
Naglaa M. Mohamed
Department of Biochemistry, Faculty of Pharmacy, Suez Canal University, Ismailia, Egypt
Russell T. Hill
Center of Marine Biotechnology, University of Maryland Biotechnology Institute, Baltimore, Maryland, USA
Raouf W. Kilada
Department of Marine Science, Faculty of Science, Suez Canal University, Ismailia, Egypt
Sherief I. Khalifa
Department of Pharmacology, Faculty of Pharmacy, Suez Canal University, Ismailia, Egypt
Soad H. Abou-El-Ela
Department of Biochemistry, Faculty of Pharmacy, Suez Canal University, Ismailia, Egypt
Asian Journal of Biochemistry
Year: 2006 | Volume: 1 | Issue: 4 | Page No.: 338-342







ABSTRACT
Morphologically, two species of giant clams Tridacna have been recognized in the Red Sea, T. squamosa and T. maxima. Similar morphology among species makes classification difficult. For proper classification, we sequenced an approximately 450-nucleotide fragment of the mitochondrial 16S rDNA gene from the tissues of Tridacna from eight locations in the Red Sea. Our results suggest that there are three novel species of Tridacna in the Red Sea. Two of these species are related to T. maxima and one is related to T. squamosa. Of the two species related to T. maxima, one species was found in Hurghada, Marsa Ghaleb and Safaga while the other was found in Abu Zenima, Abou Galum, Dahab and Nuweiba. All three novel species were found in Ras Mohamed. Our results do not support the morphological classification that suggests the existence of only two tridacnid species, T. maxima and T. squamosa in the Red Sea.
PDF Abstract XML References Citation
How to cite this article
Naglaa M. Mohamed, Russell T. Hill, Raouf W. Kilada, Sherief I. Khalifa and Soad H. Abou-El-Ela, 2006. Molecular Genetic Analysis of Giant Clam (Tridacna sp.) Populations in the Northern Red Sea. Asian Journal of Biochemistry, 1: 338-342.
DOI: 10.3923/ajb.2006.338.342
URL: https://scialert.net/abstract/?doi=ajb.2006.338.342
DOI: 10.3923/ajb.2006.338.342
URL: https://scialert.net/abstract/?doi=ajb.2006.338.342
INTRODUCTION
Among the different species of invertebrates colonizing the coral reefs in the Indo-Pacific and Red Sea areas are the giant clams of the genus Tridacna, the largest and fastest growing living members of the class Bivalvia (Yonge, 1980). They are highly morphologically derived cardiids that have evolved an obligate symbiotic association with photosynthetic dinoflagellate algae of the genus Symbodium, living in their enlarged siphonal mantle (Schneider, 1998).
Rosewater (1965) recognized six living species of giant clams: Hippopus hippopus, Tridacna gigas, T. derasa, T. maxima, T. squamosa and T. crocea. Later, Rosewater (1982) recognized Hippopus porcellanus and Benzie and Williams (1998), using electrophoretically detectable allozyme variation, confirmed that the two Hippopus species were genetically differentiated. Lucas et al. (1990) subsequently described T. tevoroa and suggested that it may represent a transitional species between Hippopus and Tridacna (Lucas et al., 1991). Therefore, the number of known giant clam species has increased from six to eight.
Tridacnins, the major protein isolated from Tridacna, has been found to react with various cell types, including red blood cells, lymphocytes, spermatozoa and tumor cells of different origins (Uhlenbruck et al., 1975, 1978). Furthermore, Tridacnin from T. maxima and T. squamosa showed stronger effects than tridacnin from T. derasa and T. gigas (Uhlenbruck et al., 1978). Recently, 24-methylenecholesterol was isolated from the Red Sea Tridacna mantle and was found to possess modest cytotoxic activity (Kilada, 2001).
Along the coasts of the Red Sea, two living tridacnid species have been recognized morphologically: T. maxima and T. squamosa (Mansour,1946a-c, 1949; Hughes, 1977; Mergner and Mastaller, 1980; Kilada et al., 1998). However, classification based on morphological structure makes it difficult to recognize proper species. Therefore, molecular classification based on comparing the sequence of mitochondrial DNA may offer new insights for a more precise classification of the genus Tridacna in the Red Sea.
We sequenced a 450 nucleotide fragment of mitochondrial, large subunit ribosomal gene (16S) from the tissues of the two species of Tridacna from different locations in the northern part of the Red Sea.
MATERIALS AND METHODS
Collection of Clam Specimens
Tridacnid clams were collected from eight locations in the Egyptian Red Sea during the spring of 2002; four in the Gulf of Aqaba-Dahab (DH), Nuweiba (NU), Abou Galum (AG) and Ras Mohamed (RM); three in the main basin of the Red Sea-Hurghada (HU), Safaga (SF) and Marsa Ghaleb (MG) and one location in the Gulf of Suez-Abu Zenima (AZ) (Fig. 1). The specimens were brought alive to the Department of Biochemistry, Faculty of Pharmacy, Suez Canal University where the animals were removed from their shells and dissected. For each specimen, a portion of the adductor muscle tissue was excised with a razor blade and frozen at -80°C. The rest of the organs were separated and frozen at -80°C and the shells were saved.
Extraction of DNA from the Clams
DNA was extracted exclusively from adductor muscle tissue from the specimens of all tridacnids. Extraction was limited to the adductor muscle tissue because symbiotic dinoflagellate zooxanthellae are found within the mantle and internal organ tissues. Roughly 200 to 300 mg of tissue was diced finally with a razor blade and the DNA was extracted using a QIAGEN DNeasy™ tissue extraction kit according to the manufacturer’s instructions (QIAGEN Gmbh, Germany).
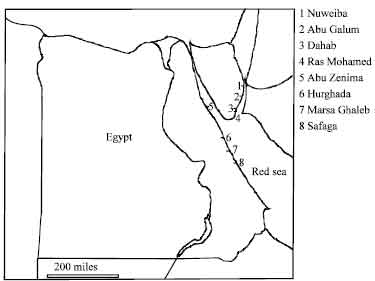 | |
| Fig. 1: | Map of Egypt indicating sample collection sites of Tridacna used in this study |
Amplification of 16S rDNA and Sequencing
An approximately 450 nucleotide portion of mitochondrial large subunit (16S) rRNA gene was amplified from the genomic DNA using the primers 16Sar 5'-CGCCTGTTTATCAAAAACAT-3' and 16Sbr 5'-CCGGTCTGAACTCAGATCACGT-3' (Kessing et al., 1989). Polymerase chain reaction (PCR) amplification was performed using an Eppendorf Master Cycler® gradient and a reaction mixture of 100 μL containing 60-120 ng of genomic DNA, 2 units of Taq polymerase enzyme, 0.5 μM of each primer, 25 mM of each dNTP, 2.5 mM MgCl2 and 1 X Taq buffer (100 mM Tris HCl and 500 mM Kcl). Cycle parameters were 1 min 94°C initial denaturation, 35 cycles of the pattern (30 sec 94°C denaturation, 30 sec 47°C annealing, 1 min 72°C extension) and 10 min 75°C final extension. Double-stranded products were isolated on 2% agarose gels, excised using a QIAquick gel extraction kit (QIAGEN, Germany). A single strand of the amplified partial 16S rDNA gene fragments was directly sequenced with the forward (16Sar) amplification primer using the protocols and reagents for cycle sequencing in the ABI Prism Dye Terminator Cycle sequencing ready reaction kit version 3 (PE Applied Biosystems). Extension products were sequenced on an ABI Prism 3700 automated DNA sequencer (PE Applied Biosystems).
Phylogenetic Analysis
The 16S rDNA sequences were aligned with available Tridacna 16S rDNA sequence data (Schneider and Foighil, 1999) from the GenBank database using PHYDIT 3.1 phylogenetic software (Chun, 1995). The computer-generated sequence alignments were adjusted manually and 387 bp was used in the final analysis. Phylogenetic reconstruction was done using Neighbor Joining (NJ) (Saitou and Nei, 1987), Maximum Parsimony (MP) and Fitch-Margoliash (F) (Fitch and Margoliash, 1967) methods. NJ analysis was performed using the tree construction algorithms available with PHYDIT V. 3.1 (Chun, 1995). Evolutionary distance matrices for the NJ and F methods were generated as described by Jukes and Cantor (1969). Tree topologies were evaluated by 1000 bootstrap re-samplings of the NJ data (Felsenstein, 1985). Phylogenetic trees were drawn with Tree View V. 1.6.6 (Page, 1996) and Canvas™ V. 5 on a Macintosh computer in Center of Marine Biotechnology, University of Maryland Biotechnology Institute, Baltimore, Maryland, USA.
RESULTS AND DISCUSSION
In the present study, the length of the amplified fragment of mitochondrial 16S rDNA gene was approximately 450 bp. The amplified region contained sufficient conserved residues to permit unambiguous alignment to be made among tridacnid species. Of the 387 sites, 115 were variable in our data. The neighbor joining tree in Fig. 2 shows a group supported by a high bootstrap value (97) made up of T. crocea, T. squamosa, T. maxima and tridacnid clams from different locations in the northern Red Sea (Fig. 1). The bootstrap value at the node T. squamosa and sample RM1 was 92. This indicates that they are closely related. The rest of the tridacnid samples are related to T. maxima (bootstrap value of 51). The genetic distance between the samples collected from Safaga, Hurghada, Marsa Ghaleb and Ras Mohamed (RM2) is negligible. Moreover, the genetic distance between samples colleted from Abu Zenima, Nuweiba, Dahab, Abou Galum and Ras Mohamed (RM3) is also insignificant.
Our phylogenetic analyses of 16S rDNA sequences suggest that there are three novel species of Tridacna in the Red Sea. Two of these species are related to T. maxima and one is related to T. squamosa. Of the two species related to T. maxima, one species was found solely in the main basin of the Red Sea (Hurghada, Marsa Ghaleb and Safaga) whereas the other was confined to the Gulfs of Aqaba and Suez (Abu Zenima, Abou Galum, Dahab and Nuweiba). Ras Mohamed, a site at the point where the two gulfs join the main basin, was found to harbor all three novel species. Our results do not support the morphological classification (Mansour, 1946b, c, 1949; Hughes, 1977; Mergner and Mastaller, 1980; Kilada et al., 1998) that suggests the existence of only two tridacnid species, T. maxima and T. squamosa in the northern Red Sea.
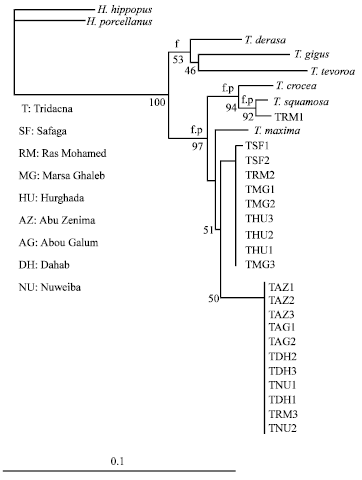 | |
| Fig. 2: | Neighbor-joining phylogenetic tree from analysis of 387 bp of 16S rDNA gene sequences of Tridacna species and Tridacnid clams from the Red Sea. f and p indicate branches which were also found using Fitch-Margoliash and maximum parsimony methods, respectively. The numbers at the nodes are percentages indicating the levels of bootstrap support, based on neighbor-joining analysis of 1,000 re-sampled data sets. The scale bar represents 0.1 substitutions per nucleotide position |
In conclusion, this study suggests that the Red Sea Tridacna population is not exclusive to the two species T. maxima and T. squamosa as was previously believed. Three novel species were discovered. More rapidly evolving sequences will be useful for future additional DNA-based genetic studies of these species.
ACKNOWLEDGMENTS
This study was funded in part by the Egyptian Academy of Scientific Research Program of the National Strategy for Biotechnology and Genetic Engineering (project code: 31). We thank Mr. Osama M. Wahba for his technical assistance. This is contribution No. 03-590 from the Center of Marine Biotechnology.
REFERENCES
- Benzie, J.A. and S.T. Williams, 1998. Phylogenetic relationship among giant clam species (Mollusca: Tridacnidae) determined by protein electrophoresis. Mar. Biol., 132: 123-133.
Direct Link - Felsenstein, J., 1985. Confidence limits on phylogenies: An approach using the bootstrap. Evolution, 39: 783-791.
CrossRefDirect Link - Fitch, W.M. and E. Margoliash, 1967. Construction of phylogenetic trees. Science, 155: 279-284.
PubMedDirect Link - Lucas, J.S., E. Ledua and R.D. Braley, 1991. Tridacna tevoroa Lucas, Ledua and Braley: A recently described species of giant clam (Bivalvia; Tridacnidae) from Fiji and Tonga. Nautilus, 105: 92-103.
Direct Link - Mansour, K., 1946. Communication between the dorsal edge of the mantle and the stomach of Tridacna. Nature, 157: 844-844.
CrossRefDirect Link - Page, R.D.M., 1996. TreeView: An application to display phylogenetic trees on personal computers. Comput. Applied Biosci., 12: 357-358.
CrossRefPubMedDirect Link - Saitou, N. and M. Nei, 1987. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol., 4: 406-425.
CrossRefPubMedDirect Link - Schneider, J.A. and D.O. Foighil, 1999. Phylogeny of giant clams (Cardiidae: Tridacninae) based on partial mitochondrial 16S rDNA gene sequences. Mol. Phylogenet Evol., 13: 59-66.
PubMedDirect Link - Uhlenbruck, G., B.A. Baldo and G. Steinhausen, 1975. Anti-carbohydrate preciptins and haemagluttins in haemolymph from Tridacna maxima (Roding). Z. Immunitatsforsch Exp. Klin. Immunol., 150: 354-363.
PubMed - Yonge, C.M., 1982. Functional morphology and evolution in the Tridacnidae (Mollusca: Bivalvia: Cardiacea). Rec. Aust. Mus., 33: 735-777.
CrossRefDirect Link