
D. Norouzian
Department of Pilot Biotechnology, Pasteur Institute of Iran, Tehran 13164, Iran
A. Akbarzadeh
Department of Pilot Biotechnology, Pasteur Institute of Iran, Tehran 13164, Iran
L. Teimoori
Department of Pilot Biotechnology, Pasteur Institute of Iran, Tehran 13164, Iran
M. Nejati
Department of Bacterial Vaccine and Antigens Production, Pasteur Institute of Iran, Tehran 13164, Iran
A. Faranghi
Department of Pilot Biotechnology, Pasteur Institute of Iran, Tehran 13164, Iran
Asian Journal of Biochemistry
Year: 2006 | Volume: 1 | Issue: 4 | Page No.: 333-337







ABSTRACT
The effects of pH on the activity of tyrosinase (EC 1.14.18.1) leading to the formation of l-3, 4-dihydroxyphenylalanine (l-dopa) by using l-tyrosine as a substrate in the presence of ascorbic acid were studied. The reaction obeyed Eq. (1) and (2). The values of KEB and PKEB were calculated to be -1x10-6 and 6 and those of KESB, PKESB were -2x10-6 and 6.7, respectively. Hydrogen ions inhibited tyrosinase activity competitively.
PDF Abstract XML References Citation
How to cite this article
D. Norouzian, A. Akbarzadeh, L. Teimoori, M. Nejati and A. Faranghi, 2006. Effect of pH on the Synthesis of L-3,4-Dihydroxyphenylalanine by Tyrosinase of Edible Mushroom. Asian Journal of Biochemistry, 1: 333-337.
DOI: 10.3923/ajb.2006.333.337
URL: https://scialert.net/abstract/?doi=ajb.2006.333.337
DOI: 10.3923/ajb.2006.333.337
URL: https://scialert.net/abstract/?doi=ajb.2006.333.337
INTRODUCTION
The enzymatic synthesis of drugs is becoming more popular and is the state of art in biotechnological processes. Most of the biocatalytic or enzymatic processes occur at mild conditions rather than harsh ones. Therefore, it is promising and is also eco-friendly to the environment due to its biodegradability. One of the most important drugs for the treatment of Parkinson's disease is l-3, 4-dihydroxyphenylalanine (l-dopa) (Neeru et al., 2003). Attempts are being made to study the production of l-dopa by fermentation (Enei and Yamada, 1986; Foor et al., 1993). This method could prove to be expensive due to purification steps for the removal of other metabolites produced during microbial production of l-dopa (Gayathri and Bradley, 2002). To synthesize l-dopa via tyrosinase (EC 1.14.18.1) in the presence of ascorbate is another route yielding the desired product and leaving out some unreacted l-tyrosine. In this study the attempts are made to evaluate the effects of various hydrogen ion concentrations on the catalytic activity of mushroom tyrosinase, at different tyrosine concentrations, leading to the formation of l-dopa. The effects of hydrogen ion concentration on the enzyme activity could be considered as activators or inhibitors and therefore, the same kinetic methods and theory can be applied so as to understand the mechanism and nature of the enzyme kinetics might be being obeyed by varying the hydrogen ion concentration on the activity of the enzyme Tyrosinase is a metalo-enzyme which catalyzes other intermediates reaction in the path way leading to the formation of melanin whereas, tyrosine hydroxylase catalyzes only the production of l-dopa. To prevent the subsequent conversion of l-dopa into melanin in the presence of tyrosinase, ascorbate is added to the reaction medium leaving l-dopa as the final product (Rose et al., 1993). Tyrosinases are found in mammalian, plant and microbial cells (Kwang et al., 2000). The enzymatic synthesis/transformation of chemicals could be influenced by reaction conditions such as pH, temperature, substrate, cofactors, shaking and shearing which contribute to the enzymes inactivity and instability. There are reports indicating the effect of pH, temperature on the reaction rates of penicillin acylase, β-galactosidase (Altan 1993; Jurado et al., 2004). In this study attempts are made to study the effect of various pH (4 to 8) on the activity of mushroom tyrosinase in the presence of l-tyrosine and ascorbic acid leading to the synthesis of l-dopa. The kinetic parameters such as Vm, Km were determined by Lineweaver Burk plot and also by plotting Vm/Km against pH, 1/Vm versus 1/H+ and Km/Vm against 1/H+ in order to determine KEB, PKEB, KES B and PKESB.
MATERIALS AND METHODS
Materials
Commercially available edible mushroom was obtained locally. l-tyrosine, l-dopa and ascorbic acid were obtained from Merck. Other reagents used were of analytical grade.
Preparation of Tyrosinase
Tyrosinase was obtained from mushroom according to the acetone precipitation procedure of Bouchillox et al. (1963). The precipitate obtained subsequent to acetone precipitation {twice with acetone and once with 30% (V/V) acetone in double distilled water} was dissolved in minimum amount of phosphate buffer pH 7 and then solid ammonium sulfate was added to give 10% saturation. The mixture was allowed to stand for 2 h at 0°C and then centrifuged at 4000 rpm,4°C for 30 min. To the supernatant ammonium sulfate was further added in order to bring the solution to 60% saturation. After allowing it to stand for 2 h at 4°C and the precipitated protein containing tyrosinase activity was recovered by centrifugation as mentioned above. The precipitate was dissolved in minimum amount of 0.05 M phosphate buffer pH 7 and desalted on the column of Sephadex G-25(2.5H30 cm) equilibrated in the same buffer and eluted at a flow rate of 1 mL minG1 (each fraction 2 mL).The desalted fractions were assayed for protein and enzyme activity. The fractions containing tyrosinase activity were pooled and maintained in 2 mL of cryo-tubes at -20°C.
Assay of Tyrosinase Activity
Tyrosinase activity was determined colorimetrically from the amount of l-dopa produced by tyrosinase action on l-tyrosine in the presence of ascorbic acid and molecular oxygen as reported by Munjal and Sawhney (2002). The reaction mixture contained 1 mL of 2.5 mM tyrosine and ascorbic acid in 0.05 M phosphate buffer pH 7 to which 0.1 mL of appropriately diluted tyrosinase solution was added. The reaction mixture was incubated at 25°C for 25 min. To the above solution, 1 mL of the following reagents were added: 2 M HCl, 2 M NaOH, 15% (w/v) sodium molybdate and finally 15% (w/v) sodium nitrite. Absorbance at λ 460 nm was recorded after 1 h against blank which contained inactivated tyrosinase. One unit of enzyme activity represents the amount of enzyme that produces 1 μmole of l-dopa in one minute under the above assay conditions.
Kinetics Study
A 10 mM solution of l-tyrosine was prepared in different buffers of various pH values {acetate buffer (pH 4.0 to 5.6) and phosphate buffer (pH 6.0 to 8) at the intervals of 0.4 pH units}. The stock substrate solution was appropriately diluted to give desired concentration of l-tyrosine. The formation of l-dopa was determined as above.
RESULTS AND DISCUSSION
The effects of pH on the kinetic of tyrosinase can be written according to the simplified mechanism as discussed by Tippon and Dixon (1979) as follows:
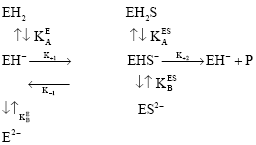 | (1) |
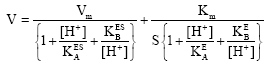 | (2) |
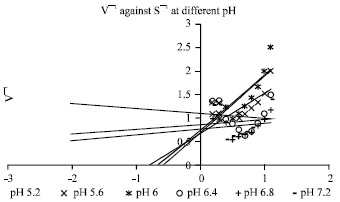 | |
| Fig. 1: | Plot of 1/V against 1/S at different pH values |
| Table 1: | The values of Km and Vm obtained by plotting 1/V vs 1/S at different pH values. The data are mean of n = 6 |
 | |
The values of Km and Vm for tyrosinase acting on l-tyrosine in the presence of ascorbic acid at different pH values are obtained through Lineweaver Burk plots (Table 1). At different pH values straight lines of 1/V vs 1/S plot intersect at points to the left of 1/V axis on the horizontal axis. At the pH values of 6 and below 6 straight lines of 1/V vs 1/S plot intersect at almost common points on vertical axis i.e., 1/V axis while at pH values above 6.4 the lines intersect at different point above horizontal axis (Fig. 1). These findings indicate that the effect of pH on the initial rate of tyrosinase on l-tyrosine is consistent with the mechanism obeying Eq. 1 and 2.
At higher [H+] ion concentration, the inhibition terms [H+]/KEA and [H+]/KESA can be neglected and consequently, because of the inhibition of [H+], a deviation of the lines from the simple intersecting pattern takes place.
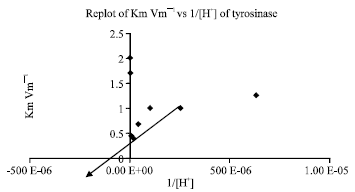 | |
| Fig. 2: | Intercept on horizontal axis yields -(1/KEB) value. Therefore, KEB and pKEB = -log KEB can be calculated. The data are mean of n = 6 |
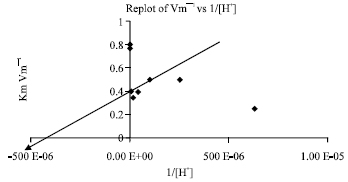 | |
| Fig. 3: | Replot of the vertical axis intercepts of the lines in Fig. 3. Intercept on the horizontal axis yields-(1/KESB). Therefore, KESB and pKESB = -log KESB can be calculated. The data are mean of n = 6 |
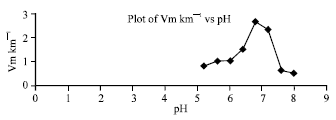 | |
| Fig. 4: | Effect of pH on the maximum velocity of L-dopa synthesis at 25°C using tyrosine as a substrate in the presence of molecular oxygen and ascorbic acid. The data are mean of n = 6 |
When the slopes i.e., Km/Vm of 1/V vs. 1/S at different pH values are plotted against 1/[H+], the ionization constant can be estimated. At higher pH, the values of [H+]/KEA and [H+]/KESA is ignored, only the values of the KEB and KESB can be estimated. The slopes of Km/Vm are re-plotted vs 1/[H+] (Fig. 2). The intercept of the line of Km/Vm vs 1/[H+] plot on the 1/[H+] axis gives -(1/KEB) value. The calculated values of KEB and PKEB are -1H10G6 and 6, respectively. However, the value of KESB is estimated by plotting 1/Vm against 1/[H+] to be-2H10G7 which is close to zero for the effect of pH on tyrosinase acting on l-tyrosine. Therefore, the effect of pH in the synthesis of l-dopa using tyrosinase as an enzyme in the presence of tyrosine, molecular oxygen and a reducing agent like ascorbic acid could be considered to be competitive and the value of PKESB is calculated to be 6.7 (Fig. 3). Therefore, [H+] ion in this reaction is binding to tyrosinase competitively and preventing the enzyme to react with its substrate at lower pH values. This was further confirmed by plotting Vm/Km at low and high pH values against pHs under study.
 | |
| Fig. 5: | Lineweaver burk plot of tyrosinase at different pH values using tyrosine as a substrate. The data are mean of n = 6 |
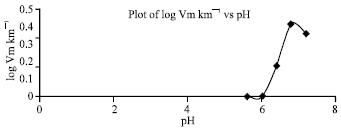
The slopes were determined to be +1 and -1 at acid and alkaline pHs. In such a model Vm results as a function of pH while Km is independent. The slope of the plot of log Vm/Km vs pH is lower than one which is due to the substrate dissociation effect (Fig. 4, 5). In conclusion, [H+] ion concentration acts as a competitive inhibitor of tyrosinase in the conversion of l-tyrosine to l-dopa in the presence of ascorbic acid.
Notation
| E | Enzyme |
| H | Hydrogen |
| [H+] | Hydrogen ion concentration |
| Km | Apparent Michaelis-Menton constant |
| KEA, KEB | Molecular dissociation constants for the free enzyme |
| KESA, KESB | Molecular dissociation constants for the enzyme-substrate complex |
| P | Product |
| S | Substrate |
| v | Enzymatic reaction rate |
| Vm | Apparent maximal enzymatic reaction rate |
REFERENCES
- Altan, E., 1993. The effect of pH on the kinetics of penicillin G acylase obtained from a mutant of E. coli ATCC11105. Process Biochem., 28: 319-324.
Direct Link - Bouchillox, S., P. McMahill and H.S. Mason, 1963. The multiple forms of mushroom tyrosinase: purification and molecular properties of the enzymes. J. Biol. Chem., 238: 1699-1707.
PubMedDirect Link - Jurdao, E., F. Camacho, G. Luzon and J.M. Viceria, 2004. Kinetics models of activity of β-galactosidase: Influence of pH, ionic concentration and temperature. Enzyme Microbiol. Technol., 34: 33-40.
Direct Link - Kong, K.H., M.P. Hong, S.S. Choi, Y.T. Kim and S.H. Cho, 2000. Purification and characterization of a highly stable tyrosinase from Thermomicrobium roseum. Biotechnol. Applied Biochem., 31: 113-118.
Direct Link - Munjal, N. and S.V. Sawhney, 2002. Stability and properties of mushroom tyrosinase and trapped in alginate and polyacrylamide and gelatin gels. Enz. Microb. Technol., 30: 613-619.
Direct Link - Munjal, S.N., S. Kumar and S.K. Sawhney, 2003. A novel method for the immobilization of tyrosinase to enhance stability. Biotechnol. Applied Biochem., 38: 137-141.
PubMedDirect Link - Ros, J.R., J.N. Rodriquez-Lopez and G.F. Canovas, 1993. Effect of L-ascorbic acid on the monophenolase activity of tyrosinase. Biochem. J., 295: 309-312.
Direct Link