
A. Bashtar
Department of Zoology, Faculty of Science, Cairo University, Dokki, Cairo, Egypt
S.A. Ahmed
Department of Medicinal Chemistry, National Research Center,
Dokki, Cairo, Egypt
A.M. Soliman
Department of Medicinal Chemistry, National Research Center,
Dokki, Cairo, Egypt
M.A. Hamed
Department of Medicinal Chemistry, National Research Center,
Dokki, Cairo, Egypt
Asian Journal of Biochemistry
Year: 2006 | Volume: 1 | Issue: 3 | Page No.: 224-235







ABSTRACT
This study has been carried out to investigate the most important aspects of metabolic integration between Schistosoma parasite and its final host under the influence of immunization with S. mansoni antigens. The possible side-effects of soluble worm antigen preparation (SWAP) and soluble egg antigens (SEA) were investigated through estimation of succinate dehydrogenase, lactate dehydrogenase, glucose-6-phosphatase, acid phosphatase and 5`-nucleotidase enzymes activities in hepatocytes of mice. Further, liver function enzymes as aspartate aminotransferase, alanine aminotransferase and alkaline phosphatase were estimated. Cells viability, no of worm burden and ova count were also investigated. The study has revealed that immunization with S. mansoni antigens protect against schistosomiasis by different degree of protection without any side-effects lead to improvement of liver enzymes under study.
PDF Abstract XML References Citation
How to cite this article
A. Bashtar, S.A. Ahmed, A.M. Soliman and M.A. Hamed, 2006. Biochemical Studies on Hepatocytes after Immunization of Mice with Schistosomal Worm and Egg Antigens. Asian Journal of Biochemistry, 1: 224-235.
DOI: 10.3923/ajb.2006.224.235
URL: https://scialert.net/abstract/?doi=ajb.2006.224.235
DOI: 10.3923/ajb.2006.224.235
URL: https://scialert.net/abstract/?doi=ajb.2006.224.235
INTRODUCTION
Schistosomiasis, a chronic and debilitating parasitic disease, affects approximately 200 million people in the developing world and imposes a substantial public health and economic impact (Abo-Madyan et al., 2004; Wang et al., 2004). Although most of the worms migrate against the blood stream to the colon and the bladder, some worms and ova are carried by the blood stream to the liver initiating schistosoma granulomata in variable numbers as well as pathological changes in the liver ranging from early to advanced stage of fibrosis, depending on the degree of ova deposition (Njenga et al.,1998; Silva et al., 2003). Kamo and Ebato (1987) marked that the worm embolism in the intrahepatic portal vein has also a role in hepatic fibrosis. Ascites and oesophagogastric varices could be demonstrated in advanced cases of Schistosoma fibrosis with portal hypertension (Bosshardt et al., 1997; Njenga et al., 1998).
Current control of the disease by chemotherapeutic agents is impractical because of the common occurrence of re-infection after treatment due to the relative resistance of the larval stages of S. mansoni to schistosomicide drugs (Silva et al., 2003). Praziquantel, the currently used drug for chemotherapeutic control, was reported to induce hemorrhages in the lung tissue of the host (Flisser and McLaren, 1989) as well as abdominal pain and diarrhoea (Kabatereine et al., 2003). On the other hand, control through molluscicides is costly and faced with the problem of distribution and accumulation of the applied chemical agents (Nabih et al., 1992). Based upon all these facts, more efforts to induce immunity against schistosomiasis by vaccination have been directed out using either homologous or heterologous antigens (Hillyer, 1995; Capron et al., 1994).
Vaccination of mice with adult worm antigen showed a reduction of the worm load, delayed ovulation, reduction in the numbers of ova in liver and stools and less sever lesions in the liver (Sobh et al., 1996). Moreover, it is positively modulate egg-antigen-induced hepatic granuloma formation which give important implications in the choice of worm-derived antigens for vaccine development (Jacobs et al., 1999).
Immunization of mice with egg antigen showed delayed ovulation and reduction of ova in the liver and stools as well as down-regulation of lymphoproliferative response, but had the worst lesions in the liver with biggest granulomata (Hassanein et al ., 1999). SEA appears to be a good candidate for developing a screening assay for the immunodiagnosis of schistosomiasis (Hillyer et al., 1986).
The aim of the present study was planned to detect the possible side-effects of Shistosoma mansoni worm and egg antigens against infection and to evaluate the protective effect of these vaccines through an estimation of succinate dehydrogenase, lactate dehydrogenase, glucose-6-phosphatase, acid phosphatase, 5` -nucleotidase, alkaline phosphatase, aspartate and alanine aminotransferase enzyme activities as well as worm burden and ova count in mice liver.
MATERIALS AND METHODS
Chemicals
All chemicals used in the present study were of high analytical grade, products of Sigma (USA), Merck (Germany), BDH (England), Riedel de Hàen (Germany) and Fluka (Switzerland).
Animals
The animals used were intact male Swiss albino mice of CDI strain obtained from Theodor Bilharz Research Institute (Cairo) of similar age (8 weeks) and weight (20-25 g). They were maintained on water and stock commercial pellet diet (El-Kahira Company for Oil and Soap) ad libitum.
Experimental Design
The animals were divided into four main groups. The first two main groups consisted of 48 mice and were divided into four equal subdivisions each. The first three subdivisions for each main group were immunized weekly with one dose of either soluble worm antigen preparations (SWAP), or soluble egg antigens (SEA) for two weeks. Mice were sacrificed 10, 20 and 30 days post the last antigen inoculation. The fourth subdivision for each main vaccinated group, received subcutaneous injection with 50 cercariae of Egyptian S. mansoni strain after one week of the last antigen dose. Animals were sacrificed 2 months post schistosomal infection. The third main group acted as infected group and consisted of 6 mice. They received one dose of normal saline weekly for two weeks, then subcutaneously injected with 50 cercariae of Egyptian S. mansoni strain according to Peters and Warren (1969) after one week of the last saline dose. The fourth main group consisted of 24 mice, divided into four subgroups, served as untreated control groups and sacrificed 10, 20, 30 and 60 days post the last saline dose.
Antigen Preparations
Soluble worm antigen preparations (SWAP) was prepared according to the method of Salih et al. (1978), while soluble egg antigens (SEA) was prepared according to the method of Boros and Warren (1970).
Antigen Administration Regimens
The total protein content of each antigen preparation was determined (Bradford, 1976) and the final concentration was adjusted with normal saline to a concentration 50 μg/ml. The immunization schedule was performed according to the method of Nabih and Soliman (1986). Each mouth was sensitized with an intraperitoneal injection of the extracted antigen with a total antigen concentration of 30 μg protein. After one week, a second i.p. injection of the same antigen was taken containing 20 μg protein.
Isolation of Hepatocytes
The liver was perfused with Hank’s solution at rate 3 ml/min, saturated with an oxygen carbon dioxide mixture (95% : 5%) at 37°C. Then, second perfusion takes place containing 5 mg collagenase type IV. The liver was transferred carefully to a beaker, cut and seated in 10 ml Hank’s solution. The free cells were filtered through a gauge to get rid of debris. The cells were centrifuged at 200 g for 2 min to precipitate the intact cells. The supernatant was decanted and the cells were suspended in 5 mL Hank’s solution for further experiments.
Parameter Assays
Enzyme activities were evaluated using end point assay method. Succinate dehydrogenase: The reduction of FAD is coupled with a reduction of tetrazolium salt as INT, the produced formazan of INT is measured colorimetrically at 490 nm (Shelton and Rice, 1957). Lactate dehydrogenase: the reduction of NAD coupled with the reduction of tetrazolium salt with PMS serving as an intermediate election carrier, the resulted formazan of INT was measured colorimetrically at 503 nm. (Babson and Babson, 1973). The three enzymes, G-6-Pase, acid phosphatase and 5`- nucleotidase were measured colorimetrically at 660 nm through measuring the inorganic phosphate release (Swanson, 1955; Wattiaux and De Duve, 1956; Bodansky and Schwartz, 1963, respectively). AST and ALT was estimated through measuring oxaloacetate and pyruvate produced, respectively (Reitman and Frankel, 1957). Alkaline phosphatase was measured as a liberated phenol in the presence of amino-4-antipyrine and sod-arsenate as a blocking agent and potassium ferricyanide as a color reagent. The developed color measured at 510 nm (Kind and King, 1954).
Worm Counting
The degree of protection or the percent of reduction in worm number after challenge was calculated by the method of Tendler et al. (1986).
Ova Count
The number of ova/g tissue was counted by the method of Cheever and Anderson (1971).
Statistical Analysis
Data in the present study are presented as mean±SE. Statistical significance values between control, infected and immunized groups were obtained by the Student t-test (Ronald et al., 1983).
RESULTS
Table 1 shows insignificant changes of cells viability in intact mice vaccinated with SWAP or SEA antigens at different durations. Significant decrease is observed in succinate dehydrogenase, lactate dehydrogenase, glucose-6-phosphatase, alkaline phosphatase, aspartate and alanine aminotransferases enzyme activities in hepatocytes of intact mice after 10 days of immunization with worm or egg antigens, while acid phosphatase and 5`-nucleotidase exhibit significant increase. After 20 days of vaccination SDH, LDH, ALP, G-6-Pase, AST and ALT still recorded significant decrease except G-6-Pase, AST and ALT which recorded insignificant changes after vaccination with SWAP. Contradictory, AP and 5`-nucleotidase showed significant increase except AP which recorded insignificant changes post vaccination with SEA. Thirty days post immunization with SWAP or SEA, all enzymes returned to its normal values.
| Table 1: | Effect of immunization with SWAP and SEA on different biochemical parameters in hepatocyes of intact mice at different durations |
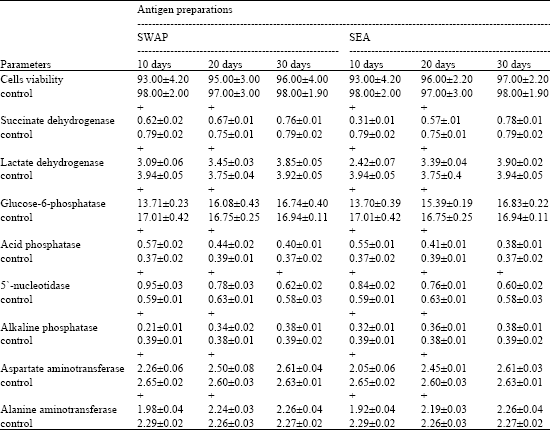 | |
| All values are mean±SE of six mice in each group, Cells viability are expressed as percentages, while enzyme, activities are expressed as μ mol/min/mg protein, SWAP and SEA are soluble worm antigen preparation and soluble egg antigen, respectively, (+) Significant level as compared to each related control group at p<0.05 | |
| Table 2: | Effect of immunization with SWAP and SEA on different biochemical parameters in hepatocyes of infected mice |
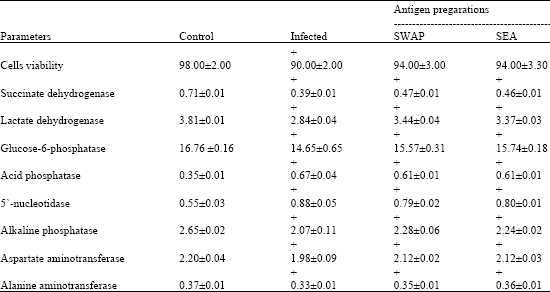 | |
| All values are mean±SE of six mice in each group, Cells viability are expressed as percentages, while enzyme activities are expressed as μ mol/min/mg protein, SWAP and SEA are soluble worm antigen preparation and soluble egg antigen, respectively, (+) Significant level as compared to each related control group at p<0.05 | |
| Table 3: | Number of worm burden and ova count in infected and immunized mice with SWAP and SEA |
 | |
| Worm burden are expressed as means ± SE. of six mice in each group, while ova count are expressed as (means±SE) x103,SWAP and SEA are soluble worm antigen preparation and soluble egg antigen, respectively, (+) Significant level as compared to infected group at p<0.05 | |
Table 2 shows cells viability in infected and immunized mice with SWAP and SEA. Infected mice recorded significant decrease, while showed insignificant changes after immunization exhibiting an enhancement level by 4.12%. Both infected and immunized mice showed significant decrease in all enzyme activities except acid phosphatase and 5`-nucleotidase which recorded significant increase. SDH, LDH, G-6-Pase, AP, 5`-nucleotidase, ALP, AST and ALT recorded amelioration by 11.27, 15.48, 5.48, 17.14, 16.37, 5.41, 8.17 and 6.37%, after vaccination with SWAP, respectively. Immunization with SEA exhibit enhancement by 9.86, 13.91, 6.50, 17.14, 14.55, 8.11, 6.61 and 6.37%, respectively.
Table 3 shows number of worm burden and ova count/ g liver tissue in immunized mice with SWAP or SEA antigens. Significant decrease in number of worm burden in immunized mice with SWAP and SEA amounting 68.18 and 50.00%, respectively. Ova count/g liver tissue recorded also significant decrease by 65.71 and 45.76%, respectively.
DISCUSSION
Concerning the cells viability, the present study revealed an insignificant decrease with all antigens used at different durations as well as upon vaccination of infected mice with different antigen preparations. Contradictory, infected mice showed significant decrease. The obtained results was confirmed by Byram et al. (1979) who stated that hepatocytes showed steatosis or ballooning degeneration with concomitant pyknosis or karyorrhexis by schistosomiasis. Also, the mitochondria appeared swollen with fewer cristae and pale matrices, while an increase in lysosomes and slight proliferation of the Golgi complexes was observed (Jones and Bogitsh, 1979). Moreover, El-Gowhary et al. (1993) mentioned that there was a marked proliferation and dilation in both rough and smooth endoplasmic reticulum with increase of ribosomes. In addition, Saber et al. (1983) and Weiner et al. (1987) remarked a reduction in messenger ribonucleic acid (mRNA). So, schistosomiasis affecting cell organelles, hence cell viability leading to cell death by extensive ova deposition (Boros and Warren, 1970).
As concerning succinate dehydrogenase enzyme activity, the present results revealed a significant decrease in its activity 10 and 20 days post vaccination with all antigen preparations. It returned to its normal control level 30 days post vaccination. This can be explained on the basis that these antigens or its metabolic products acts on gene expression as a signal, so the transcription of DNA specific sequence into messenger RNA is repressed. Gene repression is an effective way of changing enzyme activity (Van Hellemond and Teilens, 1994; Hoek et al., 1997). The present study recorded that both infected and immunized mice showed significant decrease in succinate dehydrogenase enzyme activity as compared to control group. In accordance with this, Ismail et al. (1991) claimed that the activity of succinate dehydrogenase was decreased in the liver homogenate of mice infected with S. mansoni. Daugherty (1955) suggested that this failure was related to schistosoma toxins which accumulate with the mitochondrial fraction and reflected on enzyme activity. Succinate dehydrogenase is the marker enzyme of the mitochondria (Van Noorden and Frederiks, 1992), So any changes in the enzyme activity should be accompanied with changes in the mitochondria (Jones and Bogitsh, 1979; El-Gowhary et al., 1993). This was confirmed by the same authors, who noticed that mitochondria appeared swollen with fewer cristae and pale matrix in the liver cells of mice infected with S. mansoni. An additional evidence of increased mitochondrial membrane permeability was shown by the increased release of mitochondrial enzyme ornithine carbamoyl transferase and isocitrate dehydrogenase into sera of human patients and experimental animals after infection with schistosomes (Rizk et al., 2000). The mitochondrial changes could be originated from the relative anoxia and irritation caused by toxic or metabolic products of adult worm and/or eggs, hence inhibition of mitochondrial oxidation (Ahmed and Gad, 1995), when limiting amount of oxygen are present, both SDH and Krebs cycle enzyme activities are repressed (Van Hellemond and Tielens, 1994).
Concerning lactate dehydrogenase enzyme activity of intact mice vaccinated with different antigens, the present results revealed that there was significant decrease in LDH activity after 10 and 20 days of immunization. The enzyme was returned to its normal control level after 30 days of immunization. Awadalla et al. (1975), Brunk and Cadenas (1988) attributed the decrease in LDH activity to the changes occurred on the permeability of the plasma membrane and leakage of the enzyme from the cytoplasm to blood stream.
The obtained results showed that both infected and immunized mice with different antigens used resulted a significant decrease in enzyme activity as compared to the control group. In agreement with the present results Awadalla et al. (1975) mentioned that the level of lactate dehydrogenase was decreased in the liver tissue homogenate of mice infected with S. mansoni. They attributed the decrease of enzyme activity to the increased cell anoxia and irritation by toxic or metabolic products of the worm. Metwally et al. (1990) conferred that serum LDH was significantly increased in mice infected with S. mansoni. They added that this elevation in serum enzyme activity is due to tissue damage caused by larvae in the infection period.
LDH inhibition revealed the aerobic-anaerobic switch induced by the developing parasite (Tielens et al., 1994). Moreover, lower activity in LDH is in the direction of lactate oxidation direction which could be easily correlated to the crabtree effect of schistosomes (Tielens, 1997) through which lactate is accumulated and glycogen depleted confirming inhibition of aerobic respiration and stimulation of anaerobic glycolysis through hexokinase, a rate limiting enzymes of glycolysis. The decreased enzyme activity due to the increase in anaerobic glycolysis may be attributed also to provision of energy, inhibition of Kreb's cycle, decrease NAD/NADH ratio due to inhibition of mitochondrial oxidation which favors the conversion of pyruvate to lactate, increased metabolic activity of infected liver tissues due to associated inflammation lesions, decreased hepatic blood flow, decreased utilization efficiency of other energy sources such as fat and proteins or from worms themselves which depend on anaerobic glycolysis as the main source of energy production (Ahmed and Gad, 1995).
Concerning the effect of immunization on glucose-6-phosphatase enzyme activity, the present results revealed that there is a significant decrease in enzyme activity 10 days post vaccination. A gradual increase in enzyme activity started from 20-30 days post vaccination, where the enzyme is returned completely to its normal level. Both infected and immunized mice recorded a significant decrease in enzyme activity. This is in agreement with the results of Hara et al. (1981). They attributed the decrease in enzyme activity to molecular and biological changes occurred in hepatic and granulomatous cells. This decrease in enzyme activity may be also due to disturbance occurred in either synthesis and/or degradation of glycogen (Michael et al., 1979; Rizk, 1997) or it may be due to inhibition of translocase (T1), the glucose-6-phosphate transport protein (Scott et al., 1991). This was confirmed by El-Aasar et al. (1989) who described a marked decrease in glycogen content in case of heavy schistosomal infection. Additional histochemical studies by these investigators showed that hepatocytes adjacent to granuloma to be glycogen-deficient and the obstruction of the bile ducts due to developing granuloma may also responsible for this deficiency in glycogen stores in the hepatocytes bordering granulomas. Ahmed and Gad (1995) confirmed this results by noticed a remarkable decrease in the activity of glycogen phosphorylase in S. mansoni infected mice seems to be inconsistent with the increased influx of glucose into the glycolytic pathway. However, this apparent controversy may be compromised by a concomitant decrease of glycogen synthesis as well. This significant depletion of glycogen, two and three months post infection proved the aerobic-anaerobic transition induced by developing schistosomes as hypoxia inducer (Horemans et al., 1992).
Regarding acid phosphatase enzyme activity, the present results showed a highly significant increase in enzyme activity after 10 and 20 days post immunization. The enzyme activity returned to its normal control level 30 days post vaccination with all antigens used. Both infected and immunized mice post infection with different antigen preparations showed also a significant increase in acid phosphatase enzyme activity.
The elevation in acid phosphatase enzyme activity may be due to increased catabolism as a result of the effect of antigens and its metabolites, or increased worm and egg toxins by infection, since all the lysosomal enzymes are activated in condition characterized by increased tissue catabolism (Salah et al., 1976; Rizk, 1997). Frederiks and Marx (1988) attributed the elevation in enzyme activity to aberration of the lysosomes, where acid phosphatase is the lysosomal marker enzyme. This is in accordance with Jones and Bogitsh (1979) who found histochemically an increase in the number of mature lysosomes of the liver cells in mice after S. mansoni infection. These results were confirmed by Rodrigues (1988) who observed an important changes on the lipidic constitution of the lysosomal membrane of S. mansoni infected mice. These changes are provoked by the catabolites excreted by immature or adult worms of S. mansoni present in the portal venous system.
As concerning 5`-nucleotidase enzyme activity, the present results recorded a highly significant increase in its activity 10 and 20 days post immunization. The activity returned to the normal level at 30 days post vaccination with all antigen preparations. Both infected and vaccinated mice post-infection showed also a highly significant increase in the enzyme activity as compared to the control group. This increase in the 5`-nucleotidase activity may be due to active transport occurred through the plasma membrane where the enzyme is located as a result of increased tissue catabolism by the effect of antigens and their metabolites or increased worm and egg toxins by infection which led to enhancement of the phagocytic process and discharged through the plasma membrane (Frederiks and Marx, 1988) and/or due to accelerated nucleic acid metabolism where the catabolism of purine and pyrimidine nucleotides starts with the action of 5`-nucleotidase (Hardonk, 1968). Also, the enzyme activity may be increased due to cellular injury (Michael et al., 1979). These results are in agreement with the finding of Eshchar et al. (1967) who found that 5`-nucleotidase increased significantly in parenchymal hepatic disease and Rizk (1997) who reported an increase of 5`-nucleotidase in murine hepatocytes after S. mansoni infection. These results were confirmed by El-Aasar et al. (1978) who demonstrated histochemically that the sinusoids of infected mice livers showed an increased 5`-nucleotidase enzyme activity.
In the present study, alkaline phosphatase enzyme activity in intact mice administered with S. mansoni antigens showed significant decrease after 10 and 20 days of the last vaccination. The enzyme activity returned to its normal level after 30 days of vaccination with all antigens used. As compared to control group, infected and immunized mice with SWAP and CAP showed a significant decrease in enzyme activity. The decrease in hepatic enzyme activity may be due to the release of the enzyme from the necrotic tissue or due to increased cell membrane permeability as a result of relative anoxia and irritation by toxic or metabolic wastes of the worms (Ahmed, 1994). This is in agreement with Metwally et al. (1990) who noticed that ALP activity was increased in serum of mice infected with S. mansoni. Mansy et al. (1990) attributed the increase in enzyme activity to the proliferation of bile ductules and bile canaliculi as a result of schistosomiasis.
It is concerned to study transaminases enzyme activities which showed a highly significant decrease in their activities 10 days post the last vaccination with all antigens used. A gradual recovery to the normal level started from 20 days post vaccination till they reached the exact control value after 30 days of immunization. Also in both, infected and immunized mice AST and ALT activities significantly decreased as compared to the control group. El-Aasar et al. (1989) attributed the decrease of transaminase enzyme activities in mice livers to the decrease in hepatic cell population due to liver fibrosis or due to the release of the enzyme from the damaged livers into the circulation as a result of increased cell membrane permeability.
As seen by Waldorf and Harper (1963), AST activity was decreased relative to liver protein, while ALT was increased. This is in agreement with Awadalla et al. (1975) and Varley et al. (1980) who stated that AST and ALT have a control role in amino acid metabolism. Malabsorption through intestine as a result of extensive ova deposition in the intestine of S. mansoni infection may be a contributing factor in the decrease of protein synthesis through defect in the absorption of amino acids which are considered as the building blocks of any protein molecule. Thus, a defect in protein metabolism may lead to defects in enzyme synthesis and hence, derangement of many anabolic pathways.
As concerning to worm burden and egg count, the present results recorded significant reductions in both worm and egg count upon vaccination with SWAP and CAP. In agreement with the present results, Tendler et al. (1991) found that a soluble extract of adult worm generates 75% protection in mice after challenge with S. mansoni cercariae even in the absence of adjuvant. Smithers and Terry (1967) proposed that the main stimulus for immunity in schistosomiasis is the adult worm. This indicated the existence, within adult worm antigens, of several molecules that might play a role in activation and modulation of granulomatous hypersensitivity to eggs (Hirsch and Goes, 1996).
Hassanein et al. (1997) and Kamel et al. (1998) mentioned that vaccination of mice with egg antigen caused a delayed ovulation and reduction of ova in liver and stool, hence decreased the granuloma diameter and amelioration of histopathological changes in liver. They added that immunization with SEA induced granulomatous hyporesponsiveness and down regulate lymphoproliferative response to SEA as compared to infected group, while antischistosomal IgG level was increased.
As previously mentioned above the decreased number of worms and eggs after immunization lead to decrease of Schistosoma toxins that accumulated with the mitochondrial fraction or any subcellular organelles and affected the enzyme activities. Hence, upon vaccination the enzymes activities were improved lead to enhancement in different metabolic pathways and amelioration in cells viability.
In conclusion, soluble worm antigen preparation (SWAP) have more protective effect against schistosomiasis without any side effects followed by soluble egg antigen (SEA).
REFERENCES
- Abo-Madyan, A.A.,T.A. Morsy and S.M. Motawea, 2004. Efficacy of myrrh in the treatment of schistosomiasis haematobium and mansoni in ezbet el-bakly tamyia center el-fayoum governorate egypt. J. Egypt. Soc. Parasitol., 34: 423-446.
Direct Link - Ahmed, S.A. and M.Z. Gad, 1995. Effect of schistosomal infection and its treatment on some key enzymes of glucose metabolism in mice livers. Arzneimittel Forschung, 45: 1324-1328.
Direct Link - Bodansky, O. and M.K. Schwartz, 1963. Comparative effects of L-histidine on the activities of 5`-nucleotidase and alkaline phosphatase. J. Biol. Chem., 238: 3420-3427.
Direct Link - Awadalla, H.N., A.F. Sherif, A.Z. Shafei, A.H. Khalil and F.K. Guirgis, 1975. Enzyme levels in homogenates of liver from mice infected with Schistosoma mansoni and from uninfected mice. Int. J. Parasitol., 5: 27-31.
PubMed - Bodansky, O. and M.K. Schwartz, 1963. Comparative effects of L-histidine on the activities of 5`-nucleotidase and alkaline phosphatase. J. Biol. Chem., 238: 3420-3427.
Direct Link - Boros, D.L. and K.S. Warren, 1970. Delayed hypersensitivity granuloma formation and dermal reaction induced and elicited by a soluble factor isolated from Schistosoma mansoni eggs. J. Exp. Med., 132: 488-507.
Direct Link - Bosshardt, S.C., G.L. Freeman, W.E. Secor and D.G. Colley, 1997. IL-10 deficit correlates with chronic, hypersplenomegaly syndrome in male CBA/J mice infected with Schistosoma mansoni. Parasite Immunol., 19: 347-353.
PubMed - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Brunk, U. and E. Cadenas, 1988. The potential intermediate role of lysosomes in oxygen free radical pathology. APMIS, 96: 3-13.
CrossRefDirect Link - Byram, J.E., M.J. Doenhoff, R. Musallam, L.H. Brink and F. Von Lichtenberg, 1979. Schistosoma mansoni infections in T-cell deprived mice and the ameliorating effect of administring homologous chronic infection serum. II. Pathology. Am. J. Trop. Med. Hyg., 28: 274-285.
Direct Link - Capron, A., G. Riveau, J.M. Grzych, D. Boulanger, M. Capron and R. Pierce, 1994. Development of a vaccine strategy against human and bovine schistosomiasis background and update. Trop. Geogr. Med., 46: 242-246.
PubMed - Cheever, A.W. and L.A. Anderson, 1971. Rate of destruction of Schistosoma mansoni eggs in tissues of mice. Am. J. Trop. Med. Hyg., 20: 62-68.
Direct Link - Daugherty, J.W., 1955. The effect of schistosoma mansoni infections on liver function in mice. ii. further studies on intermediary metabolism. Am. J. Trop. Med. Hyg., 4: 1072-1079.
Direct Link - El-Aasar, A.A., S.M. Hasanein, M.N. El-Bolkainy, I. El-Sebai and M.M. El-Merzabani, 1978. Bladder carcinogenesis in bilharzia infested swiss albino mice. Europ. J. Cancer, 14: 645-648.
PubMed - El-Gowhary, S.H., A.E. Rahmy, M.Z. El-Azzouni, A.I. Nagil and A. El-Medany, 1993. . Oral contraceptive pills in experimental Schistosomiasis mansoni Parasitological biochemical histopathological and ultrastructural studies. J. Egypt Soc. Parasitol., 23: 609-619.
PubMed - Eshchar, J., C. Rudzki and H.J. Zimmerman, 1967. Serum levels of 5'-nucleotidase in disease. Am. J. Clin. Pathol., 47: 598-606.
CrossRefDirect Link - Flisser, A. and D.J. McLaren, 1989. Effect of praziquantel treatment on lung-stage larvae of Schistosoma mansoni in vivo. Parasitology, 98: 203-211.
CrossRefDirect Link - Frederiks, W.M. and F. Marx, 1988. A quantitative histochemical study of 5`-nucleotidase activity in rat liver using the lead salt method and polyvinyl alcohol. Histochem. J., 20: 207-214.
CrossRef - Hara, A., K. Fukuyama and W.L. Epstein, 1981. Angiotensin-converting enzyme and other enzymes in livers of mice with experimental schistosomiasis. Exp. Mol. Pathol., 35: 199-210.
CrossRefDirect Link - Hassanein, H., M. Akl, Z. Shaker, H. El-Baz, R. Sharmy, I. Rabiae and S. Botros, 1997. Induction of hepatic egg granuloma hyporesponsiveness in murine schistosomiasis mansoni by intravenous injection of small doses of soluble egg antigen. APMIS, 105: 777-783.
CrossRef - Hassanein, H., M. Kamel, A. Badawy, N. El-Ghorab and H. Abdeen et al., 1999. Anti‐miracidial effect of recombinant glutathione S‐transferase 26 and soluble egg antigen on immune responses in murine schistosomiasis mansoni. APMIS, 107: 723-736.
CrossRefDirect Link - Hillyer, G.V., L.L. Nieves-Frau and G. Vazquez, 1986. Identification of a genus-specific Schistosoma mansoni soluble egg antigen reactive with the serum of infected patients. Am. J. Trop. Med. Hyg., 35: 1198-1204.
Direct Link - Hillyer, G.V., 1995. Comparison of purified 12 kDa and recombinant 15 kDa Fasciola hepatica antigens related to a schistosoma mansoni fatty acid binding protein. Memorias do Instituto Oswaldo Cruz, 90: 249-253.
CrossRefDirect Link - Hirsch, C. and A.M. Goes, 1996. Characterization of fractionated Schistosoma mansoni soluble adult worm antigens that elicit human cell proliferation and granuloma formation in vitro. Parasitology, 12: 529-535.
PubMed - Hoek, R.M., R.E. Van Kestern, A.B. Smit, M. de Jong-Brink and W.P. Geraerts, 1997. Altered gene expression in the brain caused by a trematode parasite neuropeptide genes are preferentially affected during parasitosis. Proc. Natl. Acad. Sci., 94: 14072-14072.
Direct Link - Horemans, A.M.C., A.G.M. Tielens and S.G. Van den Berg, 1992. The reversible effect of glucose on the energy metabolism of Schistosoma mansoni cercariae and schistosomula. Mol. Biochem. Parasitol., 51: 73-79.
CrossRefDirect Link - Jacobs, W., A. Deelder and E. Marck, 1999. Schistosomal granuloma modulation. II. Specific immunogenic carbohydrates can modulate schistosomae-egg-antigen-induced hepatic granuloma formation. Parasitol. Res., 85: 14-18.
CrossRef - Jones, L.G. and B.J. Bogitsh, 1979. Further observations on ultrastructural changes in hepatocytes of mice infected with Schistosoma mansoni. Z. Parasitenkd, 60: 185-192.
CrossRef - Kabatereine, N.B., J. Kemijumbi, J.H. Ouma, R.F. Sturrock and A.E. Butterworth et al., 2003. Efficacy and side effects of praziquantel treatment in a highly endemic Schistosoma mansoni focus at lake Albert Uganda. Trans. R. Soc. Trop. Med. Hyg., 97: 599-603.
Direct Link - Kamel, M., T.S. Abou-Shousha, D. Maghawry, R. Charmy and H.I. Hassanein, 1998. The immune responses associated with S. mansoni SEA-induced granulomatus hyporesponsiveness in two different strains of mice. International Conference on Schistosomiasis, March, 15-19, Organized by the Schistosomiasis Research Project, Ministry of Health and Population, Cairo, Egypt.
- Kamo, E. and T. Ebato, 1987. Immunopathologic studies on liver cirrhosis in albino rats infected with S. japonicum in comparison with other laboratory animals. Jap. J. Parasitol., 36: 68-79.
Direct Link - Kind, P.R.N. and E.J. King, 1954. Estimation of plasma phosphatase by determination of hydrolysed phenol with amino-antipyrine. J. Clin. Pathol., 7: 322-326.
CrossRefPubMedDirect Link - Metwally, A.A., I. Janku, F. Kemper, M.T. Khayyal, F.A. Fbeid and S.S. Botros, 1990. Effect of schistosomiasis infection on the clearance of phenazone in mice. Arzneim. Forsch., 40: 206-209.
PubMedDirect Link - Nabih, I. and A.M. Soliman, 1986. Studies on freshwater snails, specific intermediate hosts for schistosomiasis-II. Isolation of total proteins from native and irradiated snails. Cell. Mol. Biol., 32: 315-317.
PubMed - Nabih, I., M. Rizk and A.M. Soliman, 1992. Studies on carbohydrates extracted from native and chemically treated Biomphalaria alexandrina snails. Comp. Biochem. Physiol., 102: 397-402.
Direct Link - Njenga, M.N., I.O. Farah, G.K. Muchemi and M. Nyindo, 1998. Peri-portal fibrosis of the liver due to natural or experimental infection with Schistosoma mansoni occurs in the Kenyan baboon. Ann. Trop. Med. Parasitol., 92: 187-193.
CrossRef - Peters, P.A. and K.S. Warren, 1969. A rapid method of infecting mice and other laboratory animals with Schistosoma mansoni: Subcutaneous injection. J. Parasitol., 55: 558-560.
CrossRefDirect Link - Reitman, S. and S. Frankel, 1957. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol., 28: 56-63.
CrossRefPubMedDirect Link - Rodrigues, L.E., 1988. Biochemistry of schistosomiasis mansoni. vii. lipid changes of lysosomal membranes during the initial phase of liver injury. Mem. Inst. Oswaldo. Cruz., 83: 47-52.
PubMed - Salih, S.Y., A. Bartlett and A. Voller, 1978. Detection of antibodies by enzyme immunoassay in human schistosoma mansoni infection a clinical chemotherapeutic study. Trop. Parasitol., 29: 409-412.
Direct Link - Scott, H.M., M.W. Coughtrie and A. Burchell, 1991. Steroid sulfates inhibit rat hepatic microsomal glucose-6-phosphatase system. Biochem. Pharmacol., 41: 1529-1532.
Direct Link - Rice, M.E. and E. Shelton, 1957. Comparison of the reduction of two tetrazolium salts with succinoxidase activity of tissue homogenates. J. Nat. Cancer Inst., 18: 117-125.
PubMedDirect Link - Smithers, S.R. and R.J. Terry, 1967. Resistance to experimental infection with Schistosoma mansoni in rhesus monkeys induced by the transfer of adult worm. Trans. R. Soc. Trop. Med. Hyg., 61: 517-533.
CrossRefDirect Link - Sobh, M., F. Moustafa, S. Hamid and M. Ghoneim, 1996. Schistosomal-specific nephropathy in Syrian golden hamsters treatment by induction of antigen excess. Nephrol. Dial. Transplant., 11: 2178-2184.
Direct Link - Tendler, M., R.M. Pinto, L.A. Oliveira, G. Gebara and N. Katz, 1986. Schistosoma mansoni: Vaccination with adult worm antigens. Int. J. Parasitol., 16: 347-352.
CrossRefDirect Link - Tendler, M., R.M. Pinto, A. Lima, O. de, W. Savino and N. Katz, 1991. Vaccination in murine schistosomiasis with adult worm-derived antigens variables influencing protection in outbred mice. Intl. J. Parasitol., 21: 299-306.
Direct Link - Tielens, A.G., 1997. Properties and function in metabolism of schistosomal hexokinase. Biochem. Soc. Trans., 25: 127-130.
Direct Link - Van Hellemond, J.J. and A.G. Tielens, 1994. Expression and functional properties of fumarate reductase. Biochem. J., 304: 321-331.
Direct Link - Wang, Y., E. Holmes, J.K. Nicholson, O. Cloarec and J. Chollet et al., 2004. Metabonomic investigations in mice infected with schistosoma mansoni an approach for biomarker identification. Proc. Natl. Acad. Sci., 101: 12676-12681.
CrossRef - Wattiaux, R. and C. de Duve, 1956. Tissue fractionation studies. 7. Release of bound hydrolases by means of Triton X-100. Biochem. J., 63: 606-608.
PubMedDirect Link - Weiner, F.R., M.J. Czaja, M.A. Giambrone, S. Takahashi, L. Biempica and M.A. Zern, 1987. Transcriptional and post transcriptional effects of dexamethasone on albumin and procollagen. Biochemistry, 26: 1557-1562.
PubMed