an increased surge of interest on the research relating to various aspects of vaccines leading to its widespread use.
DNA vaccines are new types of sub-unit vaccines whose delivery into living cells leads to expression of protein of interest in vivo and effective induction of both humoral and cellular immunity. Immunity conferred by DNA vaccines has been shown in many animal models of various diseases including HIV, tuberculosis and cancer (Lee et al., 2004; Ivory and Chadee, 2004; Bartek et al., 2004).
Origin of DNA vaccines dates back to early nineties. Mycobacterial ribosomal and ribonucleic acid preparations were the first nucleic acid preparations reported to be immunogenic (Youmans and Youmans, 1965), which was further supported by immunogenicity of pure RNA extracted from Salmonella typhimurium (Abdelnoor, 2001) But, later RNA was found to serve as adjuvant for the antigen contaminants in the Ribosome/RNA vaccines leading to its unsuitability (Eisenstein, 1975). In 1990, successful delivery and expression of plasmid DNA directly into mouse muscle in vivo was demonstrated (Wolff et al., 1990). The production of anti-human growth hormone (hGH) antibodies and successful induction of both antibody and CTL responses in mice injected with a plasmid DNA encoding hGH and influenza virus protein respectively created the surge of interest in this novel family of vaccines (Tang et al., 1992; Ulmer et al., 1993).
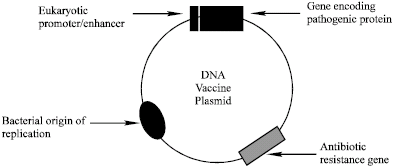 | |
| Fig. 1: | DNA vaccine |
Although a lot of work has been going on relating to development, characterization and clinical trials but major challenge is to demonstrate the clinical utility of DNA vaccines leading to its better and widespread application (Liu, 2003; Liu et al., 2006; Gurunathan et al., 2000; Garmory et al., 2003).
DNA Vaccines
DNA vaccine is a circular double stranded DNA molecule, referred to as a plasmid, containing genes encoding one or more proteins of a pathogen (Fig. 1). Plasmids are extra-chromosomal circular DNA molecules that are often present in multiple copies in bacterial cells. The plasmid contains a bacterial origin of replication by virtue of which it can replicate autonomously inside bacterial cells but not in eukaryotic cells. Such plasmids can be isolated from bacteria using simple and inexpensive protocols. Using recombinant DNA techniques, it is possible to insert a foreign gene into the plasmid molecule to generate a recombinant plasmid. When the gene is inserted downstream of DNA sequences referred to as promoter and enhancer elements to which eukaryotic RNA polymerase II and a host of proteins known as transcription factors bind, then the resultant plasmid is referred to as an eukaryotic expression plasmid.
When such plasmids are introduced into eukaryotic cells, the gene of interest is transcribed by the RNA polymerase II and other accessory proteins resulting in the synthesis of messanger RNA (mRNA) that is translated into the corresponding protein in the cytoplasm of host cells (Davis et al., 1996). In brief, the basic requirements for the backbone of a plasmid DNA vector are a eukaryotic promoter, a cloning site, a polyadenylation sequence, a selectable marker and a bacterial origin of replication (Gurunathan et al., 2000).
Advantages
| • | Plasmid DNA is found to be non-infectious, non-replicating and encodes only the antigen of interest as compared to live attenuated vaccines or viral carrier systems. 11) |
| • | It induces both cell-mediated (Th1 and CTL) and humoral immunity responsible for improved performance of the vaccine (Butts et al., 1998). |
| • | Induction of both immunity pathways effectively target virus-infected cells (Calarota et al., 2001). |
| • | The cell-based immune system offers broad spectrum of action against heterologous strains of a pathogen with longer lasting immunity (Schodel et al., 1994). |
| • | DNA vaccines induce in vivo expression of immunogens thus conserving the native conformation of epitopes (Schodel et al., 1994). |
| • | DNA vaccination can also be utilized for multiple gene expression by conserving an appropriate tertiary conformation for the induction of comformationally specific antibodies and cellular responses by including more than one immunogen gene, thus decreasing the vaccination frequency in children (Grifantini et al., 1998). |
| • | DNA vaccines offer the possibility of generating effective immune responses against diseases where other types of vaccines have failed such as malaria, AIDS and tuberculosis wherein a single antigen alone may not offer complete protection. |
| • | DNA vaccine plasmids can be constructed using simple recombinant DNA techniques and therefore it is possible to co-inoculate multiple plasmids encoding different antigens of same or different pathogens (Spier, 1996; Mahon, 2001). |
| • | Safer to use than live attenuated vaccines especially in immuno compromised hosts (Schodel et al., 1994; Lodmell et al., 1998). |
| • | DNA vaccines can be stored dry or in an aqueous solution at room temperature. DNA vaccines don’t require a cold-chain that constitutes nearly 80% of the cost of vaccinating individuals in developing nations. |
| • | DNA vaccines are stable, easy to freeze dry and reconstitute and can be manufactured inexpensively in large quantities in pure form (Spier, 1996; Lodmell et al., 1998) |
Limitations
| • | DNA vaccination approach can be used to induce immune responses only against the protein components of the pathogen and cannot substitute for polysaccharide based subunit vaccine for e.g., diseases caused by pathogens such as Pneumococcus. |
| • | Although, DNA vaccines has been demonstrated to induce protective immune response but the mechanism of DNA vaccine action is still not completely understood. The potential risks of using plasmid DNA encoding cytokines or co-stimulatory molecule in the host is not yet known. |
| • | DNA vaccines might be associated with the risk of insertional mutagenesis as the plasmid DNA integrates into the genome randomly. If plasmid DNA integrates into the host genome it may either activate oncogens or suppress tumor genes, which may lead to a malignant transformation. |
| • | On intradermal administration of DNA vaccines, the transfected epidermal cells are lost within 10-14 days because of the normal sloughing of keratinized skin tissue whereas on intramuscular administration, the transfected muscle cells remain non-dividing and random insertion is more likely to occur in replicating cells in which DNA is actively being synthesized (Simmonds et al., 1997; Robertson, 1994). |
| • | Another major concern is the induction of anti-DNA antibodies by plasmid DNA. Antibodies to DNA can cause disease and are associated with Systemic Lupus Erythematosus (SLE) (Robertson, 1994). |
| • | The minute amount of antigens produced by transfected cells may induce a state of tolerance of plasmid encoded antigen which appears to persist for long periods of time but, unresponsiveness, rather than protective immunity might result (Simmonds, 1997; Lopez-Macias et al., 1995). |
| • | Potential harmful effects of bacterial ISS (CpG motifs) must also be considered. Schwartz et al. (1997) and Pisetsky (1997) reported that CpG motifs in bacterial DNA caused inflammation in lower respiratory tract. |
Mechanism of Action of DNA Vaccines
General Uptake Mechanism
DNA vaccines are prepared by recombinant DNA technology whereby genes encoding antigenic proteins of a pathogen are inserted into the vector plasmid (Mor and Eliza, 2001; Spier, 1996). The plasmid DNA enters the cell and than into nucleus where it undergoes transcription and translation to express encoded gene product in cytoplasm, which leads to generation of humoral and cell-mediated immune response (Fig. 2).
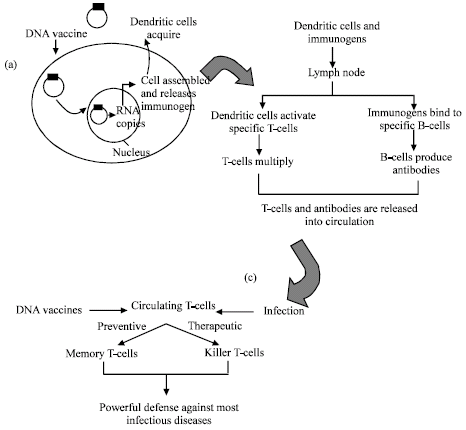 | |
| Fig. 2: | General steps involved in generation of protective immune response by DNA vaccines. (a) Cellular uptake of DNA vaccine (b) Induction of Humoral response (c) Induction of strong cellular response |
Antigen presentation essentially involves proteolytic degradation of the foreign proteins inside the eukaryotic cells and association of the proteolytic fragments with two different types of Major Histocompatibility Complex (MHC) proteins referred to as class I and class II MHC proteins. Class I molecules present protein fragments to cytotoxic T-cells whereas Class II molecules present protein fragments to T-helper cells (Fig. 3).
In case of DNA vaccination, since the pathogenic proteins are synthesized inside the host cells, both humoral and cell-mediated immune responses can be induced. Activation of branches of the immune system requires cytokines produced by TH cells. To ensure carefully regulated activation of TH cells, they can recognize only antigen that is displayed together with class MHC II molecules on the surface of Antigen Presenting Cells (APCs). APCs are specialized cells which express class II MHC molecules on their membranes and deliver a co-stimulatory signal that is necessary for TH cell activation. Antigen Processing/Presenting Cells (APC) degrade a protein antigen into smaller peptides and present the peptide to T-lymphocytes, which eventually are activated. Three cell types are classified as professional APCs: dendritic cells, macrophages and B-lymphocytes. Several other cell types, classified as non-professional APCs: Fibroblasts, thyroid epithelial cells, Glial cells and vascular endothelial cells can be induced to express class II MHC molecules or a co-stimulatory signal.
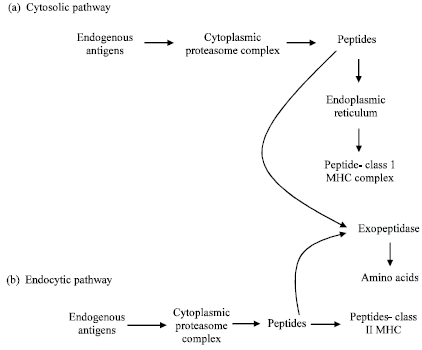 | |
| Fig. 3: | Cytosolic and endocytic pathway for antigen processing (Endogenous antigens are processed in the cytosolic pathway and exogenous antigens are processed in the endocytic pathway) |
A number of studies have been performed to determine the ability of APC to take up plasmid DNA, process it and activate both humoral and cell-mediated branches of immune system. Studies performed by Wolff et al. (1990) and Ulmer et al. (1993 and 1996) suggested that myocytes behaved as APCs on administration of plasmid DNA by intramuscular route but it does not share the later property of APCs. Agadjanyan et al. (1999) showed that muscle cells could induce antigen-specific CTL responses only when the mice were vaccinated with plasmid DNA possessing an antigen and B7 (co-stimulatory molecule) reporter genes. Keratinocytes and/or Langerhans cells might be the APC that are transfected (Klinman et al., 1997).
Later reports indicated that the bone marrow derived dendritic cell is the principal APC involved in plasmid DNA immunization. The plasmid DNA could either transfect somatic cells (myocytes, keratinocytes) or bone marrow derived dendritic cells. When the plasmid DNA is taken up by myocytes or keratinocytes, product encoded by reporter gene is than transferred to dendritic cells by cross priming. Findings by Ulmer et al. (1996) favored this processing mechanism by inducing both antibody and CTL responses to influenza nucleoprotein in mice by transplanting transfected myoblasts. Furthermore isolation of plasmid DNA from lymph node derived and skin derived dendritic cells after intra-muscular or intra-dermal administration, respectively favored direct transfection of bone marrow derived dendritic cells (Casares et al., 1997).
Mechanism of Antigen Processing/presentation
Major Histocompatibility Complex (MHC) class I and class II molecules play major role in presenting a processed antigen to T-lymphocytes. MHC class I molecule are expressed on the surface of practically all nucleated cells (York and Rock, 1996). They consist of two polypeptide chains-an alpha chain and β-2 microglobulin.
The immune system uses two different pathways to eliminate intracellular and extracellular antigens. Endogenous antigens are processed in the cytosolic pathway and presented on the membrane with class I MHC molecules; exogenous antigens are processed in the endocytic pathway and presented on the membrane with class II MHC molecules.
Intracellular proteins such as viral, protozoal, bacterial and tumor proteins are degraded into short peptides by a cytosolic proteolytic system present in all cells. Those proteins targeted for proteolysis often have a small protein called ubiquitin, attached to them. A multi-functional protease complex called a proteasome can degrade ubiquitin-protein conjugates. The proteasomes involved in antigen processing include two sub-units encoded within the MHC gene cluster, LMP2 and LMP7 and a third non-MHC protein LMP10. Transporter proteins (TAP1/TAP2) translocate peptides generated in the cytosol by the proteasome into the Rough Endoplasmic RETICULUM (RER) by a process that requires the hydrolysis of ATP. TAP is optimized to transport peptides that will interact with class I MHC molecules. Molecular chaperones facilitate the folding of poly-peptides in order to form stable class I M HC molecular complex that can exit the RER. The tri-molecular complex (alpha chain, beta-2-microglobulin and peptide) is then transported to the cell outer membrane and presented to the CD8+ CTL. These lymphocytes become fully activated following second signals that include cytokine-receptor (such as IL-2 and IL2R) and co-stimulatory molecule-receptor (such as B7 and CD28) interactions. The activated CD8+ CTL produce perforins that polymerize in the cell membrane of the target cell (e.g., cell infected with the virus) causing its death (York and Rock, 1996).
MHC class II molecules are expressed mainly on APC where they present processed antigenic peptides to TH cells (Mach et al., 1996). MHC class II molecules contain two different polypeptide chains, a 33 kDa alpha chain and a 28-kDa Beta chain, which associate by non-covalent interactions. Each chain in a class II molecule contains two external domains-α1 and α2 domains in one chain and β1 and β2 domains in the other, a transmembrane region and an intracytoplasmic region.
APC internalize exogenous antigens by phagocytosis, endocytosis or both (Watts, 1997; Castellino et al., 1997). Once an antigen is internalized it is degraded into peptides within compartments of the endocytic-processing pathway. Internalized antigen move from early to late endosomes and finally to lysosomes, encountering hydrolytic enzymes and a lower pH in each environment. When class II molecules are synthesized within the RER, three pairs of class II α, β chains associate with the pre-assembled trimer of a protein called invariant chain. This trimeric protein interacts with the peptide-binding cleft of the class II molecules, preventing any endogenously derived protein from binding to the cleft while the class II molecule is within the RER. As the proteolytic activity increases in each successive compartment, invariant chain is gradually degraded. However, a short fragment of the invariant chain termed CLIP (for class II associated invariant chain peptide) remains bound to the class II molecule after the invariant chain has been cleaved within the endosomal compartment. A non-classical class II MHC molecule called HLA-DM is required to catalyze the exchange of CLIP with antigenic peptides. Complex formed is than transported to the plasma membrane and is presented to CD4+T-lymphocytes (T-helpers). These lymphocytes become activated following second signals that include cytokine-receptor and co-stimulatory molecule-receptor interactions. There are at least 2 sub-populations of T-helpers (Th), Th1 and Th2. Activation of either of the two sub-populations predominates depending on the cytokines produced (interleukin (IL)-12 favors the Th1 response and IL-4 favors a Th2 response), the co-stimulatory molecules involved (B7.1-CD28 interaction favors a Th1 response; B7.2-CD28 interaction favors a Th2 response) and the route of entry of the antigen (Romagnani, 1997). Activated Th1 mainly produces IL-2 and gamma-interferon whereas activated Th2 mainly produces IL-4, IL-5, IL-6, IL-10 and IL-13. Cytokines produced in a Th1 response mediate Delayed Type Hypersensitivity, are involved in the activation of CTL, macrophages and NK-cells and help B-lymphocytes to eventually produce IgG2a and IgG3 class antibodies. On the other hand, cytokines produced in a Th2 response promote eosinophil proliferation and production of IgG1 and IgE class antibodies by B-lymphocytes. Maximized Th1 and CTL response are needed for protection against intracellular parasites and tumors.
Some reports indicated that exogenous proteins may be processed, complexed to MHC class I molecules and presented to CTL (Bevan, 1987; Carbone and Bevan, 1990; Kovacsovics et al., 1993; Kovacsovics and Rock, 1995; Shen et al., 1997; Reis et al., 1995; Norbury et al., 1995). By a yet not well-defined mechanism called cross priming, the antigen may be either shuttled directly into the cytosol of the APC, or it may be engulfed by phagocytosis and leaks out of the endosome/lysozome compartment into the cytosol.
DNA Immunization
Route of Administration
Several routes of plasmid DNA inoculation have been studied in animal models. These include intramuscular, sub-cutaneous, intra-periotoneal, intra-dermal, intra-venous, oral, rectal, intra-bursal, intra orbital, intra-tracheal, inta-nasal and vaginal routes (Simmonds et al., 1997; Mor et al., 1995).
In the case of a plasmid DNA vaccine for a tumor, it can be injected into the tumor site (Fuller et al., 1997). The most common routes of administration are by injecting the plasmid DNA dissolved in saline intra-muscularly or intra-dermally using a hypodermic needle or by bombarding plasmid DNA coated onto colloidal gold microparticles in the dermis or muscle using a gene gun.
Effective Dose
The dose used in mice depends on the method of administration. Usually an immune response is generated when 10-100 μg of plasmid DNA is injected and 0.1-1 μg when it is administered with a gene gun (Fynan et al., 1993). Immune response is increased when 1 or 2 boosters are given (Fuller and Haynes, 1994; Fuller et al., 1997; Mustafa et al., 1997; Richmond et al., 1997; Lu et al., 1997). However, time intervals between boosters appear to be critical. In two studies, it has been shown that an increase in the time interval between immunizing doses resulted in an increased immune response (Fuller et al., 1997; Leitner et al., 1997).
Duration of Immune Response
DNA vaccines generally produce long lasting immunity. Akbari et al. (1999) reported that antigen-specific CD4+ T-lymphocytes remained elevated for up to about 10 months following immunization with a DNA vaccine and Gurunathan et al. (2000) reported long-term antigen-specific Th1 activity in mice immunized with a DNA vaccine containing a gene that coded for a Leishmania antigen. CTL responses and antibody levels were observed for up to about 17 months in mice immunized with a DNA vaccine containing a reporter gene coding for an influenza virus protein and a DNA vaccine containing a reporter gene coding for hepatitis B protein, respectively (Raz et al., 1994; Davis et al., 1995).
The observed prolonged duration of the immune response has been reported to be due to the persistence of the antigen produced in the host. Wolff et al. (1992) detected antigen in muscle for up to more than 1 year and influenza virus nucleoprotein (Raz et al., 1994) in the dermis for one month, post plasmid DNA inoculation.
Strength of Immune Response
An optimum immunization regimen using the DNA vaccines involves priming the host and subsequently boosting with the antigen. Letvin et al. (1997) primed rhesus monkeys with plasmid DNA containing HIV env gene. Subsequent boosters were done using a combination of the plasmid DNA and HIV env protein. Strong CTL and neutralizing antibody activity were observed.
Experimental Models of DNA Vaccines
DNA vaccines have been studied in various animal models (Table 1). DNA vaccination strategy resembles virus infection, as the genes transferred by the plasmids require the host cellular machinery to be expressed and thus effective against viral infections.
| Table 1: | Models of DNA vaccines |
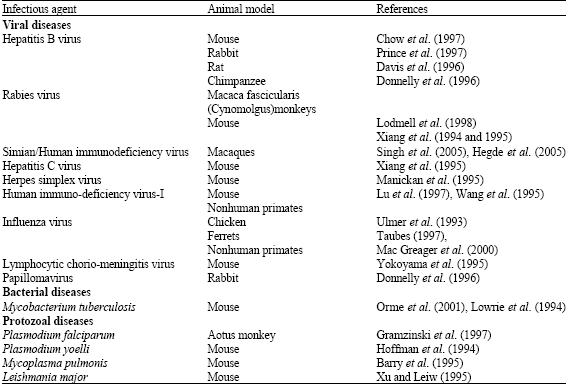 | |
However, genes from other microorganisms have also been used with success. Immunization response for HBV surface antigen in terms of the humoral response in mice mimics, to a certain extent that observed during infection in humans (Boyer et al., 2000). T-cell proliferation and cytokine secretion have been studied in several models and the cytokine profile indicates a Th1 type response characterized by the secretion of interleukin-2 and α-interferon.
The immune response can be remarked long-lasting; however the duration of the immune response does not appear to have any deleterious effects on the animals because they are protected against challenge long after immunization (Xiang et al., 1995). One human trial used plasmid vector expressing HIV-1 genes delivered to HIV-sexopositive persons by intra-muscular injection. This protocol used Marcaine to facilitate DNA uptake, although the mechanism of this effect has not been clearly delineated (Wang et al., 1993). A number of clinical trials have also been initiated since plasmid DNA is now considered an innocuous substance compared with other genetic vectors used in therapy.
Clinical Trials
Vaccines have been mostly studied for infectious diseases and so is the case with DNA vaccines. Immune responses induced by plasmid DNA vaccines depend on means of administration and/or coadministration of an immunomodulator gene. Clinical trials of DNA vaccines are under way for various diseases including cancer, influenza, Hepatitis B, HIV, malaria (Taubes, 1997). But because of the difference between humans and animals in terms of morphology and anatomy further trials are required to be done on humans to confirm its efficacy.
Phase I clinical trails were initiated to evaluate the safety and immunogenicity of HIV-1 env/rev DNA constructs in infected and uninfected persons (Mac Greager et al., 1998). The uninfected persons who received the highest dose of DNA vaccine had antigen specific lymphoproliferative responses and antigen specific production of interferon γ and β-chemokines, but these responses were weak and did not persist (Boyer et al., 2000). In the infected persons, an HIV-1 env/rev DNA vaccine construct boosted the env-specific antibodies; however no consistent effect was observed on cellular responses to HIV. Another phase I clinical trial evaluated HIV regulatory genes, such as rev, nef and tat. Immunization of infected persons with these genes enhanced cellular responses but produced no consistent changes in lymphocytes subsets or viral load (Mac Greager et al., 1998, 2000; Calarota et al., 1998). The DNA vaccines were well tolerated in doses from 20 μg upto 2500 μg; no significant local or systemic reactions were observed and no participant dropped out of the study (Mac Greager et al., 2000; Le et al., 2000).
In the study by Wang et al. (1998) involving phase I clinical trial of malaria DNA vaccines, 20 subjects were vaccinated with plasmid DNA encoding P. falciparum circumsporozoite protein (Reynolds et al., 1981). After immunization 11 of the 20 subjects showed malaria specific, classic cytotoxic T-cells (cytolytic activity restricted by HLA class I antigens) in their blood. Immunization with either a 500 μg dose or a 2500 μg dose of DNA induced a better cellular response than immunization with a 20 μg dose or 100 μg dose of DNA. These results indicated the ability of DNA vaccines to induce specific cellular immunity in humans. But, the protective effects of vaccine against malaria are still not confirmed by above findings. A phase I trial of an experimental HIV vaccine that includes an HIV A subtype gag gene and more than 40 bits of DNA encoding regions of HIV proteins is in progress in Kenya. This is the first component of a prime boost vaccination strategy and will be followed by a second vaccine using modified vaccinia virus as a vector (Reynolds et al., 1981). Another phase I clinical trial was recently started in infected and uninfected person to directly compare vaccines in which the gene is delivered as naked DNA or by attenuated adenovirus (Cohen, 2001).
A DNA vaccine against Hepatitis B virus was evaluated for safety and immunogenicity in a phase I clinical trial involving naïve healthy volunteers. A gene gun was used to propel the DNA into the skin. The hepatitis B DNA vaccine was found to be safe well tolerated and immunogenic (Roy et al., 2000).
Current phase I and II trials are studying DNA vaccines as potential immunotherapies for various cancers, including colon cancer, human follicular lymphoma and cutaneous T-cell lymphoma.
Comparison of DNA Vaccine and Conventional Vaccine
| • | DNA vaccines induce a full spectrum of immune responses in transfected cells. These exhibit freedom from inherent risks of attenuated vaccines reverting to the virulent form. |
| • | DNA vaccines, unlike viral recombinant, can be used repeatedly for different immunogens. |
| • | Conventional vaccines like inactivated or protein vaccines require addition of adjuvants for improved immune response leading to induction of local inflammatory reaction whereas DNA vaccines provide their own adjuvant through unmethlylated Cytosine linked to Guanine (CpG) sequences present in the bacterial part of the vector (Klinman et al., 1999). |
| • | Inactivated vaccines and proteins commonly induce Th2 responses, such as in the case of some infection with respiratory syncitial virus might actually exacerbate disease upon infections whereas DNA vaccines upon intramuscular immunization induce Th1 responses. |
| • | Peptide vaccines induce monospecific B or T cell responses, particularly useful for treatment of cancer, where known point mutation of a self-protein, such as p53 creates a T-cell epitope. Peptides require the addition of adjuvants, especially if the induction of CD8+ T cells is required. Furthermore, peptides are in general poorly immunogenic, which in part reflects their short serum half-life. DNA vaccines carrying mini genes for expression of single epitopes can readily be constructed to replace peptide vaccines. Many of the traditional vaccines fail to induce immune responses in neonates as most traditional vaccines are impaired in the presence of maternally transferred antibodies, whereas DNA vaccines readily stimulate T and B cell responses and when given to neonates are only marginally affected by the mother’s immune status (Table 2). |
| Table 2: | DNA vaccines in comparison with other conventional vaccines |
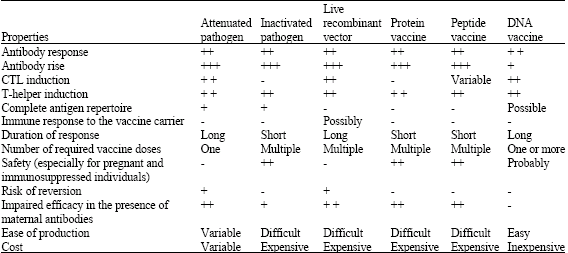 | |
| (+++: Marked, ++: Less marked, +: Least, -: Absent | |
Second Generation Vaccines
As simple plasmids, DNA vaccines effectively induce immune responses and protection in various animal models of disease, but they have induced only modest immune responses in clinical trials. So, the need for even more effective vaccines has led to yet another class of second-generation vaccines. Various strategies have been employed to improve expression of antigens. Strategies may include the incorporation of immunostimulatory sequences in the backbone of the plasmids, co-expression of stimulatory molecules, utilization of localization/secretory signals and utilization of the appropriate delivery system. Optimization of transgene expression is an important consideration. Various strategies involve the modification of regulatory elements such as promoter/enhancers or other transcriptional elements (Chapman et al., 1991; Kevissa et al., 2000). Another element which can be effectively modified includes kozak sequences which are sequences flanking the AUG initiator codon within mRNA influencing recongnition by eukaryotic ribosomes. As a result of studying the conditions required for optimal translational efficiency of expressed mammalian genes, the’ Kozak’ consensus sequence has been shown to be important (Kozak, 1987 and 1997). Codon bias is observed in all species and the use of selective codons in genes often correlates with gene expression efficiency (Zarozinski et al., 1995). Nagata et al. (1999) studied the effect of codon optimization for mammalian cells of Cytotoxic T-Lymphocyte (CTL) epitopes derived from the intracellular bacterium, Listeria monocytogenes and the parasite Plasmodium yoelii and reported that the codon optimisation level of genes correlated well with translational efficiency in mammalian cells.
The greatest deviation from random codon usage in an organism occurs in the highly expressed genes as a result of selection for codons that maximize translational efficiency (Grosjean and Fliers, 1982). A number of other studies have reported that increased immune responses may be obtained by DNA vaccination with a transgene sequence with optimised codon usage (Demil et al., 2001). The backbone of a DNA vaccine vector could be further modified to enhance immunogenicity via the manipulation of the DNA to include certain sequences, so that the DNA itself will have an adjuvantising effect. DNA vaccine vectors contain may CpG motifs (consisting of unmethylated CpG dinucleotides flanked by two 5’ purines and two 3’ pyrimidines) that, overall, induce a Th1-like pattern of cytokine production (Klinman et al., 1997) and are though to account for strong CTL responses frequently seen following DNA vaccination (Krieg et al., 1998). It is possible to augment responses to DNA vaccine vectors by incorporating CpG motifs into the DNA backbone of the plasmid (Weerathna et al., 2001). Alternatively, immune responses may be modulated or enhanced by the co-expression of stimulatory molecules or cytokines (Haddad et al., 2000) or through the use of localisation or secretory signals (Boyle et al., 1997; Lewis et al., 1997; Rice et al., 1999).
Future Prospects
Animal model studies of DNA vaccines have demonstrated success in generating protective immunity not only against viral infections but also against bacterial infection and cancer. But, the major thrust area is the establishment of its safety by means of pre-clinical and then clinical studies, while at the same time considering the benefit of a protective vaccination. As compared to other recombinant protein pharmaceuticals DNA vaccines involve the direct delivery of plasmid DNA itself rather than the protein produced by it as the final product. The potential benefits of DNA vaccination clearly outweigh the associated risks and expeditious approval of DNA vaccine clinical trials will provide important safety information for other applications.
Although an extensive DNA vaccine research has been underway, some researchers are already exploring the idea of using the genome sequence data of several pathogenic microbes to develop genomic vaccines. This approach, also known as expression library immunization, envisages immunization of animals with groups of plasmids encoding 100:1000 genes of a pathogen.
Barry et al. (1995) prepared very small amounts of DNA required with the particle bombardment method. Since a single nanogram of DNA coated on the gold particles induced an immune response, 1 μg can potentially induce 1000 different genes. On this basis a library of gene fragments was prepared from Mycoplasma pulmonis by cloning the genomic DNA into a plasmid expression vector. So, protection against M. pulmonis has been achieved after immunization with different expression libraries (Eisenbraun et al., 1993).
In spite of such an extensive and elaborative research pertaining to DNA vaccine development there are certain key areas such as amount of plasmid DNA to be administered, delivery specification to appropriate antigen presenting cells, antigen expression by DNA vaccines, number of boosters to be given and time interval between boosters need to be optimized. Therefore the major challenge in this field of research is to establish the clinical utility of DNA vaccines for the ultimate benefit of mankind.
REFERENCES
- Agadjanyan, M.G., J.J. Kim, N. Trivedi, D.M. Wilson and B. Monzavi-Karbassi et al., 1999. CD86 (B7-2) can function to drive MHC-restricted antigen-specific CTL responses in vivo. J. Immunol., 162: 3417-3427.
Direct Link - Akbari, O., N. Panjwani, S. Garcia, R. Tascon, D. Lowrie and B. Stockinger, 1999. DNA vaccination: Transfection and activation of dendritic cells as key events for immunity. J. Exp. Med., 189: 169-178.
Direct Link - Bartek, J., C. Lukas and J. Luka, 2004. Checking on DNA damage in s phase. Nat. Rev. Mol. Cell Biol., 5: 792-804.
Direct Link - Barry, M.A., W.C. Lai and S.A. Johnston, 1995. Protection against mycoplasma-infection using expression-library immunization. Nature, 377: 632-635.
Direct Link - Boyer, J.D., A.D. Cohen and S. Vogt, 2000. Vaccination of seronegativevolunteers with a human immunodeficiency virus type 1 env/rev DNAvaccine induces antigen-specific proliferation and lymphocyteproduction of β-chemokines. J. Infect. Dis., 181: 476-483.
Direct Link - Boyle, J.S., C. Koniarasm and A.M. Lew, 1997. Influence of cellular location of expressed antigen on the efficacy of DNA vaccination: Cytotoxic T lymphocytes and antibody respones are sub-optimal when antigen is cytoplasmic after intramuscular DNA immunization. Int. Immunol., 9: 1897-1906.
Direct Link - Butts, C., I. Zubkoff, D.S. Robbins, C. Shui and M. Sarzotti, 1998. DNA immunization of infants: Potential and limitations. Vaccine, 16: 1444-1449.
CrossRef - Carbone, F.R. and M. J. Bevan, 1990. Class I-restricted processing and presentation of exogenous cell-associated antigen in vivo. J. Exp. Med., 171: 377-387.
Direct Link - Calarota, S., G. Bratt and S. Nordlund, 1998. Cellular cytotoxic response induced by DNA vaccination in HIV-1 infected patients. Lancet, 351: 1320-1325.
Direct Link - Calarota, S.A., A. Kjerrstrom, K.B. Islam and B. Wahren, 2001. Gene combination raises broad human immunodeficiency virus-specific cytotoxicity. Hum. Gene Ther., 12: 1623-1637.
Direct Link - Casares, S., K. Inaba, T.D. Brumeanu, R.M. Steinman and C.A. Bona, 1997. Antigen presentation by dendritic cells after immunization with DNA encoding a major histocompatibility complex class II-restricted viral epitope. J. Exp. Med., 186: 1481-1486.
Direct Link - Castellino, F., G. Zhong and R. Germain, 1997. Antigen presentation by MHC class II molecules: Invariant chain function, protein trafficking and the molecular basis of diverse determinant capture. Hum. Immunol., 54: 159-169.
Direct Link - Chapman, B.S., R.M. Thayer, K.A. Vincent and N.L. Haigwood, 1991. Effect of intron A from human cytomegalovirus (Towne) immediate-early gene on heterologous expression in mammalian cells. Nucl. Acids Res., 19: 3979-3986.
Direct Link - Chow, Y.H., W.L. Huang, W.K. Chi, Y. Chu and M.H. Tao, 1997. Improvement of hepatitis B virus DNA vaccines by plasmids coexpressing hepatitis B surface antigen and interleukin-2. J. Virol., 71: 169-178.
Direct Link - Cohen, J., 2001. AIDS research. Merck reemerges with a bold AIDS vaccine effort. Science, 292: 24-25.
CrossRefDirect Link - Davis, H.L., R. Schirmbeck, J. Reimann and R.G. Whalen, 1995. DNA-mediated immunization in mice induces a potent MHC class I-restricted cytotoxic T lymphocyte response to the hepatitis B envelope protein. Hum. Gene Ther., 6: 1447-1456.
Direct Link - Demil, L., A. Bojak, S. Steck, M. Graf, J. Wild, R. Schirmbeck, H. Wolf and R. Wagner, 2001. Multiple Effects of Codon Usage Optimization on Expression and Immunogenicity of DNA Candidate Vaccines Encoding the Human Immunodeficiency Virus Type 1 Gag Protein. J. Virol., 75: 10991-11001.
Direct Link - Donnelly, J.J., D. Martinez, K.U. Jansen, R.W. Ellis, D.L. Montgomery and M.A. Liu, 1996. Protection against papillomavirus with a polynucleotide vaccine. J. Infect. Dis., 173: 314-320.
Direct Link - Eisenbraun, M.D., D.H. Fuller and J.R. Haynes, 1993. Examination of parameters affecting the elicitation of humoral immune responses by particle bombardment-mediated genetic immunization. DNA Cell Biol., 12: 791-797.
Direct Link - Fuller, D.H. and J.R. Haynes, 1994. A qualitative progression in HIV type 1 glycoprotein 120-specific cytotoxic cellular and humoral immune responses in mice receiving a DNA-based glycoprotein 120 vaccine. AIDS Res. Hum. Retrovir., 10: 1433-1441.
Direct Link - Fuller, D.H., M.M. Corb, S. Barnett, K. Steimer and J.R. Haynes, 1997. Enhancement of immunodeficiency virus-specific immune responses in DNA-immunized rhesus macaques. Vaccine, 15: 924-926.
CrossRefDirect Link - Fynan, E.F., R.G. Webster, D.H. Fuller, J.R. Haynes, J.C. Santoro and J.L. Robinson, 1993. DNA Vaccines: Protective immunizations by parenteral, mucosal and gene-gun inoculations. Proc. Natl. Acad. Sci. USA., 90: 11478-11482.
Direct Link - Garmory, H.S., K.A. Brown and R.W. Titball, 2003. DNA vaccines: Improving expression of antigens. Genet. Vacc. Ther., 1: 2-7.
CrossRefDirect Link - Gramzinski, R.A., D.C. Maris, D. Doolan, Y. Charoenvit, N. Obaldia, R. Rossan, M. Sedegah, R. Wang, P. Hobart, M. Margalith and S. Hoffman, 1997. Malaria DNA vaccines in aotus monkeys. Vaccine, 15: 913-915.
Direct Link - Grifantini, R., O. Finco, E. Bartolini, M. Draghi, G.D. Giudice, C. Kocken, A. Thomas, S. Abrignani and G. Grandi, 1998. Multi-plasmid DNA vaccination avoids antigenic competition and enhances immunogenicity of a poorly immunogenic plasmid. Eur. J. Immunol., 28: 1225-1232.
Direct Link - Gurunathan, S., D.M. Klinman and R.A. Seder, 2000. DNA vaccines: Immunology, application and optimization. Annu. Rev. Immunol., 18: 927-974.
CrossRefDirect Link - Haddad, D., J. Ramprakash, M. Sedegah, Y. Charoenvit, R. Baumgartner, S. Kumar, S.L. Hoffman and W.R. Weiss, 2000. Plasmid vaccine expressing granulocyte-macrophage colony-stimulating factor attracts infiltrates including immature dendritic cells into injected muscles. J. Immunol., 165: 3772-3781.
Direct Link - Hegde, R., Z. Liu, G. Mackay, M. Smith, Y. Chebloune, O. Narayan and D.K. Singh, 2005. Antigen expression kinetics and immune responses of mice immunized with non-infectious simian-human immunodeficiency virus DNA. J. Virol., 79: 14688-14697.
Direct Link - Hoffman, S.L., M. Sedegah and R.C. Hedstrom, 1994. Protection against malaria by immunization with a Plasmodium-yoelii circumsporozoite protein nucleic-acid vaccine. Vaccine, 12: 1529-1533.
Direct Link - Ivory, C. and K. Chadee, 2004. DNA vaccines: Designing strategies against parasitic infections. Genet. Vaccines Ther., 2: 17-24.
Direct Link - Kevissa, M., Von J. Kampen, R. Zurbriggen, R. Gluck, J. Reimann and R. Schirmbeck, 2000. Efficient vaccination by intradermal or intramuscular inoculation of plasmid DNA expressing hepatitis B surface antigen under desmin promoter/enhancer control. Vaccine, 18: 2337-2344.
Direct Link - Klinman, D.M., G. Yamshchikov and Y. Ishigatsubo, 1997. Contribution of CpG motifs to the immunogenicity of DNA vaccines. J. Immunol., 158: 3635-3639.
Direct Link - Klinman, D.M., K.M. Barnhart and J. Conover, 1999. CpG motifs as immune adjuvants. Vaccine, 17: 19-25.
CrossRefPubMedDirect Link - Kovacsovics, B.M., K. Clark, B. Banacerraf and K.L. Rock, 1993. Efficient major histocompatibility complex class I presentation of exogenous antigen upon phagocytosis by macrophages. Proc. Natl. Acad. Sci. USA., 90: 4942-4946.
Direct Link - Kovacsovics, B.M. and K.L. Rock, 1995. Phagosome-to-cytosol pathway for exogenous antigens presented on MHC class I molecules. Science, 267: 243-246.
CrossRefDirect Link - Kozak, M., 1987. At least six nucleotides preceding the AUG initiator codon enhance translation in mammalian cells. J. Mol. Biol., 196: 947-950.
CrossRefPubMedDirect Link - Kozak, M., 1997. Recognition of AUG and alternative initiator codons is augmented by G in position +4 but is not generally affected by the nucleotides in positions +5 and +6. EMBO J., 16: 2482-2492.
CrossRefDirect Link - Krieg, A.M., A.K. Yi, J. Schorr and H.L. Davis, 1998. The role of CpG dinucleotides in DNA vaccines. Trends Microbiol., 6: 23-27.
CrossRefPubMedDirect Link - Le, T.P., K.M. Hedstrom, Y. Charoenvit, M. Sedegah and J.E. Epstein, 2000. Safety, tolerability and humoral immune responses after intramuscular administration of a malaria DNA vaccine to healthy adult volunteers. Vaccine, 18: 1893-1901.
Direct Link - Lee, D., B.S. Graham, Y.L. Chiu, P.B. Gilbert and M.J. McElrath et al., 2004. Breakthrough infections during Phase 1 and 2 Prime-Boost HIV-1 vaccine trials with canarypox vectors (ALVAC) and Booster dose of recombinant gp120 or gp160. J. Infect. Dis., 190: 903-907.
Direct Link - Leitner, W.W., M.C. Seguin, W.R. Ballou, J.P. Seitz, A.M. Shultz, M.J. Sheehy and J. Lyon, 1997. Immune responses induced by intramuscular or gene gun injection of protective deoxyribonucleic acid vaccines that express the circumsporozoite protein from Plasmodium berghei malaria parasites. J. Immunol., 159: 6112-6119.
Direct Link - Letvin, N.L., D.C. Montefiori, Y. Yasutomi, H.C. Perry and M.E. Davies et al., 1997. Potent, protective anti-HIV immune responses generated by bimodal HIV envelope DNA plus protein vaccination. Proc. Natl. Acad, Sci. USA., 94: 9378-9383.
Direct Link - Lewis, P.J., G.J.M. Cox, L. Van Drunen, S. Hurk and L.A. Babiuk, 1997. Polynucleotide vaccines in animals: Enhancing and modulating responses. Vaccine, 15: 861-864.
Direct Link - Liu, M.A., B. Wahren and G.B.K. Hedestam, 2006. DNA vaccines: Recent developments and future possibilities. Hum. Gene Ther., 17: 1051-1061.
Direct Link - Lodmell, D.L., N.B. Ray, M.J. Parnell, L.C. Ewalt, C.A. Hanlon, J.H. Shaddock, D.S. Sanderlin and E. Charles, 1998. DNA immunization protects nonhuman primates against rabies virus Rupprecht. Nat. Med., 4: 949-952.
CrossRefDirect Link - Lopez-Macias, C., M.A. Lopez-Hernandez, C.R. Gonzalez, A. Isibasi and V. Ortiz-Navarrete, 1995. Induction of antibodies against Salmonella typhi OmpC porin by naked DNA immunization. Ann. N. Y. Acad Sci., 772: 285-288.
Direct Link - Lowrie, D.B., R.E. Tascon, M.J. Colston and C.L. Silva, 1994. Towards a DNA vaccine against tuberculosis. Vaccine, 12: 1537-1540.
Direct Link - Lu, S., K. Manson, M. Wyand and H.L. Robinson, 1997. SIV DNA vaccine trial in macaques: Post-challenge necropsy in vaccine and control groups. Vaccine, 15: 920-923.
Direct Link - Mac-Greager, R.R., J.D. Boyer, K.E. Ugen, K.E. Lacy, S.J. Gluckman and M.L. Bagarazzi, 1998. First human trial of a DNA-based vaccine for treatment of human immunodeficiency virus type 1 infection: Safety and host response. J. Infect. Dis., 178: 92-100.
Direct Link - Mac-Greager, R.R., J.D. Boyer, R.B. Ciccarelli, R.S. Ginsberg and D.B. Weiner, 2000. Safety and immune responses to a DNA-based Human Immunodeficiency Virus (HIV) type I env/rev vaccine in HIV-infected recipients: Follow-up data. J. Infect. Dis., 181: 406-414.
Direct Link - Mach, B., V. Steimle, E. Martinez-Soria and W. Reith, 1996. Regulation of MHC class II genes: Lessons from a disease. Ann. Rev. Immunol., 14: 301-331.
Direct Link - Mahon, B.P., 2001. The Rational design of vaccine adjuvants for mucosal and neonatal immunization. Curr. Med. Chem., 8: 1057-1075.
Direct Link - Manickan, E., R. Rouse, Z.Y. Yu, W.S. Wire and B.T. Rouse, 1995. Genetic immunization against herpes-simplex-virus protection is mediated by CD4 (+) T-lymphocytes. J. Immunol., 155: 259-265.
Direct Link - Mor, G., D.M. Klinman, S. Shapiro, E. Hagiwara, M. Sedegah and J.A. Norman, 1995. Complexity of the cytokine and antibody response elicited by immunizing mice with Plasmodium yoelii circumsporozoite protein plasmid DNA. J. Immunol., 155: 2039-2046.
Direct Link - Mor, G. and M. Eliza, 2001. DNA Vaccines: Immunology, tolerance and autoimmunity. Mol. Biotechnol., 19: 245-250.
Direct Link - Mustafa, F., J.F. Richmond, R. Fernandez-Lorsson, S. Lu and R. Fredriksson et al., 1997. HIV-1 Env glycoproteins from two series of primary isolates: Replication phenotype and immunogenicity. Virology, 229: 269-278.
Direct Link - Nagata, T., M. Uchijima, A. Yoshida, M. Kawashima and Y. Koide, 1999. Codon optimization effect on translational efficiency of DNA vaccine in mammalian cells: Analysis of plasmid DNA encoding a CTL epitope derived from microorganisms. Biochem. Biophys. Res. Commun., 261: 445-451.
Direct Link - Norbury, C.C., L.J. Hewlett, A.R. Prescott, N. Shastri and C. Watts, 1995. Class I MHC presentation of exogenous soluble antigen via macro-pinocytosis in bone marrow macrophages. Immunity, 3: 783-791.
Direct Link - Orme, I.M., D.N. McMurray and J.T. Belisle, 2001. Tuberculosis vaccine development: Recent progress. Trends Microbiol., 9: 115-118.
CrossRefDirect Link - Pisetsky, D.S., 1997. Immunostimulatory DNA: A clear and present danger? Nat. Med., 3: 829-829.
CrossRefDirect Link - Prince, A.M., R.G. Whalen and B. Brotman, 1997. Successful nucleic acid based immunization of newborn chimpanzees against hepatitis B virus. Vaccine, 15: 916-919.
Direct Link - Raz, E., D.A. Carson, S.E. Parker, T.B. Parr and A.M. Abai et al., 1994. Intradermal gene immu nization: The possible role of DNA uptake in the induction of cellular immunity to viruses. Proc. Natl. Acad. Sci. USA., 91: 9519-9523.
Direct Link - Reis, C., E. Sousa and R.N. Germain, 1995. Major histocompatibility complex class I presentation of peptides derived from soluble exogenous antigen by a subset of cells engaged in phagocytosis. J. Exp. Med., 182: 841-851.
PubMed - Reynolds, F.H., G.J. Todaro, C. Fryling and J.R. Stephenson, 1981. Human transforming growth factors induce tyrosine phosphorylation of EGF receptors. Nature, 292: 259-262.
CrossRef - Rice, J., C.A. King, M.B. Spellerberg, N. Fairweather and F.K. Stevenson, 1999. Manipulation of pathogen-derived genes to influence antigen presentation via DNA vaccines. Vaccine, 17: 3030-3038.
CrossRef - Richmond, J.F., F. Mustafa, S. Lu, J.C. Santoro, J.C. Weng and M. O'Connell et al., 1997. Screening of HIV-1 Env glycoproteins for the ability to raise neutralizing antibody using DNA immunization and recombinant vaccinia virus boosting. Virology, 230: 265-274.
Direct Link - Robertson, J.S., 1994. Safety considerations for nucleic acid vaccines. Vaccine, 12: 1526-1528.
Direct Link - Roy, M.J., M.S. Wu and L.J. Barr, 2000. Induction of antigen-specific CD8+ T cells, T helper cells and protective levels of antibody in humans by particle-mediated administration of a hepatitis B virus DNA vaccine. Vaccine, 19: 764-778.
Direct Link - Schwartz, D.A., T.J. Quinn, P.S. Thorne, S. Sayeed, A.K. Yi and A.M. Krieg, 1997. CpG motifs in bacterial DNA cause inflammation in the lower respiratory tract. J. Clin. Invest., 100: 68-73.
CrossRefDirect Link - Shen, Z., G. Reznikoff, G. Dranoff and K.L. Rock, 1997. Cloned dendritic cells can present exogenous antigens on both MHC class I and class II molecules. J. Immunol., 158: 2723-2730.
Direct Link - Simmonds, R.S., M.H. Shearer and R.C. Kennedy, 1997. DNA vaccine-from principle to practice. Parasitol. Today, 13: 328-331.
Direct Link - Singh, D.K., Z. Liu, D. Sheffer, G.A. Mackay, M. Smith, S. Dhillon, R. Hegde, F. Jia, I. Adany and O. Narayan, 2005. A non-infectious SHIV DNA vaccine that protects macaques against AIDS. J. Virol., 79: 3419-3428.
Direct Link - Tang, D.C., M. Devit and S.A. Johnston, 1992. Genetic immunization is a simple method for eliciting an immune response. Nature, 356: 152-154.
Direct Link - Ulmer, J.B., J.J. Donnelly, S.E. Parker, G.H. Rhodes and P.L. Felgner et al., 1993. Heterologous protection against influenza by injection of DNA encoding a viral protein. Science, 259: 1745-1749.
CrossRefDirect Link - Ulmer, J.B., R.R. Deck, C.M. Dewitt, J.I. Donnelly and M.A. Liu, 1996. Generation of MHC class I-restricted cytotoxic T lymphocytes by expression of a viral protein in muscle cells: Antigen presentation by non-muscle cells. Immunology, 89: 59-67.
Direct Link - Wang, B., J. Boyer, V. Srikantan, K. Ugen, L. Gilbert, C. Phan, K. Dang, M. Merva, M.G. Agadjanyan and M. Newman, 1995. Induction of humoral and cellular immune responses to the human immunodeficiency type 1 virus in nonhuman primates by in vivo DNA inoculation. Virology, 211: 102-112.
CrossRef - Wang, R., D.L. Doolan, T.P. Le, R.C. Hedstrom and K.M. Coonan et al., 1998. Induction of antigen-specific cytotoxic T lymphocytes in humans by a malaria DNA vaccine. Science, 282: 476-480.
CrossRefPubMedDirect Link - Watts, C., 1997. Capture and processing of exogenous antigens for presentation on mhc molecules. Ann. Rev. Immunol., 15: 821-850.
Direct Link - Weerathna, R.D., T. Wu, S.M. Efler, L. Zhang and H.L. Davis, 2001. Designing gene therapy vectors: Avoiding immune responses by using tissue-specific promoters. Gene Ther., 8: 1872-1878.
Direct Link - Wolff, J.A., R.W. Mallone, P. Williams, W. Chong, G. Acsadi, A. Jani and P.L. Felgner, 1990. Direct gene transfer into mouse muscle in vivo. Science, 247: 1465-1468.
Direct Link - Xiang, Z.Q., S. Spitalnik, M. Tran, W.H. Wunner, J. Cheng and H. Ertl, 1994. Vaccination with a plasmid vector carrying the rabies virus glycoprotein gene induces protective immunity against rabies virus. Virology, 199: 132-140.
Direct Link - Xiang, Z.Q., S.L. Spitalnik, J. Cheng, J. Erikeson, B. Wojczyk and H.C. Ertl, 1995. Immune responses to nucleic acid vaccines to rabies virus. Virology, 209: 569-579.
Direct Link - Xu, D. and F.Y. Liew, 1995. Protection against leishmaniasis by injection of DNA encoding a major surface glycoprotein, gp63, of L. major. Immunology, 84: 173-178.
Direct Link - Yokoyama, M., J. Zhang and J.L. Whitton, 1995. DNA immunization confers protection against lethal lymphocytic choriomeningitis virus infection. J. Virol., 6964: 2684-2688.
Direct Link - York, I.A. and K.L. Rock, 1996. Antigen processing and presentation by the Class-I major histocompatibility complex. Annu. Rev. Immunol., 14: 369-396.
Direct Link - Youmans, G.P. and A.S. Youmans, 1965. Immunogenic activity of a ribosomal fraction obtained from Mycobacterium tuberculosis. J. Bacteriol., 89: 1291-1298.
Direct Link - Zarozinski, C.C., E.F. Fynan, L.K. Selin, H.L. Robinson and R.M. Welsh, 1995. Protective CTL-dependent immunity and enhanced immunopathology in mice immunized by particle bombardment with DNA encoding an internal virion protein. J. Immunol., 154: 4010-4017.
Direct Link