
Ramzy A. Yousif
Department of Fisheries and Wildlife Science, College of Animal Production Science and Technology,
Sudan University of Science and Technology, Khartoum, Sudan
LiveDNA: 249.31450
Seemab Zehra
Beacon Development Company, King Abdullah University of Science and Technology, Thuwal, Jeddah, Saudi Arabia
Fouzi A. Mohamed
Department of Fisheries and Wildlife Science, College of Animal Production Science and Technology,
Sudan University of Science and Technology, Khartoum, Sudan
Asian Journal of Animal and Veterinary Advances
Year: 2022 | Volume: 17 | Issue: 1 | Page No.: 16-27







ABSTRACT
Background and Objective: Reducing fishmeal levels is key to reducing feed costs for commercial fish farming and ensuring the sustainability of this enterprise. This study was aimed to evaluate the effects of replacing fish meal protein with locust meal protein in the feeds for fingerling Oreochromis niloticus (3.5±0.94 g). Materials and Methods: Seven practical diets (350 g kg–1 crude protein, 14.90 kJ g–1 gross energy) replacing 0, 10, 20, 30,40, 50 and 60% fish meal protein by locust meal protein were prepared. The diets were fed to triplicate groups of fish near to satiation for 8 weeks. Results: The live weight gain (LWG, 262.6-719.7%), feed conversion ratio (1.58-3.18), protein efficiency ratio (0.70-1.41), specific growth rate (2.30-3.76% day–1), protein retention efficiency (3.62-9.42), hemoglobin (61.2-98.1 g dL–1), hematocrit (16.37-31.35%) and RBCs counts (13.5-23.2×106 mm–3) in fish fed diets with 0, 10,20 and 30% replacement of fish meal with locust meal did not show any significant differences. Conclusion: However, further replacement of fish meal by locust meal beyond 30% resulted in a significant fall (p<0.05) in the above parameters indicating that fish meal could be replaced by locust meal up to 30%. Based on the broken-line analysis of LWG, the optimum replacement level was found to be 29.8% which would be useful in formulating cost-effective commercial feeds for the intensive culture of this fish.
PDF Abstract XML References Citation
How to cite this article
Ramzy A. Yousif, Seemab Zehra and Fouzi A. Mohamed, 2022. Use of Locust Meal as Alternative Protein Source to Fish Meal in Practical Diets for Fingerling Oreochromis niloticus. Asian Journal of Animal and Veterinary Advances, 17: 16-27.
DOI: 10.3923/ajava.2022.16.27
URL: https://scialert.net/abstract/?doi=ajava.2022.16.27
DOI: 10.3923/ajava.2022.16.27
URL: https://scialert.net/abstract/?doi=ajava.2022.16.27
INTRODUCTION
Due to the ever-increasing demand for high-quality protein food, a need has been felt to meet this increasing demand through aquaculture as the wild stock has been exhausted due to overexploitation of the natural resources. Aquaculture is considered to be the answer to the current shortage of commercial fishery production and its intensification has led to the reliability of artificial feeding1-3. Protein is the most expensive item of feeds and is perhaps the crucial factor for the growth of cultured species4-7. Fish meal is the most alluring protein source for aquaculture diets as it is having a higher amount of protein, balanced amino acid and fatty acid profiles, more digestibility and is more palatable. Despite this, the more price of fish meals and low accessibility make it difficult to be used in most of the aquafeeds8. Therefore, the fish diet needs to be replaced with other conservation measures to promote sustainable aquaculture. Edible insects are important traditional food items in various parts of the world9-12. Orthoptera, especially locusts are often bred to feed on pets and zoo animals and have been inspected for animal feed13. The presence of large numbers of locusts that have died as a result of locust infestations makes them the ideal food for livestock, especially chickens. The expansion of aquaculture in Africa and Asia as well as the exploration for other protein sources, has resulted in the feeding of locusts and grasshopper in different trials and catfish and tilapia14. Locusts, an excellent source of protein, some species differ in their protein content i.e., from 50-60% of dry weight. It also contains sufficient amounts of iodine, phosphorus, iron, thiamine, riboflavin, niacin and carbohydrate levels are very low in locusts15-17. Several investigators are paying attention to replacing fish meals with locust meal18-22.
Tilapia has become an eminent fish species in the world and plays a growing role in the global aquaculture trade. Therefore, the development of less expensive feeds using less expensive plant and animal protein sources will contribute to future sustainable aquaculture development23-29. Tilapia is a freshwater fish belonging to the Cichlidae family. It is currently recognized as the aquatic chicken owing to its rapid growth, adaptability to disease, high meat quality, capability to grow and reproduce in captivity and feed at low trophic levels24,30,31. Tilapia is currently cultivated in more than 100 countries worldwide30,32-34. Tilapia is farmed much faster and on a larger scale than any other group of fish. Rapid advances in livestock and a wide range of uses could mean that tilapia may eventually overtake carps to become the most important domestic fish. Tilapia has already become one of the most important farmed fish and is increasingly involved in international maritime trade35,36. There are about ten species of tilapia used to make aquaculture. Nile tilapia is the most important species of tilapia in the world. As the feed is the most expensive, it usually ranges from 30-60% of the total alterable price, affected by the strength of the cultural process37-39. The development of affordable feeds using cheap and locally available plant protein sources plays a major role in its future sustainable marine development. To this end, research was accomplished to replace the fishmeal protein with locust meal protein for Nile tilapia so that cost-effective nutritionally-balanced feeds could be made for its intensive culture.
MATERIALS AND METHODS
Study area: This study was conducted during the period of (12th January to 9th March, 2021), at the hatchery of the Department of Fisheries and Wildlife Science, Sudan University of Science and Technology, Khartoum, Sudan.
Preparation of experimental diets: Feasibility of replacing fishmeal with locust meal for Nile tilapia O. niloticus fingerling by were found out by preparing seven diets replacing 0% (D1), 10% (D2), 20% (D3), 30% (D4), 40% (D5), 50% (D6) and 60% (D7) fish meal protein by locust meal protein procured from (6 Market), Haj Yousif, Khartoum, Sudan in Table 1. All diets contained 350 g kg–1 crude protein40 and 14.90 kJ g–1gross energy. All the ingredients were weighed and blended thoroughly. These were then steam cooked at 80°C in hot water and oil, mineral and vitamin premixes were included in the bowl in isolation with continuous blending at 60°C. The final diet was cut as the small cubes and stored at -20°C for further utilization. The proximate compositions of the experimental diets used in this experiment were analyzed and are mentioned in Table 1. The amino acid content of experimental diets was analysed using automatic amino acid analyzer biochrom (UK). Recovery hydrolysis for analysis of tryptophan was performed in 4 N methanesulfonic acid and for sulphur amino acids in performic acid in Table 2. The fatty acid profiles of the experimental diets were analyzed gas-liquid chromatography (Biobase Bk-Gc112A High-Performance Gas Chromatograph, Biobase Bioindustry (Shandong) Co., Ltd., Shandong, China).
Experimental system and animals: Fingerling O. niloticus were got from Hussien Fadoul Fish Farm, Soba-Khartoum, Sudan. These were transported to the hatchery of the Department of Fisheries and Wildlife Science, Sudan University of Science and Technology, Khartoum, Sudan and given a prophylactic dip in KMnO4 solution (1:3000) and stocked and kept in oxygenated plastic containers (aquarium) in fisheries laboratory.
| Table 1: | Ingredients composition of experimental diets | ||||||
Experimental diets | |||||||
| Ingredients (g kg–1 dry diet) | D1 | D2 | D3 | D4 | D5 | D6 | D7 |
| Fish meal† | 147.1 | 132.4 | 117.6 | 102.9 | 88.2 | 73.5 | 58.8 |
| Locust meal‡ | 0 | 19.1 | 38.2 | 57.3 | 76.4 | 95.4 | 114.5 |
| Groundnut cake§ | 57.7 | 57.7 | 57.7 | 57.7 | 57.7 | 57.7 | 57.7 |
| Mustard oil cake¶ | 162.2 | 162.2 | 162.2 | 162.2 | 162.2 | 162.2 | 162.2 |
| Soybean meal†† | 311.1 | 311.1 | 311.1 | 311.1 | 311.1 | 311.1 | 311.1 |
| Wheat middlingठ| 142.9 | 142.9 | 142.9 | 142.9 | 142.9 | 142.9 | 142.9 |
| Cod liver oil | 50 | 46 | 42 | 38 | 34 | 30.1 | 26.1 |
| Mineral premix§§ | 20 | 20 | 20 | 20 | 20 | 20 | 20 |
| Vitamin premix§¶ | 10 | 10 | 10 | 10 | 10 | 10 | 10 |
| Alpha cellulose | 99.1 | 98.7 | 98.4 | 97.9 | 97.5 | 97.1 | 96.7 |
| Total | 1000 | 1000 | 1000 | 1000 | 1000 | 1000 | 1000 |
| Proximate analysis (g kg–1 dry diet) | |||||||
| Protein CP | 350±1.3 | 350±2.0 | 350±1.7 | 350±6.0 | 350±5.1 | 350±2.7 | 350±4.5 |
| Ether extract | 90.1±4.0 | 90.2±6.0 | 90.4±8.0 | 90.5±2.0 | 90.6±4.0 | 90.8±2.0 | 90.9±1.0 |
| Fiber content | 58.9±3.0 | 60.5±1.0 | 62.1±2.0 | 63.7±5.0 | 65.4±3.0 | 67.0±3.0 | 68.6±2.0 |
| Ash | 55.9±1.0 | 55.7±1.0 | 55.5±3.0 | 55.2±6.0 | 55.0±4.0 | 54.7±2.0 | 54.5±4.0 |
| Calculated GE (kJ g–1, dry diet) | 14.9±0.3 | 14.9±0.2 | 14.9±2.0 | 14.9±0.1 | 14.9±0.4 | 14.9±0.3 | 14.9±0.2 |
†Fishmeal 680 g kg–1 CP, ‡Locust meal 523.9 g kg–1 CP, §Groundnut cake 520 g kg–1 CP, ¶Mustard oil cake 370 g kg–1 CP, ††Soybean meal 450 g kg–1 CP, ‡§Wheat Middling 140 g kg–1 CP, §§Mineral mixture (g kg–1 dry diet) calcium biphosphate 135.7, calcium lactate 326.9, ferric citrate 29.7, magnesium sulphate 132.0, potassium phosphate (dibasic) 239.8, sodium biphosphate 87.2, sodium chloride 43.5, aluminium chloride.6H2O 0.154, potassium iodide 0.15, cuprous chloride 0.10, manganous sulphate H2O 0.80, cobalt chloride 6H2O 1.0, zinc sulphate 7H2O 4.0 (Halver, 2002)96, §¶Vitamin mixture (g kg–1dry diet) choline chloride 5.0, inositol 2.0, ascorbic acid 1.0, niacin 0.75, calcium pantothenate 0.5, riboflavin 0.2, menadione 0.04, pyridoxine hydrochloride 0.05, thiamine hydrochloride 0.05, folic acid 0.015, biotin 0.005, alpha-tocopherol 0.4, vitamin B12 0.0001, Amipharma Laboratories Ltd, Khartoum North, Sudan (Halver, 2002)96 | |||||||
| Table 2: | Amino acid composition (g kg–1 dry matter) of the experimental diets | ||||||
Experimental diets | |||||||
D1 | D2 | D3 | D4 | D5 | D6 | D7 | |
| Arginine | 27.1±0.60 | 27.2±0.1 | 27.3±0.0 | 27.3±0.03 | 27.4±0.01 | 27.4±0.05 | 27.5±0.02 |
| Histidine | 9.9±0.1 | 9.9±0.4 | 9.9±0.1 | 9.9±0.2 | 9.9±0.3 | 9.8±0.3 | 9.9±0.1 |
| Isoleucine | 14.1±0.2 | 14.2±0.5 | 14.3±0.2 | 14.5±0.4 | 14.6±0.2 | 14.8±0.1 | 14.9±0.01 |
| Leucine | 24.6±0.5 | 24.6±0.3 | 24.6±0.1 | 24.6±0.0 | 24.6±0.2 | 24.6±0.1 | 24.7±0.01 |
| Lysine | 22.3±0.1 | 21.3±0.1 | 21.5±0.4 | 21.0±0.2 | 20.6±0.2 | 20.1±0.1 | 19.7±0.05 |
| Methionine | 6.9±0.2 | 6.9±0.3 | 6.9±0.5 | 6.7±0.1 | 6.9±0.5 | 6.9±0.2 | 6.8±0.03 |
| Cystine | 2.7±0.1 | 2.8±0.1 | 2.9±0.1 | 3.0±0.1 | 3.1±0.1 | 3.2±0.3 | 3.3±0.05 |
| Phenylalanine | 14.8±0.1 | 14.7±0.1 | 14.5±0.3 | 14.3±0.3 | 14.1±0.5 | 13.9±0.1 | 13.7±0.03 |
| Tyrosine | 14.0±0.2 | 14.0±0.4 | 14.1±0.1 | 14.1±0.1 | 14.2±0.2 | 14.2±0.3 | 14.3±0.03 |
| Threonine | 13.3±0.1 | 13.3±0.5 | 13.2±0.3 | 13.1±0.2 | 13.1±0.2 | 13.0±0.1 | 12.9±0.04 |
| Tryptophan | 2.4±0.1 | 2.5±0.3 | 2.7±0.1 | 2.8±0.1 | 3.0±0.2 | 3.2±0.1 | 3.3±0.03 |
| Valine | 17.5±0.5 | 17.5±0.2 | 17.5±0.2 | 17.5±0.1 | 17.4±0.4 | 17.4±0.3 | 17.4±0.03 |
| 1Mean values of 3 replicate ±SEM and 2Not statistically significant (p>0.05) | |||||||
At this time, the fish were fed to apparent satiation by feeding a diet consisting of mustard oil cake, soybean meal and wheat middling in the form of soft cake. For conducting the experiments, Oreochromis niloticus fingerlings (3.5±0.94 g) were separated from the above-acclimated lot and kept in triplicate groups in 80 L circular polyvinyl tanks (water volume 65 L) fitted with a continuous water flow-through (1-1.5 L min–1) system at the rate of 20 fish per tank for each dietary treatment. Fish were supplied with experimental diets as soft cake to apparent satiation twice daily at 0900 and 1600 hrs. No feed was offered to the fish on the day they were weighed. Initial and weekly weights were recorded on a top-loading balance. The feeding trial lasted for 8 weeks. Unconsumed feed was strained on a screen, dried and weighed to assess the amount of feed eaten.
Water quality parameters: Water temperature, dissolved oxygen, free carbon dioxide, pH and total alkalinity during the feeding trial were recorded following Yousif et al.41. The average water temperature, dissolved oxygen, free carbon dioxide, pH and total alkalinity over the 4 weeks feeding trial, based on daily measurements, were 25.0-28.5°C, 6.5-7.4 mg L–1, 7-8.4 mg L–1, 7.2-7.7 and 75-83.5 mg L–1, respectively.
Proximate composition analyses: After starvation of 24 hrs, a randomized sample of 25 fish were killed by dipping in MS-222 solution (200 mg L–1), minced and six subsamples of initial body composition analysed. At the end of the trial, nine fish from each replicate of dietary treatments were sampled, killed, pooled separately and frozen at -20°C for analyzing the biochemical composition. Six subsamples of the pooled samples of each replicate (n = 3×6) were subjected to proximate composition analysis. Proximate composition of the diets, initial and final carcass was worked out following Yousif et al.42 as described in the previous investigation43. Biobase high automatic oxygen bomb calorimeter (Biobase Meihua Trading Co., Ltd. China) was used for estimating the gross energy content of experimental diets.
Blood collection and analysis: Blood from the caudal vein of 9 fish per replicate was randomly collected, pooled for analyzing haematological parameters. The procured blood was transferred to heparinized Eppendorf tubes. The three subsamples (n = 3×3) of pooled samples from the heparinized tube were utilized for the assessment of the RBCs and Hb following the methods of Vani et al.44. The Hct level was examined by centrifuging fresh blood at 3600 g for 6 min in a micro Hematocrit Centrifuge (SH-120 High-Speed Micro Hematocrit, China).
Estimation of liver superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPx) activities: Liver was taken off from five starved fish of each replicate tank, pooled and five subsamples (n = 3×5) were utilized for assessing the SOD, CAT and GPx activities following the procedures of Zehra and Khan45, Khan and Khan46 and Abdel-Hameid et al.47, respectively. The procedure used by the researcher48 was utilized to evaluate the tissue protein content for the calculation of enzyme-specific activities.
Diet performance evaluation: The effects of replacement of fish meal by locust meal in diets for fingerling O. niloticus during the present experiment was assessed by considering the growth and conversion efficiency indices49 as follows:
 |
 |
Statistical analysis: The growth data were subjected to one-way analysis of variance50 before analysis, data were tested for normality using Shapiro-ilk test and the homogeneity of variance using Levene’s test for equality of variances. When a significant treatment effect was observed, Tukey’s honestly significant difference test was used for multiple mean comparisons at a p<0.05 level of significance. Quadratic regression analyses of live weight gain were done to evaluate the optimum replacement level of fish meal by locust meal51. Statistical analyses were done using origin (version 6.1, Origin Software, San Clemente, CA).
RESULTS
The amino acid composition of experimental diets was also affected by the substitution of fish meal by locust meal in Table 2. Arginine, histidine, isoleucine, leucine, lysine, methionine, cystine, phenylalanine, tyrosine, threonine, tryptophan and valine contents of the experimental diets were not significantly affected among the varying replacement groups and found to be in the range of 27.1-27.5, 9.8-9.9, 14.1-14.9, 24.6-24.7, 19.7-22.3, 6.7-6.9, 2.7-3.3, 13.7-14.8, 14-14.3, 12.9-13.3, 2.4-3.3 and 17.4-17.5 g kg–1 dry matter. The fatty acid composition of the test diets was also not affected significantly except eicosapentaenoic acid (EPA), docosahexaenoic acid (DHA) and docosapentaenoic acid (DPA) which were found to decrease in D5, D6 and D7 significantly (p>0.05) were in 40, 50 and 60% fish meal protein in Table 3. The EPA values in fish fed D5, D6 and D7 diets were recorded to be 12.4, 11.3 and 10.1 g kg–1, respectively.
| Table 3: | Fatty acids profile of the experimental diets (g kg–1) | |||||||
Experimental diets | ||||||||
| Fatty acid | D1 | D2 | D3 | D4 | D5 | D6 | D7 | |
| Sat | ||||||||
| Myristic | 14:00 | 9.3±0.2 | 9.0±0.1 | 8.6±0.2 | 8.2±0.1 | 7.8±0.1 | 7.5±0.1 | 7.1±0.1 |
| Palmitic acid | 16:00 | 65.7±1.0 | 69.3±5.0 | 72.8±7.0 | 76.4±4.0 | 80.0±2.0 | 83.5±5.0 | 87.1±7.0 |
| Stearic acid | 18:00 | 24.0±1.0 | 21.2±5.0 | 22.0±3.0 | 22.9±3.0 | 23.7±1.0 | 24.6±5.0 | 25.4±4.0 |
| Mon | ||||||||
| Palmitoleic acid | 16:1 n-7 | 16.2±2.0 | 15.4±2.0 | 14.6±1.0 | 13.8±3.0 | 13.0±4.0 | 12.2±1.0 | 11.4±3.0 |
| Oleic acid | 18:1 n-9 | 142.1±8.0 | 146.2±6.0 | 150.2±4.0 | 154.3±3.0 | 158.4±1.0 | 162.5±8.0 | 166.6±5.0 |
| Gadoleic acid | 20:1 n-11 | 15.7±2.0 | 14.6±3.0 | 13.6±5.0 | 12.5±3.0 | 11.5±3.0 | 10.5±3.0 | 9.4±0.1 |
| Erucic acid | 22:1 n-9 | 12.3±0.2 | 11.3±0.1 | 10.3±0.2 | 9.3±0.4 | 8.4±0.1 | 7.4±0.1 | 6.4±0.3 |
| n-3 LC-PUFA | ||||||||
| Linoleic acid (LA) | 18:2 n-6 | 189.4±7.0 | 189.6±6.0 | 189.7±3.0 | 189.8±3.0 | 189.9±3.0 | 190.0±8.0 | 190.2±5.0 |
| Gamma-linolenic acid (GLA) | 18:3 n-6 | 0.3±0.0 | 0.3±0.01 | 0.3±0.1 | 0.3±0.0 | 0.3±0.1 | 0.3±0.0 | 0.3±0.0 |
| Arachidonic acid | 20:4 n-6 | 1.8±0.2 | 1.8±0.1 | 1.8±0.1 | 1.8±0.3 | 1.8±0.2 | 1.8±0.3 | 1.7±0.5 |
| Alpha-linolenic acid (ALA) | 18:3 n-3 | 23.8±1.0e | 25.8±2.0de | 27.8±1.0d | 29.8±4.0d | 31.8±2.0c | 33.8±3.0b | 35.8±0.1a |
| Stearidonic acid | 18:4 n-3 | 3.2±0.1 | 3.0±0.1 | 2.7±0.3 | 2.5±0.1 | 2.2±0.1 | 2.0±0.4 | 1.7±0.2 |
| Eicosapentaenoic acid (EPA) | 20:5 n-3 | 16.9±2.0a | 15.8±1.0a | 14.7±2.0ab | 13.5±4.0ab | 12.4±5.0b | 11.3±1.0c | 10.1±3.0d |
| Docosapentaenoic acid (DPA) | 22:5 n-3 | 5.7±0.1a | 5.3±0.1a | 4.8±0.4a | 4.4±0.2ab | 3.9±0.1b | 3.5±0.2c | 3.0±0.2d |
| Docosahexaenoic acid (DHA) | 22:6 n-3 | 22.0±4.0a | 20.6±3.0a | 19.2±1.0a | 17.8±3.0ab | 16.3±1.0b | 14.9±1.0c | 13.5±5.0d |
| 1Mean values of 3 replicate ±SEM, 2Not statistically significant (p>0.05) and alphabetical letters showed different significance level | ||||||||
| Table 4: | Growth performance and per cent survival of fingerling Oreochromis niloticus fed diets containing varying replacement levels of fish meal by locust meal1,2 | ||||||
Varying replacement levels of fish meal by locust meal | |||||||
D1 | D2 | D3 | D4 | D5 | D6 | D7 | |
| Initial weight (g fish–1) | 3.6±0.03 | 3.5±0.04 | 3.6±0.03 | 3.6±0.01 | 3.4±0.01 | 3.4±0.05 | 3.5±0.04 |
| Final weight (g fish–1) | 29.22±0.4a | 28.34.±0.3a | 29.51±0.1a | 27.42±0.3b | 21.01±0.5c | 17.13±0.3d | 12.69±0.5e |
| Absolute weight gain (g fish–1) | 25.62±0.2a | 24.84±0.4a | 25.91±0.1a | 23.82±0.4b | 17.61±0.6c | 13.73±0.5d | 9.19±0.5e |
| Live weight gain (%)3 | 711.7±1.6a | 709.7±1.1a | 719.7±2.4a | 661.6±3.6b | 517.9±4.3c | 403.8±3.2d | 262.6±2.4e |
| Feed conversion ratio4 | 1.64±0.06d | 1.61±0.02d | 1.58±0.03d | 1.62±0.04d | 1.98±0.03c | 2.54±0.02b | 3.18±0.01a |
| Specific growth rate (% day–1) | 3.74±0.05a | 3.73±0.01a | 3.76±0.05a | 3.63±0.02b | 3.25±0.03c | 2.89±0.05d | 2.30±0.03e |
| Protein efficiency ratio5 | 1.36±0.03b | 1.38±0.02b | 1.41±0.03a | 1.37±0.01c | 1.12±0.05c | 0.87±0.03d | 0.70±0.01d |
| Protein deposition (%)6 | 20.37±0.2a | 20.14±0.2a | 20.94±0.1a | 20.30±0.3a | 15.22±0.1b | 11.07±0.2c | 8.05±0.2d |
| Protein gain (g fish–1) | 3.85±0.01a | 3.62±0.02b | 3.86±0.01a | 3.52±0.03b | 2.39±0.04c | 1.74±0.01d | 1.06±0.03d |
| Protein retention efficiency (%) | 9.17±0.5a | 9.06±0.3a | 9.42±0.2a | 9.13±0.1a | 6.85±0.5b | 4.98±0.4c | 3.62±0.2d |
| Survival (%) | - | 93±2.6 | 100 | 97±3.2 | 100 | 100 | 100 |
| 1Mean values of 3 replicate ±SEM, 2Not statistically significant (p>0.05) and alphabetical letters showed different significance level | |||||||
The fish fed diets D5, D6 and D7 exhibited the DPA content 3.9, 3.5 and 3.0 g kg–1, respectively. Similarly lower values of DHA (16.3, 14.9 and 1350 g kg–1) in groups fed on D5, D6 and D7 diets was noted.
Over the 8 weeks feeding trial. Replacement of fish meal by locust meal on protein to protein basis was found to be feasible up to 30% as evident by insignificant differences (p<0.05) among the live weight gain (661.6-719.7%), feed conversion ratio (1.58-1.64), the protein efficiency ratio (1.36-1.41), specific growth rate (3.63-3.76%) and protein deposition (20.14-20.94%) of fish fed diets D1, D2, D3 and D4 wherein 0, 10, 20 and 30% fish meal protein was replaced by locust meal protein in Table 4. However, further replacement of fish meal by locust meal protein (beyond 30%) resulted in a significant decrease (p<0.05) in growth and conversion efficiencies. Significantly (p>0.05) poorest LWG (262%), FCR (3.18), low PER (0.70), SGR (2.30%) and PD (8.05) were detected in fish fed diet D7 wherein 60% fish meal protein is replaced with locust meal protein. Based on the broken-line analysis of LWG, the optimum replacement level was found to be 29.8% in Fig. 1.
The body composition of the fish was significantly altered by the different replacement levels of locust meal in Table 5. No remarkable differences in moisture content (772-779 g kg–1 wet weight basis) were evident in fish fed diets D1, D2, D3 and D4. However, remarkable variations in body moisture were detected in fish fed diets D5 (753 g kg–1), D6 (742 g kg–1) and D7 (731 g kg–1) compared to that of D1, D2, D3 and D4. Ash content differed insignificantly (21.6-22.1 g kg–1 wet weight basis) among the groups.
| Table 5: | Carcass composition (g kg–1 wet weight) of fingerling Oreochromis niloticus fed diets containing varying replacement levels of fish meal by locust meal1,2 | |||||||
Varying replacement levels of fish meal by locust meal | ||||||||
Initial | D1 | D2 | D3 | D4 | D5 | D6 | D7 | |
| Moisture | 748.0±22.0 | 779.4±5.6a | 771.1±8.3a | 774.3±9.7a | 772.1±7.5a | 753.6±5.4b | 742.1±7.2c | 731.1±7.8d |
| Crude protein | 108.0±3.0 | 145.1±5.2a | 141.2±0.5a | 143.9±0.1a | 142.7±0.9b | 131.1±0.8c | 122.8±0.6d | 113.2±0.4e |
| Crude fat | 43.0± 2.0 | 33.7±3.8e | 34.5±0.4d | 34.1±0.5de | 35.2±0.8d | 46.1±0.3c | 49.1±0.4b | 54.7±0.6a |
| Ash | 23.0± 0.2 | 21.6±1.8 | 22.1±0.5 | 21.7±0.3 | 21.9±0.6 | 22.1±0.6 | 21.5±0.4 | 21.6±0.2 |
| 1Mean values of 3 replicate ±SEM, 2Not statistically significant (p>0.05) and alphabetical letters showed different significance level | ||||||||
| Table 6: | Hematological indices and antioxidant status of fingerling Oreochromis niloticus fed diets containing varying replacement levels of fish meal by locust meal1,2 | ||||||
Varying replacement levels of fish meal by locust meal | |||||||
D1 | D2 | D3 | D4 | D5 | D6 | D7 | |
| Hematocrit (%) | 31.24±0.4a | 30.11±0.3a | 29.88±0.1a | 31.35±0.5a | 27.18±0.2b | 21.68±0.1c | 16.37±0.3d |
| Red blood corpuscles (106 mm–3) | 2.18±0.2a | 2.32±0.1a | 2.27±0.1a | 2.19±0.3a | 1.82±0.2b | 1.63±0.1c | 1.35±0.4d |
| Hemoglobin (g dL–1) | 9.81±0.1a | 9.38±0.9a | 9.11±0.4a | 9.72±0.7a | 8.62±0.8b | 7.19±0.7c | 6.12±1.2d |
| Superoxide dismutase (U mg–1 protein) | 82.4±13.0a | 80.6±12.0a | 79.4±16.0a | 71.2±24.0a | 63.7±13.0b | 53.9±15.0c | 45.1±12.0d |
| Catalase (U mg–1 protein) | 34.1±4.0a | 31.5±5.0a | 35.2±3.0a | 28.3±9.0a | 24.8±4.0b | 19.3±4.0c | 13.6±5.0d |
| Glutathione peroxidase activity (U mg–1 protein) | 187.3±24.0a | 182.4±26.0a | 185.6±23.0a | 173.9±1.2a | 164.1±33.0b | 151.7±44.0c | 139.2±51.0d |
| 1Mean values of 3 replicate ±SEM, 2Not statistically significant (p>0.05) and alphabetical letters showed different significance level | |||||||
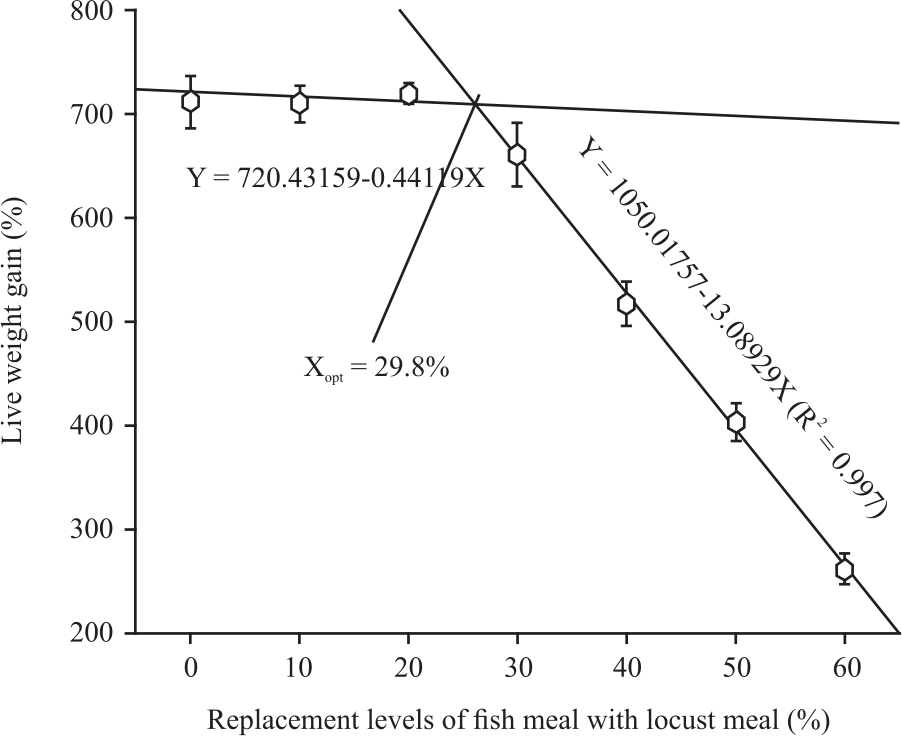 |
| Fig. 1: | Broken-line relationship of live weight gain to replacement levels of fish meal with locust meal for fingerling Oreochromis niloticus |
No significant differences amongst the body protein (141-145 g kg–1 wet basis) of the fish fed diets of D1, D2, D3 and D4 replacing 0, 10, 20 and 30%fish meal protein was evident. Whereas in fish fed diet D7 replacing 60% fish meal showed a sharp (p<0.05) decline (113 g kg–1 wet basis) in body protein. No remarkable differences (p>0.05) among body fat were evident in fish fed diets D1, D2, D3 and D4 (33.7-34.5 g kg–1 wet basis). However, a significant increase (p<0.05) in body fat was noted in fish fed dietsD5 (46.1 g kg–1), D6 (49.1 g kg–1) and D7 (54.7 g kg–1) replacing 40, 50 and 60% fish meal protein. In the current investigation, survival was not significantly affected among the variable treatments and was found to be in the range of 93-100% (Table 1).
The effects of different replacing levels of fish meal by locust meal on liver SOD, CAT, GPx activities and haematological (Hct, RBCs and Hb) tools in fingerling O. niloticus is mentioned in Table 6. The almost constancy (p<0.05) of liver SOD, CAT and GPx activities with the hiking replacing values of fish meal by locust meal up to 30% was recorded and, thereafter, a declining trend was noted in fish fed diets D5, D6 and D7 wherein 40, 50 and 60% fish meal protein was replaced by locust meal protein. The poor values of SOD (45.1 U mg–1 protein), CAT (13.6 U mg–1 protein) and GPx activities (139.2 U mg–1 protein)was recorded in fish fed D7 diet.The stable response of Hct, RBCs and Hb were noted in fish fed diets from 10 to 30% replacement of fish meal by locust meal. While the declining trend for blood tools was recorded on further replacement of fish meal by locust meal. The lowest values of Hct (16.3%), RBCs (1.35 ×106 mm–3) and Hb (6.12 g dL–1) were recorded in fish fed diet D7.
DISCUSSION
Choosing feed ingredients is a crucial parameter in the design and making of supplemented quality feed products of any aquatic species51-56. Fish meal is an important ingredient that is widely used as a source of animal protein has the high protein and essential amino acids but a higher price than available sources of plant protein57-61. Besides this, the accessibility of fish meal is falling day by day due to its high requirement in other than aquaculture industry like livestock, poultry etc. The reduced stock of fish meal in future will intensely affect fish production. Considering this, it is essential to partially reduce or eliminate fish meal in the fish diet. The method to reduce fish meal from fish diets is to replace it with alternate protein which should be less expensive and easily accessible, allowing for continued expansion of aquaculture. Because of this, various plant and animal protein sources have been assessed for the replacement of fish meal4,9,54,59,61-65. Locust meal is a good source of protein, amino acids and fatty acids profile21,66-68. Several workers have attempted to replace a fish meal with a locust meal in the past19-22. Amino acids, because they influenced fish growth and price, have found great value in fish nutrition studies69. Fish in the first months of life grow rapidly and that is why the supply of amino acids at this time is essential for body protein and energy70. Amino acids are the crucial substrates for protein synthesis and essential flow regulators in several metabolic pathways71. An imbalance of the amino acid diet pattern can lead to a fall in dietary intake and protein consumption72,73. Also, essential fatty acids in the diet must be taken into account when determining the nutritional value of the ingredients and feeds74. The need for essential fatty acids can only be met by providing long chain (LC) Polyunsaturated fatty acids (PUFA) in the diet, especially α-linolenic acid (LNA, 18: 3ω3) and linoleic acid (LA, 18: 2ω6) and various eicosapentaenoic acid (EPA, 20: 5ω3) and docosahexaenoic acid (DHA, 22: 6ω3) depending on the species72,75. In this study, however, the amino acid composition was not significantly affected among the varying replacement groups. The diets D1, D2 and D3 with 10, 20 and 30% replacing fish meal with locust meal showed the balanced fatty acid profile of the experimental diets in these groups. Whereas diets D5, D6 and D7 significantly (p>0.05) wherein 40, 50 and 60% fish meal protein are replaced by locust meal protein have the lower values of EPA, DHA and DPA leading to an imbalanced fatty acid profile for the groups which fed on these diets. Data of the present study revealed that 30% of the fish meal protein could be replaced by locust meal without hampering the growth of the O. Niloticus suggesting that the amino acids and fatty acid profile of the diets up to 30% replacement of fish meal protein with locust meal protein was as per the need of the fish. However, the further substitution of fish meal with locust meal protein in diet D5, D6 and D7 depressed growth and conversion efficiencies. This reduced performance at higher replacement of fish meal by locust meal (D5, D6 and D7) may be due to the lower amounts of eicosapentaenoic acid (EPA), docosapentaenoic acid (DPA) and docosahexaenoic acid (DHA)in the D5, D6 and D7 diets. In the present study, fish showed a relatively small but significant decrease (p<0.05) in SGR with increasing replacement of fish meal with locust meal beyond 30%. This may be because of the nutritional imbalance particularly the shortage of the essential fatty acids at higher levels of locust meal inclusion in the diet. High FCR noted at higher inclusion of locust meal in D7 (60% replacement) maybe because of the low nutrient utilization from locust meal at higher inclusion level. A similar observation on replacing fish meal with locust meal was also found for other aqua species20-22. Related outcomes were also detected by Brah et al.76, where in body weight and weight gain significantly fall with increasing higher substitution rates of fish meal with grasshopper meal. Deng et al.77 have also documented that decreased final body weight, AWG and SGR were noted with the increasing levels of replacement of fish meal by locust meal in broiler diet. Tran et al.67 conveyed that locust meals may be utilized to replace part of fish meal in fish diets.
Antioxidant power is important for maintaining complex fish antibodies, which include enzymatic and non-enzymatic antioxidant activities. Antioxidant enzymes including SOD, catalase and GPx form the first line of enzymatic defence against free radicals in organisms43,77,78. GPx involves in the exclusion of hydroxyl free radicals (OH°) and promotes the decomposition of hydrogen peroxide in water and oxygen and restores lipid hydroperoxide79. The SOD and GPx and catalase (CAT) are the main antioxidant enzyme group that promotes the conversion of reactive oxygen species to less reactive species80,81. Superoxide dismutase is considered to be the first line of protection against oxygen82,83. Cao et al.84 reported that excessive replacement of fish meal by maggot meal would make shrimp in a state of oxidative stress. Using more than 4.9% yellow mealworm Tenebrio molitor meal as a replacement for a fish meal might affect antioxidant enzyme activities of pearl gentian grouper, Epinephelus lanceolatus ♂ × Epinephelus fuscoguttatus ♀85. Dietary replacement of FM by 3% yeast hydrolysate could improve the antioxidant capability of juvenile Jian carp86. In the current research, an almost constancy (p<0.05) of liver SOD, CAT and GPx activities with the enhancing replacing values of fish meal by locust meal up to 30% was recorded and, thereafter, the declining trend was noted in fish fed diets D5, D6 and D7 wherein 40, 50 and 60% fish meal protein was replaced by locust meal protein.
Haematological analyses may indicate the health status of fish73,87. A haematological examination of fish demonstrated the stable response of Hct, RBCs and Hb in fish fed diets D1, D2, D3 and D4 with 0-30% replacement of fish meal by locust meal. While the declining trend for blood tools was recorded on further replacement of fish meal by locust meal (D5, D6 and D7). Feeding of dried locusts for 91 days did not harm haematological parameters but resulted in a slight reduction in the number and diminished ovarian steroidogenesis, which could reduce fertility88-90. Migratory locust food (Locusta migratoria) can replace up to 25% of the fishmeal in the isoproteic diet of Nile tilapia fingerlings without harmful influences on digestive function, growth function and haematological tools91,92.
Except for growth fall at higher amounts of locust inclusion in fish fed D5, D6 and D7, no diet linked mortality or any other nutritional pathologies were perceived in this research. The reverse relationship between growth parameters and higher replacement levels of fish meal by locust in fish diets has been noted in several previous researches18,20-22,93. Replacement of fish meal with locust meal of Nile tilapia may be affected by several biological factors such as species, life stage, sex, feeding habits, diet composition, resembles studies reported of replacing fish meal with locust meal in several other studies in broilers67,76,94,95.
CONCLUSION
The current experiment showed that a 30% inclusion level of locust meal produced the best result in terms of growth. It is, therefore, recommended that locust meal can be incorporated up to 30% as a replacement for fish meal without compromising the growth of fish. There are a variety of protein sources that can be used in aquaculture without affecting growth function, feed performance and body composition, given the need for amino acids, locust meal can be used as another protein ingredient in fish food.
SIGNIFICANCE STATEMENT
This study discovered that locust meal could be beneficial for the Nile tilapia fingerlings Oreochromis niloticus as an alternative protein source to fish meal in the limit of 30% in the diet, to formulate cost-effective commercial feeds for Oreochromis niloticus.
ACKNOWLEDGMENT
The authors are thankful to the Head of Department of Fisheries and Wildlife Science, Sudan University of Science and Technology for providing necessary facilities and the Ministry of Higher Education and Scientific Research, Sudan for conducting this experiment.
REFERENCES
- Noori, N.A., 2013. Effect of replacement of fish meal by soybean on growth, survival, feed utilization and production cost of fingerlings common carp (Cyprinus carpio L.) reared in the float cages. Int. J. Recent Sci. Res., 4: 308-312.
Direct Link - Zhang, L., X.D. Yang, Y.Y. Zhang, J. Yang and G.X. Qi et al., 2014. Changes in oleic acid content of transgenic soybeans by antisense RNA mediated posttranscriptional gene silencing. Int. J. Genomics, Vol. 2014.
CrossRefDirect Link - Dawood, M.A.O., S. Koshio, M. Ishikawa and S. Yokoyama, 2015. Effects of partial substitution of fish meal by soybean meal with or without heat-killed Lactobacillus plantarum (LP20) on growth performance, digestibility, and immune response of amberjack, Seriola dumerili juveniles. Bio. Med. Res. Int., Vol. 2015.
CrossRefDirect Link - Yue, Y.R. and Q.C. Zhou, 2008. Effect of replacing soybean meal with cottonseed meal on growth, feed utilization and hematological indexes for juvenile hybrid tilapia, Oreochromis niloticus × O. aureus. Aquaculture, 284: 185-189.
CrossRefDirect Link - Rathinam, A.M.M., K.K. Joe, P. Vijayagopal, J. Vidya, V.A. Leslie and R. Sundar, 2014. Effect of dietary protein levels in the formulated diets on growth and survival of juvenile spiny lobster Panulirus homarus (linnaeus). Ind. J. Fish., 61: 67-72.
Direct Link - Tunde, A.B., M.P. Kuton, A.A. Oladipo and L.H. Olasunkanmi, 2015. Economic analyze of costs and return of fish farming in Saki-East local government area of Oyo state, Nigeria. J. Aquacult. Res. Dev., Vol. 6.
CrossRefDirect Link - Kurbanov, A.R., R.Y. Milusheva, S.S. Rashidova and B.G. Kamilov, 2015. Effect of replacement of fish meal with silkworm (Bombyx mori) pupa protein on the growth of Clarias gariepinus fingerling. Int. J. Fish Aqua. Stud., 2: 25-27.
Direct Link - Yıldırım, Ö., Ü. Acar, A. Türker, M.C. Sunar and O.S. Kesbiç, 2014. Effects of replacing fish meal with peanut meal (Arachis hypogaea) on growth, feed utilization and body composition of mozambique tilapia fries (Oreochromis mossambicus). Pak. J. Zool., 46: 497-502.
Direct Link - Sitja-Bobadilla, A., S. Pena-Llopis, P. Gomez-Requeni, F. Medale, S. Kaushik and J. Perez-Sanchez, 2005. Effect of fish meal replacement by plant protein sources on non-specific defence mechanisms and oxidative stress in gilthead sea bream (Sparus aurata). Aquaculture, 249: 387-400.
CrossRef - Banjo, A.D., O.A. Lawal and E.A. Songonuga, 2006. The nutritional value of fourteen species of edible insects in Southwestern Nigeria. Afri. J. Biotech., 5: 298-301.
Direct Link - Makkar, H.P.S., G. Tran, V. Heuze and P. Ankers, 2014. State-of-the-art on use of insects as animal feed. Anim. Feed Sci. Technol., 197: 1-33.
CrossRefDirect Link - Koutsos, L., A. McComb and M. Finke, 2019. Insect composition and uses in animal feeding applications: A brief review. Ann. Entomol. Soc. Am., 112: 544-551.
CrossRefDirect Link - van Huis, A., J. van Itterbeeck, H. Klunder, E. Mertens, A. Halloran, G. Muir and P. Vantomme, 2013. Edible Insects: Future Prospects for Food and Feed Security. FAO, Rome, Italy, ISBN: 978-92-5-107595-1, Pages: 187.
Direct Link - Finke, M.D., 2002. Complete nutrient composition of commercially raised invertebrates used as food for insectivores. Zoo Biol., 21: 269-285.
CrossRefDirect Link - Longvah, T., K. Mangthya and P. Ramulu, 2011. Nutrient composition and protein quality evaluation of eri silkworm (Samia ricinii) prepupae and pupae. Food Chem., 128: 400-403.
CrossRefDirect Link - Mohamed, E.H.A., 2015. Determination of nutritive value of the edible migratory locust Locusta migratoria, Linnaeus, 1758 (Orthoptera: Acrididae). Int. J. Adv. Pharm. Biol. Chem., 4: 144-148.
Direct Link - Okoye, F.C. and J.C. Nnaji, 2005. Effects of substituting fish meal with grasshopper meal on growth and food utilization of the Nile tilapia, Oreochromis niloticus fingerlings. Proceedings of the 19th Annual Conference of Fisheries Society of Nigeria, November 29th-December 3rd, 2005, FISON, Ilorin, Nigeria.
Direct Link - Alegbeleye, W.O., S.O. Obasa, O.O. Olude, K. Otubu and W. Jimoh, 2012. Preliminary evaluation of the nutritive value of the variegated grasshopper (Zonocerus variegatus L.) for African catfish Clarias gariepinus (Burchell. 1822) fingerlings. Aquacult. Res., 43: 412-420.
CrossRefDirect Link - Buyukcapar, H.M., F. Gunlemez and A. Kamalak, 2012. Effect of partially replacing fish meal with honey locust seed (Gleditsia triacanthos) on growth, feed utilisation and body composition of mirror carp, Cyprinus carpio, fingerlings. J. Appl. Anim. Res., 40: 8-12.
CrossRefDirect Link - Grace, O.I., 2015. Effects of grasshopper meal in the diet of Clarias gariepinus fingerlings. J. Aquacult. Res. Dev., Vol. 6.
CrossRefDirect Link - Yousif, R.A., M.A. Khan and S. Zehra, 2021. Effect of replacing fishmeal with soybean meal on growth, feed conversion andcarcass composition of fingerling Oreochromis niloticus (Nile tilapia). SUST J. Agric. Vet. Sci., 22: 44-59.
Direct Link - Mzengereza, K., O.V. Msiska, F. Kapute, J. Kang’ombe, W. Singini and A. Kamangira, 2014. Nutritional value of locally available plants with potential for diets of Tilapia Rendalli in pond aquaculture in NkhataBay, Malawi. J. Aquacult. Res. Dev., Vol. 5.
Direct Link - Yakubu, A.F., N.A. Nwogu, J.O. Apochi, E.D. Olaji and T.E. Adams, 2014. Economic profitability of Nile tilapia (Oreochromis niloticus Linnaeus 1757) in semi flow through culture system. J. Aquat. Sci., 2: 1-4.
CrossRefDirect Link - Pai, I.K., M.S. Altaf and K.N. Mohanta, 2016. Development of cost effective nutritionally balanced food for freshwater ornamental fish black molly (Poecilia latipinna). J. Aquacult. Res. Dev., Vol. 7.
Direct Link - Sayed, J.A., H. Iqbal, H. Ayenuddin, S. Al-Amin and K. Samsad et al., 2018. Development of low cost formulated quality feed for growth performance and economics of Labeo rohita cultured in cage. AACL Bioflux., 11: 1486-1494.
Direct Link - Limbu, S.M., 2020. The effects of on-farm produced feeds on growth, survival, yield and feed cost of juvenile African sharptooth catfish (Clarias gariepinus). Aquacult. Fish., 5: 58-64.
CrossRefDirect Link - Bhujel, R.C., 2014. A Manual for Tilapia Business Management. CABI., United Kingdom, ISBN-13 9781780641362, Pages: 214.
CrossRefDirect Link - Chirapongsatonkul, N., N. Mueangkan, S. Wattitum and K. U-taynapun, 2019. Comparative evaluation of the immune responses and disease resistance of Nile tilapia (Oreochromis niloticus) induced by yeast β-glucan and crude glucan derived from mycelium in the spent mushroom substrate of Schizophyllum commune. Aquacult. Rep., Vol. 15.
CrossRefDirect Link - El-Sayed, A.F.M., 2006. Tilapia Culture. CABI Wallingford, UK, ISBN: 9780851990149, Pages: 275.
CrossRefDirect Link - Bhujel, R.C., D.C. Little and A. Hossain, 2007. Reproductive performance and the growth of pre-stunted and normal Nile tilapia (Oreochromis niloticus) broodfish at varying feeding rates. Aquaculture, 273: 71-79.
CrossRefDirect Link - Watanabe, W.O., T.M. Losordo, K. Fitzsimmons and F. Hanley, 2005. Tilapia production systems in the Americas: Technological advances, trends and challenges. Rev. Fish. Sci., 10: 465-498.
CrossRefDirect Link - Fitzsimmons, K., 2006. Prospect and Potential for Global Production. In: Tilapia Biology, Culture and Nutrition, Lim, C. and Webster, C.D. (Eds). Food Products Press, New York, USA., pp: 51-72.
Direct Link - Prabu, E., C.B.T. Rajagopalsamy, B. Ahilan, I.J.M.A. Jeevagan and M. Renuhadevi, 2019. Tilapia-an excellent candidate species for world aquaculture: A review. Ann. Res. Rev. Biol., 31: 1-14.
CrossRefDirect Link - Perera, A.D. and R.C. Bhujel, 2021. Field cricket (Gryllus bimaculatus) meal (FCM) to replace fishmeal in the diets for sex reversal and nursing of Nile tilapia (Oreochromis niloticus) fry. Aquacul. Res., 52: 4946-4958.
CrossRefDirect Link - Prabu, E. and A.A.V. Santhiya, 2016. An overview of bioremediation towards aquaculture. J. Aquacul. Trop., 31: 155-164.
Direct Link - Abdelghany, A.E., 2000. Optimum Dietary Protein Requirements for Oreochromisniloticus L. Fry Using Formulated Semi Purified Diet. In: K. Fitzsimmons and J.C. Filho (Eds.). Tilapia aquaculture in 21st century. Proc. from the 5th Intl. Symp. on tilapia in aquaculture. Rio de Janeiro, Brazil, 3-7 September 2000. 101-108. https://ag.arizona.edu/azaqua/tilapia/ista/ista5.htm
- Yousif, R.A., Masyamsir, Dhahiyat, Sunarto and Zahidah, 2017. Effect of physico-chemical conditions on the structure and composition of the phytoplankton community at jatinangor, Indonesia. Int. J. Curr. Microbiol. Appl. Sci., 6: 4188-4195.
Direct Link - Ahmed, I., 2009. Dietary total aromatic amino acid requirement and tyrosine replacement value for phenylalanine in Indian major carp: Cirrhinus mrigala (Hamilton) fingerlings. J. Appl. Ichthyol., 25: 719-727.
CrossRefDirect Link - Zehra, S. and M.A. Khan, 2019. Effects of different levels of dietary cyanocobalamin on growth, liver cyanocobalamin concentration, antioxidant capacity, intestinal enzymes and non-specific immune response for optimum inclusion in the commercial feeds of fingerling Channa punctatus (bloch). Aquaculture, Vol. 511.
CrossRefDirect Link - Vani, T., N. Saharan, S.D. Roy, R. Ranjan, A.K. Pal, G.M. Siddaiah and R. Kumar, 2012. Alteration in haematological and biochemical parameters of Catla catla exposed to sub-lethal concentration of cypermethrin. Fish Physiol. Biochem., 38: 1577-1584.
CrossRefDirect Link - Zehra, S. and M.A. Khan, 2014. Dietary phenylalanine requirement and tyrosine replacement value for phenylalanine for fingerling Catla catla (Hamilton). Aquaculture, 433: 256-265.
CrossRefDirect Link - Khan, Y.M. and M.A. Khan, 2020. Dietary folic acid requirement of fingerling catla, Catla catla (Hamilton). Aquacult. Nutr., 26: 1035-1045.
CrossRefDirect Link - Abdel-Hameid, N.A.H., S. Zehra and M.A. Khan, 2017. Dietary copper requirement of fingerling Channa punctatus (Bloch) based on growth, feed conversion, blood parameters and whole body copper concentration. Aquacul. Res., 48: 2787-2797.
CrossRefDirect Link - Qian, Y., X.F. Li, D.D. Zhang, D.S. Cai, H.Y. Tian and W.B. Liu, 2015. Effects of dietary pantothenic acid on growth, intestinal function, anti-oxidative status and fatty acids synthesis of juvenile blunt snout bream Megalobrama amblycephala. PLoS ONE, Vol. 10.
CrossRefDirect Link - Hardy, R.W. and S.J. Kaushik, 2021. Fish Nutrition. 4th Ed., Elsevier Science, Netherlands, ISBN: 9780128231593, Pages: 922.
CrossRefDirect Link - Robbins, K.R., A.M. Saxton and L.L. Southern, 2006. Estimation of nutrient requirements using broken-line regression analysis. J. Anim. Sci., 84: E155-E165.
PubMedDirect Link - Shearer, K.D., 2000. Experimental design, statistical analysis and modelling of dietary nutrient requirement studies for fish: A critical review. Aquacult. Nutr., 6: 91-102.
CrossRefDirect Link - Zamal, H., P. Barua, B. Uddin and K.S. Islam, 2008. Application of ipil-ipil leaf meal (IILM) as feed Ingredient for monosex tilapia fry (Oreochromis niloticus) in terms of growth and economics. Aquacul. Asia Maga., 5: 31-33.
Direct Link - Koumi, A.R., B.C. Atse and L.P. Kouame, 2009. Utilization of soya protein as an alternative protein source in Oreochromis niloticus diet: Growth performance, feed utilization, proximate composition and organoleptic characteristics. Afr. J. Biotechnol., 8: 91-97.
Direct Link - Shiu, Y.L., S.L. Hsieh, W.C. Guei, Y.T. Tsai, C.H. Chiu and C.H. Liu, 2013. Using Bacillus subtilis E20-fermented soybean meal as replacement for fish meal in the diet of orange-spotted grouper (Epinephelus coioides, Hamilton). Aquacult. Res., 46: 1403-1416.
CrossRefDirect Link - Aziza, A. and A.A. El-Wahab, 2019. Impact of partial replacing of dietary fish meal by different protein sources on the growth performance of Nile tilapia (Oreochromis niloticus) and whole body composition. J. Appl. Sci., 19: 384-391.
CrossRefDirect Link - Stenberg, O.K., E. Holen, L. Piemontese, N.S. Liland, E.J. Lock, M. Espe and I. Belghit, 2019. Effect of dietary replacement of fish meal with insect meal on in vitro bacterial and viral induced gene response in atlantic salmon (Salmo salar) head kidney leukocytes. Fish Shellfish Immunol., 91: 223-232.
CrossRefDirect Link - Vechklang, K., S. Boonanuntanasarn, S. Ponchunchoovong, N. Pirarat and C. Wanapu, 2011. The potential for rice wine residual as an alternative protein source in a practical diet for Nile tilapia (Oreochromis niloticus) at the juvenile stage. Aquacult. Nutr., 17: 685-694.
CrossRef - Wu, Y., H. Han, J. Qin and Y. Wang, 2014. Replacement of fishmeal by soy protein concentrate with taurine supplementation in diets for golden pompano (Trachinotus ovatus). Aquacult. Nutr., 21: 214-222.
CrossRefDirect Link - Sharawy, Z., A.M.A.S Goda and M.S. Hassaan, 2016. Partial or total replacement of fish meal by solid state fermented soybean meal with Saccharomyces cerevisiae in diets for Indian prawn shrimp, Fenneropenaeus indicus, postlarvae. Anim. Feed Sci. Technol., 212: 90-99.
CrossRefDirect Link - Devic, E., W. Leschen, F. Murray and D.C. Little, 2017. Growth performance, feed utilization and body composition of advanced nursing Nile tilapia (Oreochromis niloticus) fed diets containing black soldier fly (Hermetia illucens) larvae meal. Aquacult. Nutr., 24: 416-423.
CrossRefDirect Link - Shao, J., W. Zhao, S. Han, Y. Chen, B. Wang and L. Wang, 2018. Partial replacement of fishmeal by fermented soybean meal in diets for juvenile white shrimp (Litopenaeus vannamei). Aquacult. Nutr., 25: 145-153.
CrossRefDirect Link - Ju, Z.Y., D.F. Deng and W. Dominy, 2012. A defatted microalgae (Haematococcus pluvialis) meal as a protein ingredient to partially replace fishmeal in diets of Pacific white shrimp (Litopenaeus vannamei, Boone, 1931). Aquaculture, 354-355: 50-55.
CrossRefDirect Link - Macias-Sancho, J., L.H. Poersch, W. Bauer, L.A. Romano, W. Wasielesky and M.B Tesser, 2014. Fishmeal substitution with Arthrospira (Spirulina platensis) in a practical diet for Litopenaeus vannamei: Effects on growth and immunological parameters. Aquaculture, 426-427: 120-125.
CrossRefDirect Link - Ding, Z., Y. Zhang, J. Ye, Z. Du and Y. Kong, 2015. An evaluation of replacing fish meal with fermented soybean meal in the diet of Macrobrachium nipponense: Growth, nonspecific immunity, and resistance to Aeromonas hydrophila. Fish Shellfish Immunol., 44: 295-301.
CrossRefDirect Link - Yousif, R.A., A.M. Hamed, F.A. Dungos and G.A. Yagob, 2019. Effect of replacing fishmeal with baobab seed meal (Adansona digitata) on growth, feed conversion and carcass composition for Nile tilapia fry (Oreochromis niloticus). Egypt. Acad. J. Biol. Sci. B. Zool., 11: 97-105.
CrossRefDirect Link - Rumpold, B.A. and O.K. Schlüter, 2013. Potential and challenges of insects as an innovative source for food and feed production. Innovative Food Sci. Emerging Technol., Vol. 17.
CrossRefDirect Link - Tran, G., V. Heuzé and H.P.S. Makkar, 2015. Insects in fish diets. Anim. Frontiers, 5: 37-44.
CrossRefDirect Link - Paul, A., M. Frederich, R. Uyttenbroeck, S. Hatt and P. Malik et al., 2016. Grasshoppers as a food source? A review. Biotechnol. Agron. Soc. Environ., 20: 337-352.
Direct Link - Zehra, S. and M.A. Khan, 2014. Dietary tryptophan requirement of Fingerlingcatla catla (hamilton) based on growth, protein gain, RNA/DNA ratio, haematological parameters and carcass composition. Aquacult. Nutr., 21: 690-701.
CrossRefDirect Link - Terjesen, B.F., K.J. Lee, Y. Zhang, M. Failla and K. Dabrowski, 2006. Optimization of dipeptide-protein mixtures in experimental diet formulations for rainbow trout (Oncorhynchus mykiss) alevins. Aquaculture, Vol. 254. 254: 517-525.
CrossRefDirect Link - Jobgen, W.S., S.K. Fried, W.J. Fu, C.J. Meininger and G. Wu, 2006. Regulatory role for the arginine-nitric oxide pathway in metabolism of energy substrates. J. Nutr. Biochem., 17: 571-588.
CrossRefDirect Link - Mohammed, F.A., R.A. Yousif, F.M. Hilal, R.A. Adam and T.K. Ahmed, 2020. The effect of dietary methionine levels on growth, feed conversion and protein retention efficiency of Nile tilapia (Oreochromis niloticus) fingerlings. Nusantara Biosci., 12: 21-27.
CrossRefDirect Link - Zehra, S. and M.A. Khan, 2020. Dietary folic acid requirement of fingerling Channa punctatus (bloch) based on growth, protein productive value and liver folic acid concentrations. Anim. Feed Sci. Technol., Vol. 262.
CrossRefDirect Link - Doğan, G. and Ö.O. Ertan, 2017. Determination of amino acid and fatty acid composition of goldband goatfish [Upeneus moluccensis (bleeker, 1855)] fishing from the Gulf of Antalya (Turkey). Int. Aquat. Res., 9: 313-327.
CrossRefDirect Link - Hixson, S.M., 2014. Fish nutrition and current issues in aquaculture: The balance in providing safe and nutritious seafood, in an environmentally sustainable manner. J. Aquacult. Res. Dev., Vol. 5.
CrossRefDirect Link - Brah, N., F.M. Houndonougbo and S. Issa, 2018. Grasshopper meal (Ornithacris cavroisi) in broiler diets in Niger: Bioeconomic performance. Int. J. Poul. Sci., 17: 126-133.
CrossRefDirect Link - Deng, J., K. Mai, L. Chen, H. Mi and L. Zhang, 2015. Effects of replacing soybean meal with rubber seed meal on growth, antioxidant capacity, non-specific immune response, and resistance to Aeromonas hydrophila in tilapia (Oreochromis niloticus × O. aureus). Fish Shellfish Immunol., 44: 436-444.
CrossRefDirect Link - Ighodaro, O.M. and O.A. Akinloye, 2018. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alexandria J. Med., 54: 287-293.
CrossRefDirect Link - Liu, Y., J.Y. Wang, B.S. Li, H.J. Qiao, X.D. Liu, T.T. Hao and X.Y. Wang, 2018. Dietary manganese requirement of juvenile hybrid grouper, Epinephelus lanceolatus × E. fuscoguttatus. Aquacult. Nutr., 24: 215-223.
CrossRefDirect Link - Fang, K.M., F.C. Cheng, Y.L. Huang, S.Y. Chung, Z.Y. Jian and M.C. Lin, 2013. Trace element, antioxidant activity and lipid peroxidation levels in brain cortex of gerbils after cerebral ischemic injury. Biol. Trace Elem. Res., 152: 66-74.
CrossRefDirect Link - Tang, Q.Q., L. Feng, W.D. Jiang, Y. Liu and J. Jiang et al., 2013. Effects of dietary copper on growth, digestive and brush border enzyme activities and antioxidant defense of hepatopancreas and intestine for young grass carp (Ctenopharyngodon idella). Biol. Trace Elem. Res., 155: 370-380.
CrossRefDirect Link - Fattman, C.L., L.M. Schaefer and T.D. Oury, 2003. Extracellular superoxide dismutase in biology and medicine. Free Radical Biol. Med., 35: 236-256.
CrossRefPubMedDirect Link - Zhang, H., R. Sun, W. Xu, H. Zhou, W. Zhang and K. Mai, 2016. Dietary manganese requirement of juvenile large yellow croaker Larimichthys crocea (Richardson, 1846). Aquaculture, 450: 74-79.
CrossRefDirect Link - Cao, J.M., J. Yan, Y.H. Huang, G.X. Wang and R.B. Zhang et al., 2012. Effects of replacement of fish meal with housefly maggot meal on growthperformance, antioxidant and non-specific immune indexes of juvenile Litopenaeus vannamei. J. Fish. China, 36: 529-537.
CrossRefDirect Link - Song, S.G., S.Y. Chi, B.P. Tan, G.L. Liang and B.Q. Lu et al., 2018. Effects of fishmeal replacement by Tenebrio molitor meal on growth performance, antioxidant enzyme activities and disease resistance of the juvenile pearl gentian grouper (Epinephelus lanceolatus ♂ × Epinephelus fuscoguttatus ♀). Aquacult. Res., 49: 2210-2217.
CrossRefDirect Link - Yuan, X.Y., G.Z. Jiang, C.C. Wang, K.P. Abasubong, Q. Zou, Y.Y. Zhou and W.B. Liu, 2018. Effects of partial replacement of fish meal by yeast hydrolysate on antioxidant capability, intestinal morphology, and inflammation-related gene expression of juvenile jian carp (Cyprinus carpio var. jian). Fish Physiol. Biochem., 45: 187-197.
CrossRefDirect Link - Fazio, F., C. Faggio, S. Marafioti, A. Torre, M. Sanfilippo and G. Piccione, 2013. Effect of water quality on haematological and biochemical parameters of Gobius niger caught in Faro Lake (Sicily). Iran. J. Fish. Sci., 12: 219-231.
Direct Link - Johri, R., R. Singh and P.K. Johri, 2010. Effect of different formulated plant and animal diet on hematology of Clarias batrachus linn. under laboratory conditions. biochem. Cell Arch., 10: 283-291.
Direct Link - Johri, R., R. Singh and P.K. Johri, 2011. Studies on ovarian activity in formulated feed treated Clarias batrachus linn. J. Experi. Zool. Ind., 14: 111-115.
Direct Link - Johri, R., R. Singh and P.K. Johri, 2011. Histopathological examination of the gill, liver, kidney, stomach, intestine, testis and ovary of Clarias batrachus linn. during the feeding on different formulated feeds. J. Experi. Zool. Ind., 14: 77-79.
Direct Link - Sánchez-Muros, M.J., F.G. Barroso and C. de Haro, 2016. Brief Summary of Insect Usage as an Industrial Animal Feed/Feed Ingredient. In: Insects as Sustainable Food Ingredients. Dossey, A.T., J.A. Morales-Ramos and M.G. Rojas, Elsevier Netherlands Pages: 37.
CrossRefDirect Link - Ojha, S., A. El-Din Bekhit, T. Grune and O.K. Schlüter, 2021. Bioavailability of nutrients from edible insects. Curr. Opin. Food Sci., 41: 240-248.
CrossRefDirect Link - Mohammed, F.A., H.M.A. Suliman, R.A. Yousif, A.A. Mohamed, A.M. Alhafez and M.E. Rahma, 2020. Effect of different levels of locust meal on growth, feed conversion and carcass composition for nile tilapia fry (Oreochromis niloticus). Int. J. Oceanogr. Aquacult., Vol. 4.
CrossRefDirect Link - Sun, T., R.J. Long and Z.Y. Liu, 2013. The effect of a diet containing grasshoppers and access to free-range on carcase and meat physicochemical and sensory characteristics in broilers. Br. Poult. Sci., 54: 130-137.
CrossRefDirect Link - Nginya, E.S., J.O. Ondiek, A.M. King’ori and J.M. Nduko, 2019. Evaluation of grasshoppers as a protein source for improved indigenous chicken growers. Livest. Res. Rural Dev., Vol. 31.
Direct Link - Halver, J.E. and R.W. Hardy, 2002. Fish Nutrition. 3rd Ed., Academic Press, USA, ISBN: 9780080494920, Pages: 500.
Direct Link