
Tangu Nehemiah Njegwe
Department of Biochemistry, Faculty of Science, University of Bamenda, P.O. Box 39, Bambili, Cameroon
Kada Sanda Antoine
Department of Biological Sciences, Faculty of Science, University of Bamenda, P.O. Box 39, Bambili, Cameroon
Kaghou Mabel Mbifung
Department of Biochemistry, Faculty of Science, University of Bamenda, P.O. Box 39, Bambili, Cameroon
Njouonkou André-Ledoux
Department of Biological Sciences, Faculty of Science, University of Bamenda, P.O. Box 39, Bambili, Cameroon
Manfo Tsagué Faustin Pascal
Department of Biochemistry and Molecular Biology, Faculty of Science, University of Buea, P.O. Box 63, Buea, Cameroon
LiveDNA: 237.21514
Nono Vanessa Ninkeh
Department of Biochemistry, Faculty of Science, University of Bamenda, P.O. Box 39, Bambili, Cameroon
Mbouh Mariama
Department of Biochemistry, Faculty of Science, University of Bamenda, P.O. Box 39, Bambili, Cameroon
Nantia Akono Edouard
Department of Biochemistry, Faculty of Science, University of Bamenda, P.O. Box 39, Bambili, Cameroon
LiveDNA: 237.20868
Asian Journal of Animal and Veterinary Advances
Year: 2022 | Volume: 17 | Issue: 4 | Page No.: 146-156







ABSTRACT
Background and Objective: Pesticides largely used in agriculture and public health hygiene have shown negative effects on mammalian systems. Mushrooms thanks to their content in bioactive compounds have been considered as promising pharmacological sources. This study aimed at evaluating the protective effects of Ganoderma boninense aqueous extract (GBE) on the liver and reproductive parameters in Parastar-exposed male Wistar rats. Materials and Methods: Forty eight male rats (150±2 g) equitably divided into 6 groups were given either distilled water (5 mL kg–1), Parastar (6.23 mg kg–1), GBE (50, 150 mg kg–1) or Parastar (6.23 mg kg–1) and GBE (50, 150 mg kg–1) for 60 days. At the end of treatment, liver and reproductive organs weights were recorded and epididymal sperm characteristics determined. Liver and testis histology and biomarkers (testosterone levels, oxidative stress biomarkers, vesicular fructose) were also evaluated. Results: Treatment of animals with Parastar resulted in a decrease (p<0.05) of liver weights and increased (p<0.05) activities of alanine and aspartate aminotransferases. The insecticide decreased (p<0.05) epididymal sperm density and motility, serum testosterone and vesicular fructose levels. Moreover, Parastar increased lipid peroxidation, decreased glutathione levels, superoxide dismutase and catalase activities and induced histopathological changes in the testis and liver. Interestingly, all Parastar altered liver and reproductive parameters were counteracted with the co-administration of any of the two doses of GBE. Conclusion: This study provides evidence that G. boninense is a promising mushroom in protecting the liver and reproductive functions against Parastar-induced toxicity.
PDF Abstract XML References Citation
How to cite this article
Tangu Nehemiah Njegwe, Kada Sanda Antoine, Kaghou Mabel Mbifung, Njouonkou André-Ledoux, Manfo Tsagué Faustin Pascal, Nono Vanessa Ninkeh, Mbouh Mariama and Nantia Akono Edouard, 2022. Ganoderma boninense (Ganodermataceae) Aqueous Extract Protects Against Liver and Reproductive Alterations in Parastar-Exposed Male Wistar Rats. Asian Journal of Animal and Veterinary Advances, 17: 146-156.
DOI: 10.3923/ajava.2022.146.156
URL: https://scialert.net/abstract/?doi=ajava.2022.146.156
DOI: 10.3923/ajava.2022.146.156
URL: https://scialert.net/abstract/?doi=ajava.2022.146.156
INTRODUCTION
The occurrence of toxic chemicals in the environment is an issue of great concern1. Amongst environmental toxicants, are pesticides which are widely used in agriculture and veterinary medicine to boost productivity, particularly in developing countries including Cameroon2,3. They are used to protect crops against agricultural pests, diseases and weeds4. Though designed to cause harm or death to agricultural pests, these chemicals have been harmful to unintended species including humans and wild animals5. The use of pesticides is associated with health hazards including male reproductive dysfunction, hypertension, skin rashes, asthma, cancer, breathing difficulty, dizziness, vision problems, cough and gastrointestinal tract disorders, alteration of the immune, hormonal, neurological and digestive systems3,5.
Several mammalian physiological systems and organs are severely intoxicated by pesticides including affection of the reproductive and liver functions5,6. The pathogenicity of pesticides on the liver is commonly through the induction of cellular oxidative stress resulting from an increase in the production of reactive oxygen species (ROS) and alteration of the antioxidant defenses7,8. The liver plays an essential role in the detoxification of organisms and metabolism of xenobiotics such as pesticides and their metabolites9. Not surprisingly, the liver is a major non-reproductive target for sex steroid action and it interacts with reproductive system in a bidirectional fashion and its function is related to the reproductive axis10. Parastar like other pesticides could be hepatotoxic on non-target species with short and long term consequences although information on the hepatotoxicity of this insecticide is yet to be elucidated.
Parastar, an insecticide formulation that contains a pyrethroid lambda-cyhalothrin and neonicotinoid imidacloprid is largely used in the Santa sub-division of the North West region of Cameroon against insects on crops such as tomatoes, carrots, green leafy vegetables and spices3. Lambda-cyhalothrin acts on voltage-sensitive sodium channels by inducing repetitive activity on the nervous system especially the sensory nervous system as a result of prolonged closure time and increased in sodium permeability of the nerve membrane-associated with excitation, resulting in a reduction in action potential threshold and repetitive firing, paralysis or death of insects4,11. Imidacloprid binds and activates the nicotinic acetylcholine receptors (excitatory) affecting synaptic transmissions12. Despite these beneficial roles of Parastar, its excessive and non-controlled use affects non-targeted organisms through various exposures such as contact with surfaces sprayed with the chemicals or consumption of foods and water contaminated with the pesticide residues13. Exposure to pesticides is considered a risk factor for many diseases or conditions such as infertility, digestive tract and nervous system disorders4,13. As parastar is generally used in Cameroon for crop protection and yield, a practice that may lead to human exposure to the agrochemical among humans, searching for solutions to prevent, mitigate or counteract the negative effects of the agrochemical on farmer's health is therefore of great importance.
Mushrooms are not only utilized as food but also as a source for the development of new and safe drugs and nutraceuticals14,15. The mushroom Ganoderma boninense (G. boninense) is a basidiomycete white-rot fungus on oil palm. It belongs to the genus Ganoderma and is reported to be used in folk medicine for the treatment of gastric ulcer, chronic hepatitis, hypertension, nephritis, asthma, arthritis, bronchitis, insomnia, cancer, diabetes and anorexia16. The extract of G. boninense is reported to have antioxidant activity17. The antioxidant properties of G. boninense could support certain beneficial efficiency as scavengers or protective candidates against toxicants associated to oxidative stress. G. boninense may thus constitute a potentially useful resource for new and safe drugs for the treatment of various ailments, including liver toxicity and male reproductive dysfunctions. This study was therefore designed to investigate the effects of the aqueous extract of G. boninense on the liver and reproductive parameters in Parastar (insecticide) exposed male Wistar rats.
MATERIALS AND METHODS
Study area: The study was carried out at the Biochemistry Laboratory of the University of Bamenda (Cameroon), and some biochemical analyses were done at the Biotechnology Unit of the University of Buea (Cameroon) while histological analyses were carried out at the Histology Laboratory of the Evangelic University of Cameroon. The study was conducted from June-October, 2020.
Experimental animals: Forty-eight male Wistar rats (Rattus norvegicus) (2 months old, the average weight of 150±2 g) were obtained from the University of Dschang Animal Laboratory, West Region, Cameroon. The animals were housed under standard conditions (room temperature of about 25°C) with a 12 hrs light/12 hrs dark cycle and given food and water ad-libitum. They were handled according to the Cameroon National Veterinary Laboratory as a reference by the approval and health control No 003/19 CCS/MINEPIA/RD-NW/DD-ME/SSV. All the animals were acclimatized for two weeks before the beginning of the experiment.
Collection of G. boninense and preparation of aqueous extract: The wild fruiting bodies of G. boninense were collected from oil palm, in Foumban (West Region of Cameroon) in June 2019 and identified by a botanist and mycologist (Prof. Njouonkou Andre-Ledoux) of the Faculty of Science of the University of Bamenda.
The fresh mushrooms were washed with tap water to remove debris and epiphytes. Then, they were shredded into smaller pieces before being dried to a constant weight in an oven at 50°C. The oven-dried mushroom was crushed into powder using a blender and extraction proceeded as described by Mohammed et al.18 with some modifications. The dried sample (571 g) was macerated in 5 L of distilled water at room temperature for 24 hrs and filtered through Whatman #4 paper. Then, the process was repeated twice to maximize the extraction. The solvent was evaporated at 50°C to dryness in an oven and 55 g brown extract was obtained giving an extraction yield of 9.63%.
Chemicals: Parastar 40 WP containing imidacloprid (20 g kg–1) and lambda-cyhalothrin (20 g kg–1) was gotten from the local market of Santa (Cameroon). It is imported and distributed by FIMEX International SABP, Douala, Cameroon. ELISA testosterone kit was obtained from Calbiotech Inc. (Calbiotech, Inc., Spring Valley, CA) while albumin, cholesterol, Alanine Aminotransferase (ALT) and Aspartate Aminotransferase (AST) kits were manufactured and distributed by Biopharcam S.A (Bafoussam, Cameroon). Hydrogen peroxide, sodium carbonate and sodium hydroxide were obtained from BDH chemicals Ltd Poole (England). Trichloroacetic Acid (TCA) and Thiobarbituric Acid (TBA) were purchased from Sigma Aldrich (France) while 2,2-dithio-5,5’-dibenzoic acid was obtained from Burgoynes and Co. (Mumbai, India). Other chemicals were of analytical grades.
Experimental design: Forty-eight male Wistar rats were divided into six groups of 8 rats each which received daily either distilled water (5 mL kg–1), Parastar (6.23 mg kg–1), GBE (50, 150 mg kg–1) or Parastar (6.23 mg kg–1) and GBE (50, 150 mg kg–1), respectively for 60 days. The choice of this period was guided by the time for complete spermatogenesis in rats, which is 53-60 days19. The dose of Parastar (6.23 mg kg–1) was based on the toxicity of its constitutive ingredients, LD50 (lethal dose 50) values as described previously13. Meanwhile, the doses of mushroom GBE extract (50, 150 mg kg–1) were defined from previous studies on animal model20. Throughout the experimental duration, the animals were given food and water ad-libitum and body weights were recorded once every 2 days. At the end of the treatment, the rats were anaesthetized, sacrificed and blood samples were collected for serum preparation. The animals’ liver and reproductive organs (testes, epididymides, seminal vesicles and ventral prostates) were dissected out and weighed. A 20% (20 W/V) homogenate of the organs (liver, right testis, seminal vesicle) were prepared in a phosphate buffer (0.1M, pH 7.2) and used for biochemical analyses. The left testis and a portion of the liver were kept in 10% of formalin for histological analysis. The cauda epididymis was chopped in 0.9% sodium chloride at 37°C and the motility and density of emerged sperm cells were evaluated under 400× magnification using an ordinary light microscope, according to the method described by Ngoula et al.21.
Determination of biochemical parameters: Serum was used to assess testosterone levels and AST and ALT activities, albumin and total cholesterol levels following the manufacturer’s procedures. In liver and testicular homogenates, levels of glutathione (GSH) and Thiobarbituric Acid Reactive Substances (TBARS) were determined as previously reported by Kada et al.22. The activities of superoxide dismutase and catalase were determined in the homogenates using the methods described elsewhere22,23. The biochemical parameters in the liver and testis were corrected using protein levels determined with bovine serum albumin as standard22. Seminal vesicle fructose concentrations was determined according to the method previously described24, using fructose as standard.
Histopathological examination of liver and testis: A small portion of liver or the left testis was fixed in 10% neutral buffered formalin and then dehydrated with grades of ethanol (70, 80, 90, 95 and 100%). Dehydration was then followed by clearing the samples in 2 changes of xylene. Samples were impregnated with 2 changes of molten paraffin wax, then embedded and blocked out. Sections of 5 μm thickness were cut using a rotary microtome and mounted on glass slides. The slides were stained with haematoxylin and eosin and the stained sections were examined under the light microscope (OLYMPUS, Japan) for histopathological changes.
Statistical analysis: Data were analyzed by one-way analysis of variance using Dunnett’s multivariate test followed by a mean comparison between different groups using Student-Newman-Keuls test (parametric test) or Games-Howell’s test (nonparametric test) when appropriate. The analyses were performed using Statistical Package for Social Science (SPSS) version 26. The p<0.05 were considered statistically significant.
RESULTS
Animal body weight: The body weight of animals exposed to Parastar alone, GBE alone or co-administration of the insecticide and GBE in animals for 64 days is presented in Fig 1. In general, the animal body weight increased over time and there was no difference among treatments.
Liver and reproductive organ weights: The relative weight of the liver and reproductive organs from rats exposed to Parastar alone, GBE alone or co-treatment are presented in Table 1. Exposure of animals to Parastar resulted in a significant decrease (p<0.05) in liver weight but did not affect the weights of reproductive organs (testes, epididymides, seminal vesicles and prostate) when compared to the negative control receiving distilled water. Treatment of male rats with the two doses of GBE normalized the relative weights of the liver and reproductive organs as compared to the group given distilled water. Co-treatment of rats with Parastar and both doses of GBE increased (p<0.05) the epididymis weight while the addition of 50 mg kg–1 GBE to Parastar increased (p<0.05) the weight of the seminal vesicles when compared to the group given Parastar only. Co-treatment of animals with Parastar and the two doses of GBE counteracted the decreasing effect of Parastar on the liver weight as compared to the Parastar treated group.
Serum albumin, cholesterol levels and serum aminotransferase activities: The serum levels of albumin and cholesterol and the serum aminotransferase activities of rats exposed to either Parastar alone, GBE alone or co-treatment are summarized in Table 2. Parastar significantly decreased (p<0.05) serum albumin levels in animals as compared to the group given distilled water. Treatment of rats with any of the two doses of GBE normalized serum albumin levels when compared to the distilled water control group (Table 2).
Parastar treatment increased serum cholesterol levels in male rats when compared to the group given distilled water. Treatment of animals with 150 mg kg–1 GBE decreased (p<0.05) serum cholesterol as compared to the distilled water control group. Concomitant administration of Parastar and any of the two doses of GBE to the animals significantly (p<0.05) mitigated the negative effect of Parastar on serum cholesterol levels when compared to groups given Parastar alone and distilled water (Table 2).
Administration of Parastar to animals significantly increased (p<0.05) their ALT and AST activities as compared to the group given distilled water. Treatment of rats with any of the two doses of GBE slightly did not affect serum ALT and AST activity as compared to the control group given distilled water. Co-treatment of 150 mg kg–1 GBE and Parastar significantly prevented the insecticide-induced high ALT and AST activity (Table 2).
Levels of serum testosterone, vesicular fructose and sperm characteristics: The serum testosterone, vesicular vesicular fructose concentrations and epididymal sperm characteristics of rats exposed to either Parastar alone, GBE alone or co-treatment are summarized in Table 3.
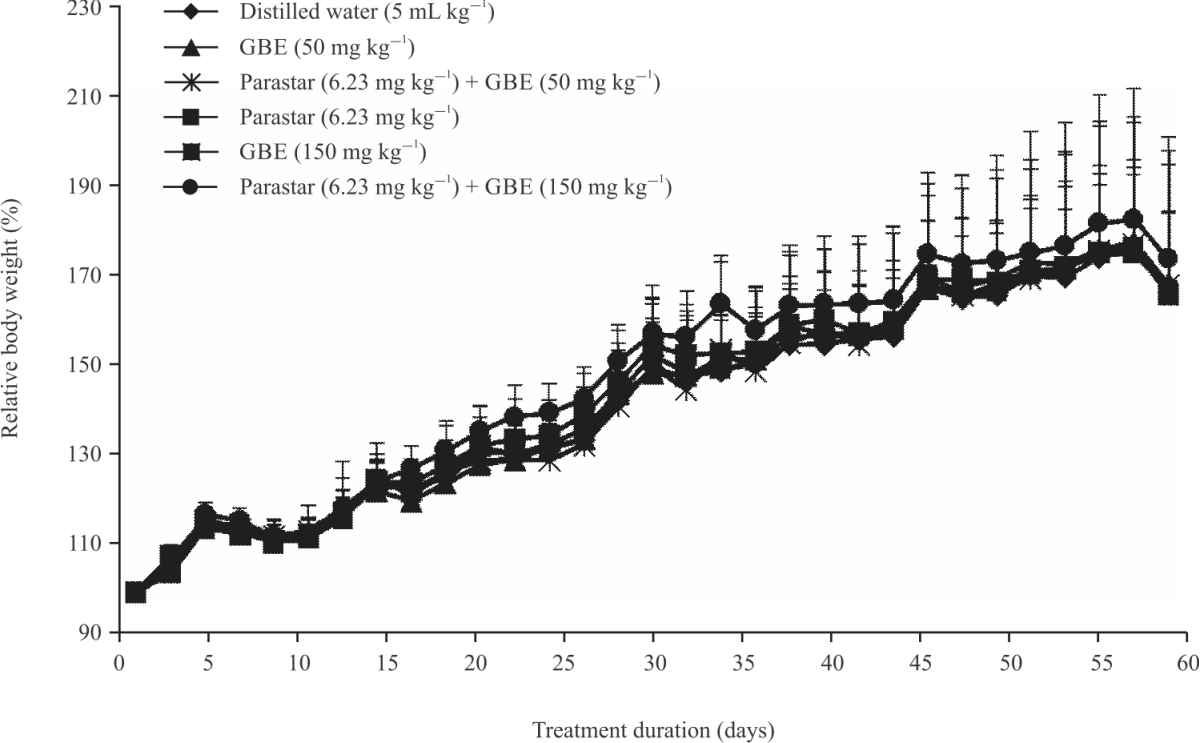 |
| Fig. 1: | Body weight variation of the rats during 60 days of treatment |
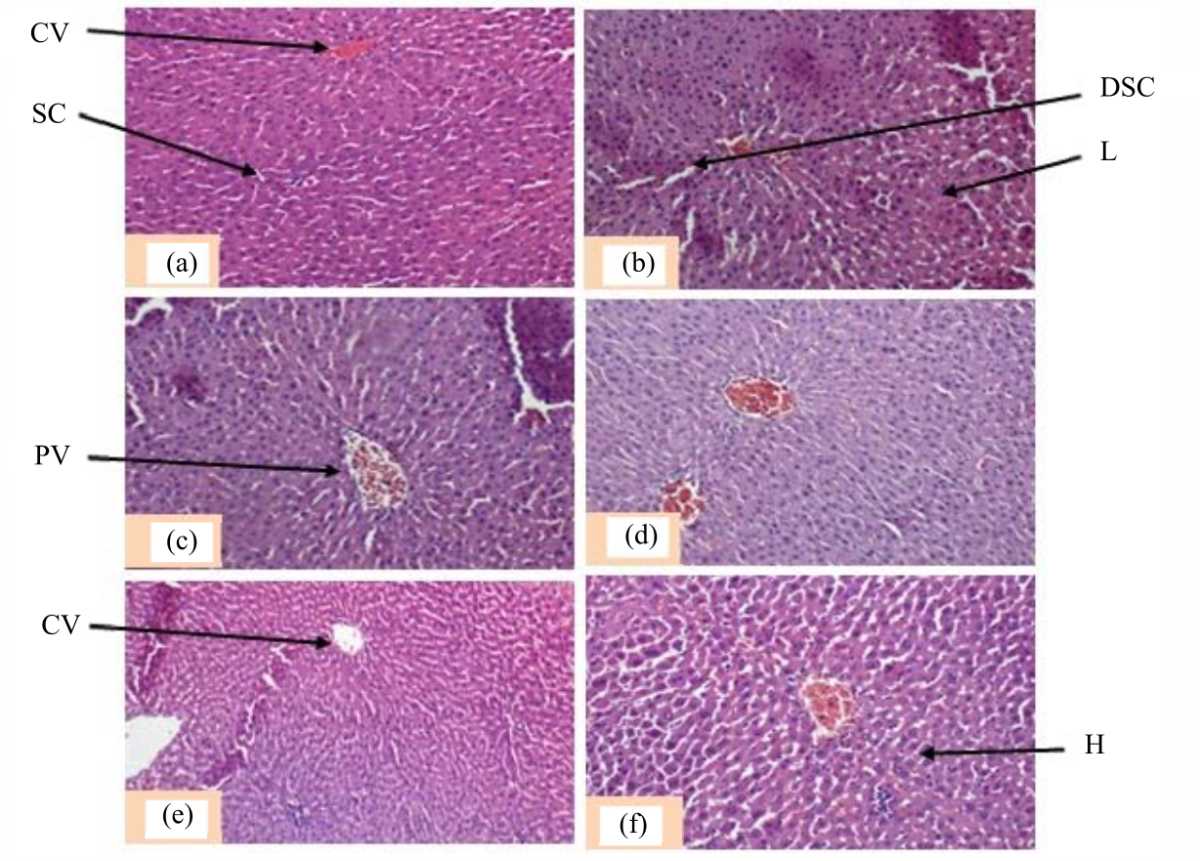 |
| Fig. 2(a-f): | Photomicrographs of the transverse section of rat livers, (a) Animals treated with distilled water (5 mL kg–1), (b) Parastar (6.23 mg kg–1), (c,d) GBE (50, 150 mg kg–1) and (e, f) Parastar (6.23 mg kg–1)+GBE (50, 150 mg kg–1) Magnification 400×, Staining: Hematoxylin/eosin, CV: Centrilobular vein, DSC: Dilated sinusoidal capillaries, H: Hepatocytes, L: Leucocytes, PV: Portal vein and SC: Sinusoidal capillaries |
| Table 1: | Relative liver and reproductive organ weights of different rat groups | |||||
| Negative control | Parastar treated groups | G. boninense only treated groups | ||||
| Distilled water | Parastar | Parastar+GBE | Parastar+GBE | GBE | GBE | |
| Treatments | (5 mL kg–1) | (6.23 mg kg–1) | (50 mg kg–1) | (150 mg kg–1) | (50 mg kg–1) | (150 mg kg–1) |
| Liver (g/100 g b.wt.) | 3.07±0.27a | 1.47±0.24b | 2.97±0.28a | 2.907±0.17a | 3.08±0.18a | 2.99±0.20a |
| Testes (g/100 g b.wt.) | 1.41±0.19a | 1.38±0.17a | 1.25±0.23a | 1.23±0.11a | 1.31±0.18a | 1.36±0.22a |
| Seminal vesicle (g/100 g b.wt.) | 0.30±0.05a | 0.36±0.04a | 0.51±0.06b | 0.35±0.04a | 0.40±0.08a | 0.36±0.06a |
| Epididymis (g/100 g b.wt.) | 0.58±0.08a | 0.54±0.07a | 0.85±0.11b | 0.74±0.12b | 0.53±0.10a | 0.52±0.06a |
| Prostate (g/100 g b.wt.) | 0.14±0.04a | 0.12±0.01a | 0.16±0.07a | 0.14±0.02a | 0.10±0.02a | 0.13±0.03a |
| Values in the table represent the Mean±SD of 8 rats per group and different letters mean significant difference (p<0.05) among treatments (Student-Newman-Keuls test or Games-Howell’s test when appropriate) | ||||||
Parastar significantly decreased (p<0.05) serum testosterone and vesicular fructose levels in animals when compared to the distilled water control group. In the presence of the insecticide, both doses of GBE normalized serum testosterone and vesicular fructose levels in animals when compared to the group given distilled water. Given alone, the mushroom extract (50, 150 mg kg–1) did not affect the serum testosterone and vesicular fructose levels as compared to the distilled water treated group.
In male rats, Parastar significantly decreased (p<0.05) sperm density and motility when compared to the vehicle control group. Co-administration experiments showed a preventive effect (p<0.05) of either 50 mg kg–1 or 150 mg kg–1 GBE on Parastar-induced decrease of sperm density and motility. As compared to the distilled water treated group, only the dose 50 mg kg–1 increased (p<0.05) sperm density while both doses increased (p<0.05) sperm motility.
Liver and testicular biochemical oxidative stress markers: In the liver and testes, Parastar significantly decreased (p<0.05) catalase activity and cellular glutathione levels, while this insecticide seemed not to have a significant effect on antioxidant biomarkers in the testes when compared to the group given distilled water (Table 4). The Parastar related alteration on oxidative stress biomarkers in the liver were significantly (p<0.05) prevented upon co-administration with any of the two doses of GBE to the animals (Table 4). Given alone, the mushroom extract did not affect the oxidative stress parameters either in the liver or testis.
Histopathological changes in liver and testis:The histological structure of the liver of rats that received distilled water (Fig. 2a) appears normal, the stained sections showed a centrilobular vein (CV), well visible hepatocytes (H) separated between them by sinusoid capillaries (SC). The histological sections of the liver of the rat given Parastar only showed dilation of sinusoid capillaries (DSC) and leucocytes (L) infiltration (Fig. 2b).
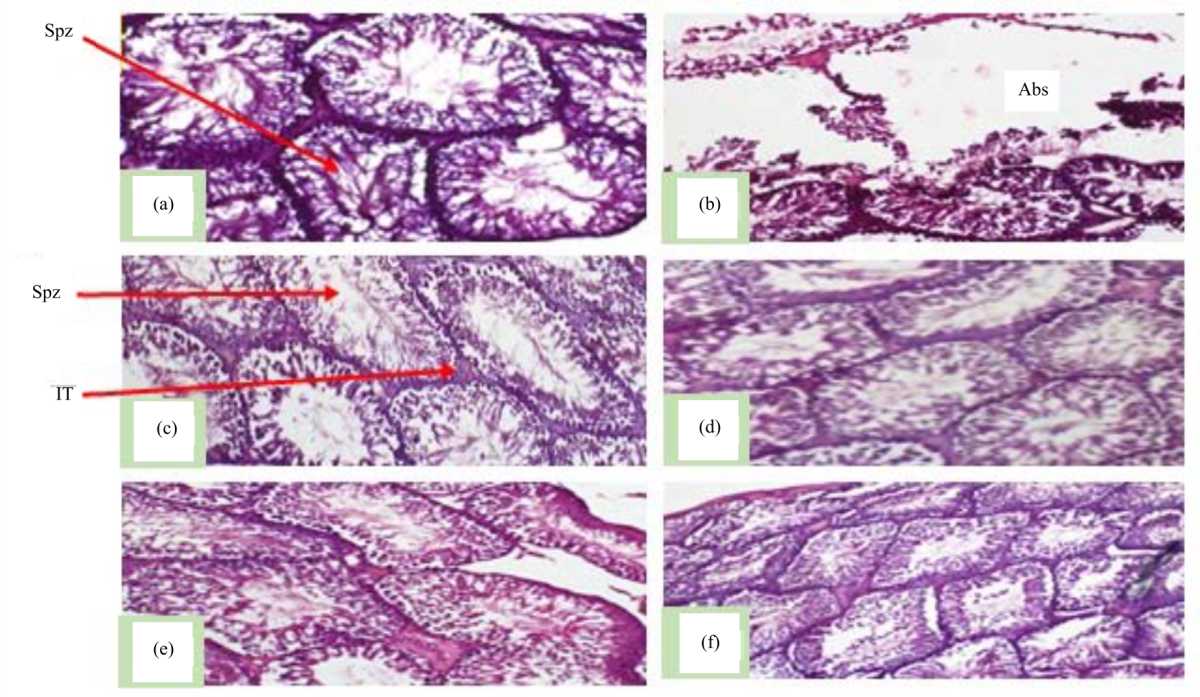 |
| Fig. 3(a-f): | Photomicrographs of the transversal section of the testes of rats, (a) Animals treated with distilled water (5 mL kg–1), (b) Parastar (6.23 mg kg–1), (c,d) GBE (50, 150 mg kg–1) and (e,f) Parastar (6.23 mg kg–1)+GBE (50, 150 mg kg–1) Magnification 400×, Staining: Hematoxylin/eosin, Abs: Absence of sperms, Spz: Spermatozoa and IT: Interstitial tissue |
| Table 2: | Serum biomarkers of liver cellular toxicity of different rat groups | |||||
| Negative control | Parastar treated groups | G. boninense only treated groups | ||||
| Distilled water | Parastar | Parastar+GBE | Parastar+GBE | GBE | GBE | |
| Treatments | (5 mL kg–1) | (6.23 mg kg–1) | (50 mg kg–1) | (150 mg kg–1) | (50 mg kg–1) | (150 mg kg–1) |
| Serum albumin (g dL–1) | 2.96±0.42a | 1.85±0.06b | 3.45±0.28ab | 3.01±0.23a | 3.08±0.35a | 3.03±0.60a |
| Serum cholesterol (mg dL–1) | 115.68±15.66a | 140.50±45.11a | 78.73±4.77c | 78.90±7.15c | 86.21±9.08a | 81.88±5.25b |
| Serum ALT (U L–1) | 3.15±0.88a | 4.86±0.60b | 4.31±0.52b | 3.91±0.28a | 3.91± 0.80a | 3.69±0.87a |
| Serum AST (U L–1) | 23.40±3.91a | 22.00±5.29a | 33.25±10.92c | 19.50±4.212a | 25.00±7.39a | 17.20±4.21a |
| Values in the table represent the Mean±SD of 8 rats per group, values not sharing the same superscript letters (a-b) differ significantly at p< 0.05 among treatments (Student-Newman-Keuls test), ALT: Alanine aminotransferase and AST: Aspartate aminotransferase | ||||||
| Table 3: | Serum biomarkers of liver function, serum testosterone levels, epididymal sperm characteristics and vesicular fructose concentration | |||||
| Negative control | Parastar treated groups | G. boninense only treated groups | ||||
| Distilled water | Parastar | Parastar+GBE | Parastar+GBE | GBE | GBE | |
| Treatments | (5 mL kg–1) | (6.23 mg kg–1) | (50 mg kg–1) | (150 mg kg–1) | (50 mg kg–1) | (150 mg kg–1) |
| Serum testosterone (ng mL–1) | 1.93±0.06a | 1.10±0.00b | 1.93±0.05a | 1.93±0.06a | 1.95±0.10a | 1.95±0.05a |
| Vesicular fructose (mM) | 0.011±0.002a | 0.004±0.001b | 0.015±0.003a | 0.013±0.002a | 0.012±0.003a | 0.015±0.003a |
| Sperm density×106 g–1 of the epididymis | 1019.20±119.10a | 360.79±83.84b | 1254.32±152.31c | 1123.91±89.14ac | 1174.93±208.81c | 1025.33±163.47a |
| Sperm motility (%) | 28.82±5.44a | 18.73±3.68b | 41.27±4.23c | 41.50±3.53c | 41.50±3.53c | 53.70±2.07d |
| Values in the table represent the Mean±SD of 8 rats per group and values not sharing the same superscript letters (a-d) differ significantly at p<0.05 among treatments (Student-Newman-Keuls test) | ||||||
Photomicrographs of the liver of the rats treated with the plant extract (GBE) and the co-treatment with Parastar and the aqueous extract did not show a major change (Fig. 2c-f) as compared to those from the vehicle control group.
Figure 3 represents a transversal section of the animal testes. The picture shows a normal structure of testis of rats from the vehicle group (that received distilled water), with numerous spermatozoa (Spz) within seminiferous tubules (Fig. 3a). Parastar treated group (Fig. 3b) revealed oedema in the interstitial tissues (IT), irregular arrangement and detachment of spermatogenic cells, severe loss of seminiferous tubules and Leydig cells, sloughing of germ cells and reduced hematoxylin/eosin staining intensity.
| Table 4: | Liver and testicular proteins and biochemical parameters of oxidative stress of different rat groups | |||||
| Negative control | Parastar treated groups | G. boninense only treated groups | ||||
| Distilled water | Parastar | Parastar+GBE | Parastar+GBE | GBE | GBE | |
| Treatments | (5 mL kg–1) | (6.23 mg kg–1) | (50 mg kg–1) | (150 mg kg–1) | (50 mg kg–1) | (150 mg kg–1) |
| Liver parameter | ||||||
| GSH (mmole/mg of proteins) | 1.73±0.03a | 1.61±0.01b | 1.75±0.07a | 1.79±0.03a | 1.74±0.05a | 1.76±0.05a |
| TBARS (nmole/mg of proteins) | 0.29±0.01a | 0.32±0.01a | 0.30±0.02a | 0.29±0.01a | 0.29±0.01a | 0.29±0.01a |
| SOD (IU/mg of proteins) | 23.40±3.91a | 22.00±5.29a | 33.25±10.92c | 19.50±4.212a | 25.00±7.39a | 17.20±4.21a |
| Catalase activity (IU/mg of proteins) | 39.86±5.78a | 34.79±2.08b | 36.86±2.24a | 37.22±2.20a | 36.81±2.31a | 37.95±3.86a |
| Testicular parameters | ||||||
| GSH (mmole/mg of proteins) | 1.03±0.07a | 0.96±0.031a | 0.88±0.04a | 1.03±0.09a | 1.01±0.07a | 1.05±0.09a |
| TBARS (nmole/mg of proteins) | 0.17±0.02a | 0.20±0.02a | 0.22±0.04a | 0.21±0.04a | 0.17±0.02a | 0.17±0.03a |
| SOD (IU/mg of proteins) | 5.79±0.94a | 5.10±0.79a | 5.33±0.51a | 6.86±0.71ab | 5.83±0.54a | 6.83±0.76ab |
| Catalase activity (IU/mg of proteins) | 119.84±8.08a | 109.63±20.42a | 122.24±10.98a | 127.24±10.40ab | 127.09±11.78ab | 128.24±14.37ab |
| Values in the table represent the Mean±SD of 8 rats per group, values not sharing the same superscript letters (a-b) differ significantly at p<0.05 among treatments (Student-Newman-Keuls test), GSH: Reduced glutathione, SOD: Superoxide dismutase and TBARS: Thiobarbituric acid reactive substances | ||||||
There was no major modification on the ultrastructure of the testes of rats treated, respectively with GBE (50, 150 mg kg–1), Parastar+GBE (50, 150 mg kg–1) (Fig. 3c-f) as compared to those from the vehicle control group.
DISCUSSION
Parastar induced reproductive toxicity through decreased serum testosterone levels, vesicular fructose concentrations, sperm density and motility of treated rats. Testosterone is important for the development and maintenance of secondary sexual characteristics as well as the maintenance of spermatogenesis25. The decrease in testosterone levels is consistent with findings of Nantia et al.13 who observed a dose-dependent decrease in serum testosterone levels, especially at the highest dose of Parastar (6.23 mg kg–1) after 35 days of rat treatment. The decrease in testosterone levels could be due to alteration of the gonadotropin, Luteinizing Hormone (LH), which is responsible for the production of testosterone26. Fructose is an important energy source for spermatozoa and hence a higher probability of their survival and fertilization power. Fructose secretion by the seminal vesicle is androgen-dependent27. Depletion of vesicular fructose could result from the decrease in testosterone secretion caused by pesticide treatment. This hampers the glycolytic metabolism of spermatozoa resulting in low energy for spermatozoa survival, abnormal sperm functions that could result in infertility28. A consistent study was conducted by Desai et al.29 who observed a significant reduction in vesicular fructose in Swiss Albino mice treated for 45 days with 3 and 6 mg kg–1 b.wt., of deltamethrin, a synthetic pyrethroid insecticide. A direct association exists between serum testosterone and sperm motility/fertility29. Decrease in sperm density and motility may be related at least partly to decreased testosterone levels following the pesticide treatment. Such observations were made by Nantia et al.13 who revealed a decrease in sperm motility and density of male rats exposed to Parastar. One of the considered mechanisms of pesticide toxicity on male reproductive function is through oxidative stress30. The testes contain antioxidant systems that maintain the balance between reactive oxygen species (ROS) and antioxidants including GSH and enzymes preventing oxidative stress (OS). Glutathione (GSH), a ubiquitous tripeptide, γ-Glu-Cys-Gly, being the first line of defence against the deleterious effects of ROS, is an important antioxidant in most mammalian cells and it contains the thiol moiety which is a potent reducing agent31. GSH neutralizes superoxide anions and hydroxyl radicals by giving its electrons. Superoxide dismutase (SOD) and Catalase (CAT) enzymes work in synergy sequentially to neutralize free radicals. SOD catalyses the dismutation of superoxide anion (O–2) to hydrogen peroxide (H2O2) and oxygen (O2). CAT together with GSH peroxidase degrades H2O2 to water (H2O) and oxygen (O2). Treatment with Parastar decreased GSH levels and the activities of CAT and SOD while increasing the levels of Thiobarbituric Acid Reactive Substances (TBARS). Such observations were made by Nantia et al.13 in the testicular homogenates. Parastar may promote lipid peroxidation by altering the physiological bilayer structure, modifying membrane properties such as permeability, resulting in an increase in TBARS in the testes. Moreover, the antioxidant defence system may be overwhelmed by Parastar leading to OS as a result of the generation of free radicals and active intermediates in the testes exceeding the ability of antioxidants (including, GSH, SOD and CAT) to neutralize and inactivate them (free radicals). The testicular function was further assessed through histological analysis of the organ. Parastar led to testicular damages characterized by moderate to severe seminiferous tubule degeneration, sloughing, atrophy and degeneration of the germ cells, oedema and Leydig cell tumour, arrest of spermatogenesis and spermiogenesis and reduction in the hematoxylin/eosin staining intensity. This is similar to the findings of Najafi et al.32 who observed adverse histological changes in testicular tissue and sperm mortality. Parastar negatively affected the structure and function of Leydig cells, which is associated with androgen, testosterone. This is in line with a previous study according to which pyrethroids and their metabolites disrupt the function of multiple nuclear hormone receptors33. Thus, exposure to Parastar is a major issue as it hampers the male reproductive function13.
The liver is not only a major effector of endobiotic and xenobiotic metabolism but also a major non-reproductive target organ of sex steroids action. Therefore, its function affects the reproductive axis especially adaptation of energy metabolism to sex-specific reproductive needs10. Also, liver weight is an essential parameter in toxicological studies. Liver weights were increased in rats treated with Parastar. This increase could be due to Parastar interfering with metabolism. This result is consistent with an earlier study of Mehmood et al.34 with imidacloprid, one of the active ingredients in Parastar. Liver function was evaluated by assessing serum albumin and cholesterol concentrations and the activities of transaminases. Parastar decreased serum albumin and significantly increased serum cholesterol levels and activity of Alanine Transaminase (ALT). A decrease in albumin could be due to Parastar interfering with amino acid metabolism which is associated with liver disease35. An increase in cholesterol could be due to the ability of Parastar to interfere with lipid metabolism and utilization. This result corroborates with the observations of Mehmood et al.34 following exposure of male rats to imidacloprid. ALT is confined in the cytosol of hepatocytes and a more sensitive marker of liver injury. The increase in the activity of ALT in the Parastar group suggests degeneration or necrosis of hepatocytes which causes an increase in cell membrane permeability and leads to increased transaminases in blood. Like the testes, the liver contains important antioxidant systems that maintain the balance between ROS and antioxidants preventing oxidative damage. Parastar decreased GSH levels and activities of CAT and SOD while increasing the levels of TBARS. This result is consistent with the previous study of Nantia et al.13 who reported the potential of Parastar to induce oxidative stress in male rats. In correlation with the biochemical alterations, histological alterations in the liver cellular structure of Parastar-exposed rats revealed dilation of sinusoidal capillaries and leukocyte infiltration in liver tissues. This result corroborates with the observations of Djahra et al.36 following exposure of albinos male Wistar rats to lambda-cyhalothrin, an ingredient in Parastar formulation. Therefore, Parastar is a toxic insecticide capable of altering hepatic function as well as the aforementioned reproductive parameter, an alarming issue. Considering use of this insecticide by farmers of the Santa subdivision of the Northwest region of Cameroon and elsewhere37, there is a need for a proper remedy of this situation to preserve human health.
As an option towards solving the toxicity posed by Parastar, we co-administered the insecticide with both doses of the aqueous extract of G. boninense, a mushroom that exhibits safe38 and antioxidant properties17,39 and some health benefits20. Co-administration of Parastar with both doses of the mushroom extract normalized the reducing effect of Parastar on serum testosterone and vesicular fructose levels. Co-treatment of male rats also restored sperm density and motility to normalcy. Hence, GBE could possess counteracting effects on the inhibitory properties of Parastar on Luteinizing Hormone (LH) secretion by the anterior pituitary gland, responsible for the production of testosterone that maintains the process of spermatogenesis29. Moreover, the mushroom extract normalized testicular levels of TBARS and GSH, activities of SOD and CAT against Parastar toxicity. This suggests that the extract is rich in non-enzymatic secondary metabolites that prevent oxidative stress (OS) thanks to its undeniable beneficial earlier reported antioxidant effects17. Moreover, the concomitant treatment of 150 mg kg–1 GBE with Parastar revealed significant higher activities of SOD and CAT when compared to the Parastar group, thus partly supporting the synergistic effect of these enzymes and the effectiveness of the higher dose of GBE. This agrees with earlier studies that revealed antioxidant contents of the members of Ganodermataceae40-42 and in particular, G. boninense17,39. As a consequence, the extract proved to improve the antioxidant status, contributing to the elimination of ROS thus alleviating Parastar-induced reproductive toxicity. Moreover, impaired sperm characteristics, testosterone levels and liver damages by Parastar corroborated with the histology of the testes. Rats administered with both doses of GBE revealed no marked pathological changes. Upon co-administration, both doses of the mushroom extract alleviated the toxic effect of the pesticide on interstitial cells containing the continuous arrangement of Leydig cells. In fact, upon co-administration, results revealed the normal intensity of the hematoxylin/eosin stains, supporting the beneficial effects of the mushroom extract that reversed the toxic effect of the pesticide that caused a reduction in stain intensity (the cell nuclei normally take up the haematoxylin/eosin stains), thus revealing the pharmacological potentials of GBE in enhancing male reproductive function.
The co-administration of Parastar with GBE increased the liver weights which suggest the induction and presence of some xenobiotic enzymes in G. boninense which could lead to an increase in protein synthesis. Co-administration of the mushroom extract and Parastar significantly increased serum albumin levels alleviating the decrease by Parastar. Improvement could be due to natural enzymes or metabolites in G. boninense that induced the anabolism of amino acids. Co-administration not only mitigated but alleviated the negative effects of Parastar with a significant decrease in serum cholesterol concentration. Early reports support the cholesterol-lowering capacity of patented products derived from other species of the Ganodermataceae family which may be thanks to the presence of lipid-lowering ingredients such as beta-glucans in Ganoderma genus interfering with the absorption of cholesterol into the blood43. Notwithstanding, co-treatment of 150 mg kg–1 GBE and Parastar prevented the high ALT activity induced by the insecticide while both doses of GBE significantly decreased AST activity as opposed to Parastar. This is consistent with the study of Djahran et al.36 who reported similar results with lambda-cyhalothrin, one active ingredient of Parastar, that observed elevated serum AST and ALT activities which were alleviated after treatment with Coriandrum sativum aqueous extract. The results of concomitant administration on the activities of ALT and AST suggest that GBE could contain metabolites that interact with the liver plasma membrane and reduce their permeability as opposed to Parastar. Moreover, the extract normalized the levels of liver TBARS and GSH, activities of SOD and CAT against Parastar toxicity. Studies have demonstrated that members of the Ganodermataceae can reduce oxidative damage by directly scavenging free radicals generated in the cell and reducing membrane peroxidation40-42. Based on liver histological examination, co-administration of Parastar and both doses of GBE alleviated the severity of hepatic injury induced by Parastar. This suggests the mushroom’s richness in bioactive compounds, especially triterpenoids and polysaccharides, which play critical roles in a variety of biological effects earlier reported44,45. These results support the hepato-protective effects of the mushroom as reported by other natural products such as medicinal plants46,47.
Reproductive toxicity of xenobiotics including pesticides is not limited to its direct effects on the reproductive system. It can also result from biochemical reactions of xenobiotic metabolism by the liver with negative consequences on the endocrine-function10,48. The liver and the reproductive system work synergistically affecting each other in a multifaceted bidirectional fashion. Not surprisingly, earlier reports have described the association between energy homeostasis in the liver and male gonadal function with liver diseases associated with reproductive dysfunction. Consequently, reproductive alteration is largely restored or improved by successful liver transplantation10,49,50. The mushroom extract may act through factors implicated in male reproductive function regulation such as the stimulation of liver and testicular defence systems, preventing Parastar-induced hepatotoxicity and reproductive toxicity.
CONCLUSION
The findings of this study showed that Parastar altered liver and reproductive functions evidenced through decreased relative liver weights, increased activities of aminotransferases (alanine aminotransferase and aspartate aminotransferase) and decreased epididymal sperm density and motility, serum testosterone and vesicular fructose levels. Moreover, this insecticide increased lipid peroxidation, decreased glutathione levels, superoxide dismutase and catalase activities and induced pathological changes in the testis and liver. Interestingly, the aqueous extract of G. boninense (GBE) efficiently protects the liver and reproductive parameters against Parastar-induced toxicity. This study provides evidence that G. boninense is a promising mushroom in protecting the liver and reproductive functions against pesticide-induced toxicity. However, further studies should be conducted to show the mechanism of action and active ingredients in the mushroom extract with pharmacological effects.
SIGNIFICANCE STATEMENT
This study highlights the beneficial effects of the aqueous extract from the mushroom G. boninense, in protecting male reproductive and liver functions against the insecticide formulation Parastar. This beneficial effect of G. boninense was at least in part attributed to the antioxidant and androgenic activity of active substances from the mushroom. The mushroom has been used as a foodstuff and may be easily accepted by humans as a potential protective agent against pesticide residues, though this requires further confirmation in humans.
ACKNOWLEDGMENTS
The authors would like to thank Prof. Oumar Mahamat and Mrs. Ayinuih Sahfua Asamaou for their technical assistance in conducting the experiments.
REFERENCES
- Bao, L.J., Y.L. Wei, Y. Yao, Q.Q. Ruan and E.Y. Zeng, 2015. Global trends of research on emerging contaminants in the environment and humans: A literature assimilation. Environ. Sci. Pollut. Res., 22: 1635-1643.
CrossRefDirect Link - Prasanthi, K., Muralidhara and P.S. Rajini, 2005. Fenvalerate-induced oxidative damage in rat tissues and its attenuation by dietary sesame oil. Food Chem. Toxicol., 43: 299-306.
CrossRefDirect Link - Sonchieu, J., M.B. Ngassoum, A.E. Nantia and P.S. Laxman, 2017. Pesticide applications on some vegetables cultivated and health implications in Santa, North West-Cameroon. SSRG Int. J. Agric. Environ. Sci., 4: 39-46.
Direct Link - Sanda, A.K., A.E. Nantia, T.F.P. Manfo, R.T. Toboh and R.E. Abende et al., 2022. Subchronic administration of parastar insecticide induced behavioral changes and impaired motor coordination in male Wistar rats. Drug Chem. Toxicol., 45: 426-434.
CrossRefDirect Link - Manfo, F.P.T., P.F. Moundipa, H. Dechaud, A.N. Tchana, E.A. Nantia, M.T. Zabot and M. Pugeat, 2012. Effect of agropesticides use on male reproductive function: A study on farmers in Djutitsa (Cameroon). Environ. Toxicol., 27: 423-432.
CrossRefDirect Link - Jamal, F., Q.S. Haque, S. Singh and M. Arshad, 2016. The influence of pesticides on hepatic and renal functions in occupational sprayers of rural Malihabad, Lucknow (India). Toxicol.: Open Access, Vol. 2.
CrossRefDirect Link - Monserrat, J.M., P.E. Martinez, L.A. Geracitano, L.L. Amado and C.M. Martins, 2007. Pollution biomarkers in estuarine animals: Critical review and new perspectives. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol., 146: 221-234.
CrossRefDirect Link - Vona, R., L. Pallotta, M. Cappelletti, C. Severi and P. Matarrese, 2021. The impact of oxidative stress in human pathology: Focus on gastrointestinal disorders. Antioxidants, Vol. 10.
CrossRefDirect Link - Mossa, A.T.H., T.M. Heikal, M. Belaiba, E.G. Raoelison, H. Ferhout and J. Bouajila, 2015. Antioxidant activity and hepatoprotective potential of Cedrelopsis grevei on cypermethrin induced oxidative stress and liver damage in male mice. BMC Complement. Altern. Med., Vol. 15.
CrossRefDirect Link - Grossmann, M., M.E. Wierman, P. Angus and D.J. Handelsman, 2019. Reproductive endocrinology of nonalcoholic fatty liver disease. Endocr. Rev., 40: 417-446.
CrossRefDirect Link - Ratnasooriya, W.D., S.S. Ratnayake and Y.N. Jayatunga, 2002. Effects of pyrethroid insecticide ICON (lambda cyhalothrin) on reproductive competence of male rats. Asian J. Androl., 4: 35-41.
PubMedDirect Link - Baligar, P.N. and B.B. Kaliwal, 2001. Induction of gonadal toxicity to female rats after chronic exposure to mancozeb. Ind. Health, 39: 235-243.
CrossRefDirect Link - Nantia, E.A., A.S. Kada, F.P.T. Manfo, N.N. Tangu, K.M. Mbifung, D.H. Mbouobda and A. Kenfack, 2018. Parastar insecticide induced changes in reproductive parameters and testicular oxidative stress biomarkers in Wistar male rats. Toxicol. Ind. Health, 34: 499-506.
CrossRefDirect Link - Yang, J.H., H.C. Lin and J.L. Mau, 2002. Antioxidant properties of several commercial mushrooms. Food Chem., 77: 229-235.
CrossRefDirect Link - Barros, L., P. Baptista and I.C.F.R. Ferreira, 2007. Effect of Lactarius piperatus fruiting body maturity stage on antioxidant activity measured by several biochemical assays. Food Chem. Toxicol., 45: 1731-1737.
CrossRefPubMedDirect Link - Hapuarachchi, K.K., W.A. Elkhateeb, S.C. Karunarathna, C.R. Cheng and A.R. Bandara et al., 2018. Current status of global Ganoderma cultivation, products, industry and market. Mycosphere, 9: 1025-1052.
CrossRefDirect Link - Mohammed, A., A.B. Adelaiye, M.S. Abubakar and E.M. Abdurahman, 2007. Effects of aqueous extract of Ganoderma lucidum on blood glucose levels of normoglycemic and alloxan-induced diabetic wistar rats. J. Med. Plants Res., 1: 34-37.
Direct Link - Perrard, M.H., N. Sereni, C. Schluth-Bolard, A. Blondet and S.G. d'Estaing et al., 2016. Complete human and eat ex vivo spermatogenesis from fresh or frozen testicular tissue. Biol. Reprod., 95: 1-10.
CrossRefDirect Link - Jin, C.B., 2005. Ganoderma boninense: Fungus with blood sugar lowering property. J. Trop. Med. Plants, 6: 159-163.
Direct Link - Ngoula, F., P. Watcho, M.C. Dongmo, A. Kenfack, P. Kamtchouing and J. Tchoumboue, 2007. Effects of pirimiphos-methyl (an organophosphate insecticide) on the fertility of adult male rats. Afr. Health Sci., 7: 3-9.
PubMedDirect Link - Sanda, A.K., E. Aneng, M. Mbouh, F.P.T. Manfo, S.N. Ngimgoh and E.A. Nantia, 2021. Testicular antioxidants and testosterone enhancing effects of the hydro-ethanolic extract of Rauvolfia vomitoria (Apocynacaea) in male Wistar rats. Invest. Med. Chem. Pharmacol., Vol. 4.
CrossRefDirect Link - Boufack J.L., N.A.J. Guiekep, A.E. Nantia, B.N.Vemo and A. Kenfack et al., 2021. Reproductive toxicity of paraquat in male guinea pig (Cavia porcellus). Asian J. Anim. Vet. Adv., 7: 15-24.
Direct Link - Sanda, A.K., D. Fotsing, O. Mahamat, V. Natesan and J. Rajendran et al., 2021. Effect of Allanblackia floribunda stem bark aqueous extract in male albino wistar rats: Androgenic activities and penile nitric oxide (NO) production. Asian J. Pharm. Pharmacol., 7: 63-73.
CrossRefDirect Link - Motofei, I.G. and D.L Rowland, 2005. The physiological basis of human sexual arousal: Neuroendocrine sexual asymmetry. Int. J. Androl., 28: 78-87.
CrossRefDirect Link - Harris, I.D., C. Fronczak, L. Roth and R.B. Meacham, 2011. Fertility and the aging male. Rev. Urol., 13: 184-190.
Direct Link - Al-Daghistani, H.I., A.W.R. Hamad, M. Abdel-Dayem, M. Al-Swaifi and M.A. Zaid, 2010. Evaluation of serum testosterone, progesterone, seminal antisperm antibody, and fructose levels among Jordanian males with a history of infertility. Biochem. Res. Int., Vol. 2010.
CrossRefDirect Link - Guyton, A.C. and J.E. Hall, 2006. Textbook of Medical Physiology. 11th Edn., Elsevier Saunder, Philadelphia, ISBN: 9788481749267, Pages: 1116.
Direct Link - Desai, K.R., N. Moid, P.B. Patel and H.N. Highland, 2016. Evaluation of deltamethrin induced reproductive toxicity in male swiss albino mice. Asian Pac. J. Reprod., 5: 24-30.
CrossRefDirect Link - Manfo, F.P.T., E.A. Nantia and P.P. Mathur, 2014. Effect of environmental contaminants on mammalian testis. Curr. Mol. Pharm., 7: 119-135.
Direct Link - Apel, K. and H. Hirt, 2004. Reactive oxygen species: Metabolism, oxidative stress and signal transduction. Annu. Rev. Plant Biol., 55: 373-399.
CrossRefPubMedDirect Link - Najafi, G., M. Razi, A. Hoshyar, S. Shahmohamadloo and S. Fayzi, 2010. The effect of chronic exposure with imidacloprid insecticide on fertility in mature male rats. Int. J. Fertil. Steril., 4: 9-16.
Direct Link - Du, G., O. Shen, H. Sun, J. Fei and C. Lu et al., 2010. Assessing hormone receptor activities of pyrethroid insecticides and their metabolites in reporter gene assays. Toxicol. Sci., 116: 58-66.
CrossRefDirect Link - Mehmood, T., M. Saeed, M.M. Ahmad, M. Sikram, F. Siddique and Q. Tabassam, 2017. Effect of imidacloprid (insecticide) on serum biochemical parameters and degenerative lesions in male rat's liver. Sindh Univ. Res. J. (Sci. Ser.), 49: 605-612.
Direct Link - Nyblom, H., U. Berggren, J. Balldin and R. Olsson, 2004. High AST/ALT ratio may indicate advanced alcoholic liver disease rather than heavy drinking. Alcohol Alcoholism, 39: 336-339.
CrossRefPubMedDirect Link - Djahra, A.B., M. Benkaddour, S. Benkherara and B. Ouahiba, 2020. Antioxidant and hepatoprotective potential of Coriandrum sativum L. against hepatic injury by lambda-cyhalothrin insecticide. J. Drug Delivery Ther., 10: 182-188.
CrossRefDirect Link - Pouokam, G.B., W.L. Album, A.S. Ndikontar and M.E.H. Sidatt, 2017. A pilot study in Cameroon to understand safe uses of pesticides in agriculture, risk factors for farmers' exposure and management of accidental cases. Toxics, Vol. 5.
CrossRefDirect Link - Sasidharan, S., O. Jinxuan and L.Y. Latha, 2010. Toxicity of Ganoderma boninense methanol extract in mice. Mycology, 1: 85-91.
CrossRefDirect Link - Rawat, A., M. Mohsin, P.S. Negi, A.N. Sah and S. Singh, 2013. Evaluation of polyphenolic contents and antioxidant activity of wildly collected Ganoderma lucidum from central Himalayan hills of India. Asian J. Plant Sci. Res., 3: 85-90.
Direct Link - Rajasekaran, M. and C. Kalaimagal, 2011. In vitro antioxidant activity of ethanolic extract of a medicinal mushroom, Ganoderma lucidum. J. Pharm. Sci. Res., 3: 1427-1433.
Direct Link - Celık, G.Y., D. Onbaslı, B. Altınsoy and H. Allı, 2014. In vitro antimicrobial and antioxidant properties of Ganoderma lucidum extracts grown in Turkey. Eur. J. Medic. Plants, 4: 709-722.
CrossRefDirect Link - Obodai, M., D.L.N. Mensah, A. Fernandes, N.K. Kortei and M. Dzomeku et al., 2017. Chemical characterization and antioxidant potential of wild Ganoderma species from Ghana. Molecules, Vol. 22.
CrossRefDirect Link - Berger, A., D. Rein, E. Kratky, I. Monnard and H. Hajjaj et al., 2004. Cholesterol-lowering properties of Ganoderma lucidum in vitro, ex vivo, and in hamsters and minipigs. Lipids Health Dis., Vol. 3.
CrossRefDirect Link - Isaka, M., P. Chinthanom, S. Kongthong, K. Srichomthong and R. Choeyklin, 2013. Lanostane triterpenes from cultures of the basidiomycete Ganoderma orbiforme BCC 22324. Phytochemistry, 87: 133-139.
CrossRefDirect Link - Baby, S., A.J. Johnson and B. Govindan, 2015. Secondary metabolites from Ganoderma. Phytochemistry, 114: 66-101.
CrossRefDirect Link - Ozturk, I.C., F. Ozturk, M. Gul, B. Ates and A. Cetin, 2009. Protective effects of ascorbic acid on hepatotoxicity and oxidative stress caused by carbon tetrachloride in the liver of Wistar rats. Cell Biochem. Funct., 27: 309-315.
CrossRefPubMedDirect Link - Zeghib, K. and D.A. Boutlelis, 2019. Protective role of aqueous extract of Atriplex halimus L. against benzene-induced damage on renal function and glomerular cells in rats. Asian J. Pharm. Clin. Res., 12: 387-392.
Direct Link - Schuppe, H.-C., P. Wieneke, S. Donat, E. Fritsche, F.-M. Köhn and J. Abel, 2000. Xenobiotic metabolism, genetic polymorphisms and male infertility. Andrologia, 32: 255-262.
CrossRefDirect Link - Rostyslav, V.B., M.V. Drahulian, P.V. Buchek, and T.P. Gulko, 2018. High regenerative capacity of the liver and irreversible injury of male reproductive system in carbon tetrachloride-induced liver fibrosis rat model. EPMA J., 9: 59-75.
CrossRefDirect Link - Samuel, V.T. and G.I. Shulman, 2018. Nonalcoholic fatty liver disease as a nexus of metabolic and hepatic diseases. Cell Metab., 27: 22-41.
CrossRefDirect Link