
Elham A. Abd-Allah
Department of Zoology, Faculty of Science, The New Valley University, 72511 El-Kharga, Egypt
LiveDNA: 20.29863
Ahlam A. Alahmadi
Department of Biological Sciences, Faculty of Science, King Abdulaziz University, Jeddah, Saudi Arabia
Hanan S.A. Waly
Department of Zoology, Faculty of Science, Assiut University, Assiut, Egypt
Bassam A. Alahmadi
Department of Biology, College of Science, Taibah University, Saudi Arabia
Asian Journal of Animal and Veterinary Advances
Year: 2020 | Volume: 15 | Issue: 1 | Page No.: 1-12







ABSTRACT
Background and Objective: Curcumin (CUR) is the main ingredient of turmeric, has been confirmed to possess anti-inflammatory and anti-oxidative characteristics. Liver diseases are a global health issue as the liver is the essential detoxifying organ. We conducted our study to verify the protective effects of CUR against carbon tetrachloride (CCL4), which stimulates liver and spleen injury in mice. Materials and Methods: Thirty adult male mice were randomly divided into 3 groups (8-10 mice/group). First group served as control group, second one intraperitoneally injected with CCL4 (0.1 mL kg–1, i.p.). The third group intraperitoneally injected with CCL4 (0.1 mL kg–1) and co-treated with curcumin (200 mg kg–1) intragastrically, the experiment continued for 8 weeks, 3 times/weekly. Results: The CCL4 increased serum liver enzymes (ALT, AST and ALP), mRNA expression levels of TNF-α, IL-1 and IL-6 were detected as pro-inflammatory markers and 5LO, NOX2 and COX-2 as inflammatory biomarkers for liver and spleen, significant elevations was detected in LPO levels. GSH, GST, GPx, CAT and SOD levels as antioxidants were significantly lessened. Histopathological alterations were also detected. Interestingly, curcumin supplementation of CCL4 treated group decreased expression of inflammatory markers as well as improved antioxidant status and histoarchitecture in liver and spleen. Conclusion: This data suggested that curcumin protect the liver and spleen from acute CCL4 induced injury in a rodent model by suppressing hepatic and splenic oxidative stress expression levels of inflammatory markers. Therefore, curcumin are potential therapeutic antioxidant agents against acute hepatotoxicity and spleenotoxicity.
PDF Abstract XML References Citation
How to cite this article
Elham A. Abd-Allah, Ahlam A. Alahmadi, Hanan S.A. Waly and Bassam A. Alahmadi, 2020. Anti-inflammatory Properties of Curcumin May Be Useful for Treating the Deleterious Effects of CCL4 on Liver and Spleen. Asian Journal of Animal and Veterinary Advances, 15: 1-12.
DOI: 10.3923/ajava.2020.1.12
URL: https://scialert.net/abstract/?doi=ajava.2020.1.12
DOI: 10.3923/ajava.2020.1.12
URL: https://scialert.net/abstract/?doi=ajava.2020.1.12
INTRODUCTION
Liver fibrosis is a process arises in response to varieties of liver injuries such as toxic or drugs exposure1, it is characterized by the excess deposition of the components of extracellular matrix which commonly ending with hepatic dysfunction, even hepatocellular carcinoma, leading to increased morbidity and mortality2. Toxic reagents are one of the utmost causes inducing liver damage and fibrosis that cannot be ignored3. The reasons of drug-induced liver injury are multifactorial, including toxic effects caused by reactive metabolites, reactive oxygen species ROSs, inflammatory reactions and imbalances between cellular damage and protective responses4. Every living cell also has many types of defense mechanisms to counterpoise the negative influence of ROSs, such as antioxidant enzymes: Catalase, superoxide dismutase or glutathione peroxidase, so the detoxification of ROS is a major requirement of aerobic life, which accomplished via several enzymatic and non-enzymatic antioxidant mechanisms that are available in different cell components5. The CCL4, usually used as a laboratory component remarkable by its harmfulness leading to liver lesion and liver fibrosis, has been extensively applied in liver-related studies6. It is notable that CCL4-initiated intense liver damage in a murine model which is a classical system for studying possible hepatoprotective factors, owing to the matching of its molecular mechanism with acute chemical liver and spleen injury in humans7.
A relationship between the liver and spleen has been suggested at least for three main reasons. Anatomically, the 2 organs are important portal circulation components. Histologically, the liver and spleen have a like reticuloendothelial structures, which constantly contribute in substance exchange and cellular movement8. Immunologically, both the liver and spleen play vital roles in immune homeostasis and clearance of pathogens. Therefore, the concept of a liver-spleen axis has been proposed as crossroads for immunity, pathogen clearance and metabolism in several conditions including chronic liver diseases9. Earlier studies have confirmed effects of innate and adaptive immune cells in progress of liver fibrosis or cirrhosis10. Nevertheless, direct evidence for the participation of splenic immune cells or spleen-derived factors has only recently arisen, proposing that splenic contributions to hepatic fibrogenesis, hepatic immune microenvironment dysregulation and the disturbance of liver recovery may be responsible11.
Unfortunately, artificial drugs used in the remediation of liver diseases are inadequate and sometimes can have dangerous side effects. This is why many people in the world over including those in developed countries turning complementary and alternative medicine. Many common medications use herbal medication for the treatment of liver diseases12. The CUR is an active naturally ingredient of turmeric rhizomes, has been shown to modulate oxidative stress and is considered to be a potent antioxidant13. The CUR has a polyphenolic structure and diketone functional groups, it is a stronger antioxidant inhibitor of lipid peroxidation than other flavonoids, which have a single phenolic hydroxyl group14. The CUR represents a class of anti-inflammatory and antioxidants reported to be a scavenger of the formed ROS15.
So, the objective of this study is to provide information concerning CUR as an antioxidant that has demonstrated utility in improving liver and spleen diseases via the regulation of COX-2, NOX2, 5-LO and pro-inflammatory cytokines (IL-1, IL-6 and TNF-α) beside its effects on enzymatic and non-enzymatic antioxidant defense mechanisms, on mice utilizing CCL4 as a model hepatotoxic and splenotoxic agent that allowed for the evaluation of necrosis, inflammation and fibrosis.
MATERIALS AND METHODS
Chemicals and reagent kits: Carbon tetrachloride, curcumin, thiobarbituric acid, GSH, superoxide dismutase, epinephrine and 5,5’-dithio-bis-(2-nitrobenzoic acid; DTNB) were purchased from Sigma-Aldrich (St. Louis, MO, USA). Activities of GSH-Px, GSH-ST, glucose-6-phosphate dehydrogenase (G6PD) and catalase (CAT) were determined using GSH-Px, GSH-ST, G6PD and CAT commercial kits (Bio-Diagnostic Company, Egypt). All other chemicals were bought from nearby sources with the highest analytical degree.
Animals, experimental design and doses: A total of 30 healthy adult BALB/c male mice weighting 25-30 g/each were purchased from the Theodor Bilharz Institute (Imbaba, Cairo, Egypt) and assigned into groups. All experimental animals preserved in a specific pathogen-free environment. The experimental animal protocols were done according to guidelines accepted by the Institutional Animal Care and Use Committee, which were approved by Assiut University. All procedures concerning the experiment were performed according to the Declaration of Helsinki and the guidelines for the care and use of experimental animals proven by the National Institutes of Health (NIH). Prior to the experiment, all animals for one week were acclimatized in plastic cages (10 animals per cage) inside a well-ventilated room (animal room, Zoology Department, Faculty of Science, Assiut University, the experiment lasted from mid-January 2019 until mid-March of the same year). The animals were kept under typical laboratory conditions (temperature of 23°C, relative humidity of 60-70% and a 12 h light/dark cycle) and were fed a diet of standard commercial pellets (contain 20% crude protein and 11% crude fiber, rich in protein and energy) and water containing libitum. Later, 1 week after acclimation, mice were randomly separated into 3 groups (10 mice each, n = 10).
Animal treatments: Control mice were intraperitoneally injected (i.p.) with olive oil (vehicle; 0.1 mL kg–1 b.wt.) 3 times/weekly for 8 weeks. The CCL4 group received CCL4 (0.1 mL kg–1, i.p.) 3 times/weekly for 8 weeks. The CCL4-curcumin group, intraperitoneally injected with CCL4 (0.1 mL kg–1) and curcumin (200 mg kg–1) was intragastrically administered 3 times/weekly for 8 weeks. The third group (CCL4-curcumin group) intragastrically received curcumin (200 mg kg–1) prior 3 days of CCL4 treatment.
In the second group, toxicity was stimulated in the liver and spleen. All animals received good human care and the study protocols were in agreement with Institutional Guidelines for the use of laboratory animals.
Collection and preparation of samples: All animals were sacrificed at day 57 post-CCL4 injection and blood was collected from the heart to obtain plasma for biochemical studies. We stored the plasma at -80°C until use. Removed liver and spleen, washed in saline (4°C), cut into small pieces. Part was immersed in 10% neutral buffered formalin for histopathological study. Other samples were homogenized in 0.1 M cold phosphate buffered (PBS) (pH 7.4) using IKA Yellow line DI homogenizer (18 Disperser, Germany) at 6000 rpm for 1 h at 4°C. Supernatant was frozen at -20°C for biochemical assays. For RNA extraction and gene expression analysis another samples were suspended in trizol solution.
Analytical measurements: Alkaline phosphatase activity (ALP, IU L–1) were determined as Belfield and Goldberg16, aspartate aminotransferase (AST, U L–1) and alanine aminotransferase (ALT, U L–1) activities were measured by standard spectrophotometric, according to the method described by Reitman and Frankel17. The total protein content in liver and spleen tissues were determined colormitically using the method of Lowry et al.18 Malondialdehyde (MDA) levels in liver and spleen tissues were estimated by the method of Ohkawa et al.19, using thiobarbituric acid (TBA). Glutathione (GSH) levels were measured deepening on the method of Beutler et al.20. Superoxide dismutase (SOD) activity was estimated using the method of Misra and Fridovich21. The CAT, GSH-Px, GSH-ST and G6PD were determined according to the methods described by Aebi22, Paglia and Valentine23, Habig et al.24 and Kornberg et al.25, respectively.
Gene expression analysis: The effect of CUR administration on the expression for some selected genes implicated in the inflammation and toxicity of liver and spleen were studied using real-time quantitative PCR (RT-PCR). We designed the primers used for quantitative (RT-PCR) analysis using the Primer Express 1.5 software (Applied Biosystems). The mouse primers were designed as following: Nox2, Forward: 5’-TCCGTATTGTGGGAGACTGG-3’, Reverse: 5’-AAAGGGCGTG ACTCCAATC-3’; COX-2 Forward: 5’-CCTAGATAACAGAGCCGC TTTC-3’, Reverse: 5’-TTTCACCATAGAATCCAGTCCG-3’; 5-LO Forward: 5’-ATTGCCATCCAGCTCAACCAAACC-3’, Reverse: 5’-TGGCGATACCAAACACCTCAGACA-3’; TNF-α Forward: 5-ATGAGCACAGAAAGCATGA-3, Reverse: 3-AGTAG ACAGAAGAGCGTGGT-5; IL-6 Forward: 5-ACCGAGCTCTGTT GACAAG-3, Reverse: 3-TCCTCGCCACACTTCTCTTT-5; IL-1β Forward: 5-GCACTACAGGCTCCGAGATGAAC-3, Reverse: 3-TTG TCGTTGCTTGGTTCTCCTTGT-5 and normalized using GAPDH Forward: 5-GTTGTCTCCTGCGACTTCA-3, Reverse: 3-GGTGGTCC AGGGTTTCTTA-5. After performing the indicated treatments, RNA was extracted and then the corresponding cDNA was prepared and then real time PCR was applied for gene expression analysis as earlier designated26. All cDNA samples were managed in a 96-well plate using the following cycling conditions: 10 min at 95°C and 40 cycles at 95°C for 15 sec ended by 1 min at 60°C. These data were analyzed according to Livak and Schmittgen27.
Histopathological analysis: After blood withdrawal anesthetized animals were euthanized by cervical dislocation then dissected for extraction of liver and spleen. Pieces of each organs 2×2 mm were fixed in 10% buffered for 24 h and routinely processed for paraffin embedding.
Blocks has been prepared, then sections 5 were cut and stained with hematoxylin and eosin stain H and E28. The stained slices were examined by the light microscope (Olympus CX31, Japan) and photographed using a digital camera (Olympus, Camedia C-5060, Japan).
Statistical analysis: By using one-way analysis of variance (ANOVA) followed by Newman-keuls Multiple Comparison Test as a post-test, we analyzed the results statistically. These analysis were executed using a computer prism program for Windows, version 7.0 (Graph Pad software, Inc, San Diego CA. USA). The level of significance between groups was accepted at p<0.05, 0.01 and the data were expressed as mean±Standard error (SE).
RESULTS
Analytical measurements
Serum function tests: The results indicate that mice administered with CCL4 (0.1 mL kg–1), showed marked hepatic damage (group 2) when compared to control (group 1) this was evidenced by a marked increase in the levels of serum liver enzymes, (AST, ALT and ALP) in (group 2) when compared to control (group 1), co-treatment of mice with CUR (group 3) resulted in a significant decrease in these enzymes (Table1).
Oxidative stress markers
A-Free radicals
A1-Lipid peroxidation (LPO): A significant increase of liver and spleen tissues of malondialdehyde (MDA) in concentrations was observed in group 2, (animals treated with CCL4) as a product of lipid peroxidation, CUR treatment (200 mg kg–1) reduced this CCL4-induced increase in MDA (Table 2).
B-Antioxidants
B1-Enzymatic antioxidants: It was observed that treatment of animals with CCL4 (group 2) resulted in a significant decrease in the activities of CAT, GSH-Px, GSH-ST and SOD in liver and spleen tissues, while activates of G6PD were significantly increased. Co-treatment with CUR (group 3) restored all these activities nearly to the control results (Table 2).
B2-Non-enzymatic antioxidants: The data shown in (Table 2) confirm the decrease in GSH level in CCL4-treated mice, co-treatment with CUR significantly recovered the decreased total GSH level (Table 2).
Gene expression analysis (anti-inflammatory and pro-inflammatory markers): The mRNA expression of TNF-α, IL-6, IL-1, NOX2, COX-2 and 5-LO levels of liver and spleen were significantly increased in mice administrated with CCL4 compared to control group. Significant reduction in all raised parameters was observed when CUR was given along with CCL4 compared with CCL4 treated group (Fig. 1-4).
| Table 1: Effect of CCL4 on liver enzymes in experimental treated rats | |||
| Treatments | |||
| Groups | ALP (U L–1) | AST (U L–1) | ALT (U L–1) |
| Control (olive oil 0.1 mL kg–1) | 83.95 1.091 | 121.8±0.16 | 33.46±0.24 |
| CCL4 (0.1 mL kg–1) | 209.5±1.72a*** | 202.3±0.21a*** | 64.68±0.32a*** |
| Curcumin (200 mg kg–1) | 142.1±1.91b*** | 151.9±0.36b*** | 43.33±0.25b*** |
Values represent Means±SE, aSignificant difference between control and CCL4 treated mice, bSignificant difference between CCL4 and curcumin treated mice, ***p<0.001 | |||
| Table 2: Effect of CCL4 on oxidative stress markers in liver tissue in experimental treated rats | |||
| Parameters | Control | CCL4 | Curcumin |
| LPO (nmol MDA mg–1 protein) liver | 0.831±0.03 | 1.775±0.04a*** | 0.725±0.02b*** |
| LPO (nmol MDA mg–1 protein) spleen | 0.623±0.04 | 2.201±0.14a*** | 0.798±0.03b*** |
| CAT (U mg–1 protein) liver | 85.13±1.2 | 52.14±0.73a *** | 70.87±1.12b *** |
| CAT (U mg–1 protein) spleen | 55.07±1.12 | 24.91±1.049a*** | 43.63±1.71b*** |
| SOD (U mg–1 protein) liver | 14.50±0.28 | 6.558±0.30a*** | 8.832±0.45b** |
| SOD (U mg–1 protein) spleen | 5.797±0.19 | 2.783±0.18a*** | 4.877±0.23b*** |
| G6PD (mU mg–1 protein) liver | 18.02±0.18 | 39.96±0.23a*** | 23.36±0.30b*** |
| G6PD (mU mg–1 protein) spleen | 6.866±0.18 | 25.59±0.28a*** | 17.82±0.21b*** |
| GSH-Px (nmole min–1 mg–1) liver | 109.3±1.8 | 79.86±1.10a*** | 101.0±0.39b*** |
| GSH-Px (nmole min–1 mg–1) spleen | 60.32±0.48 | 38.06±0.99a*** | 53.71±0.58b*** |
| GST (nmole min–1 mg–1) liver | 154.7±1.16 | 75.65±0.90a*** | 113.0±1.18b*** |
| GST (nmole min–1 mg–1) spleen | 95.19±1.03 | 43.42±0.89a*** | 75.23±0.80b*** |
| GSH (μg mg–1 protein) liver | 28.03±1.21 | 16.36±0.77a*** | 22.54±0.82b*** |
| GSH (μg mg–1 protein) spleen | 15.68±1.13 | 8.402±0.96a*** | 11.41±1.13b* |
Values represent Means±SE, aSignificant difference between control and CCL4 treated mice, bSignificant difference between CCL4 and curcumin treated mice, *p<0.05, **p<0.01, ***p<0.001 | |||
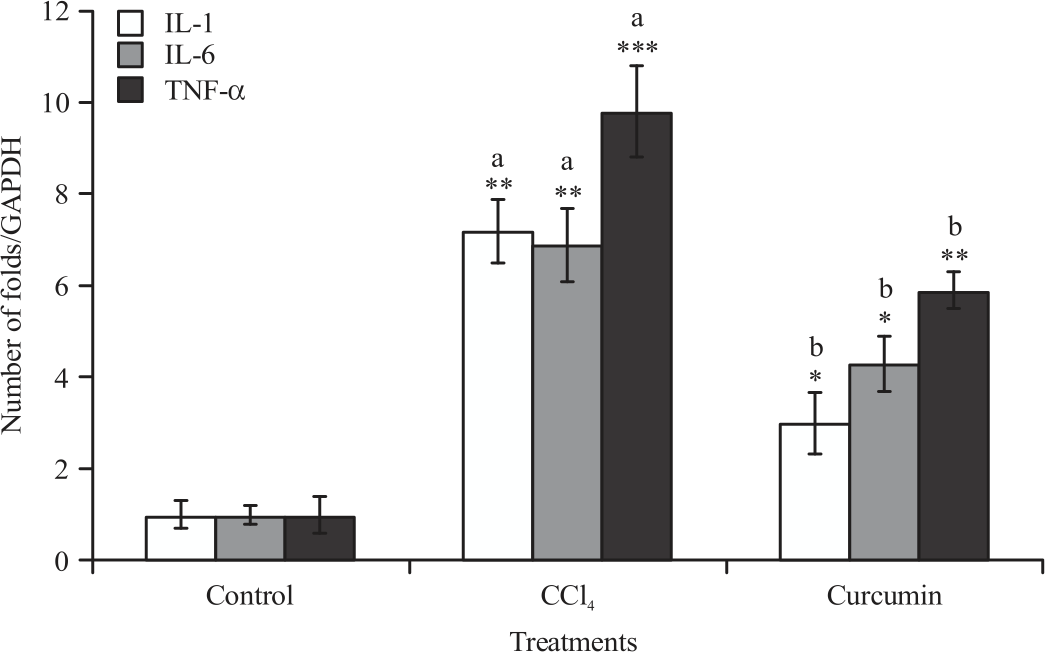 |
| Fig. 1: | Effect of CCL4 and curcumin on IL-1, IL-6 and TNF-α mRNA expression, measured using real time PCR in BALB/c mice liver Data were reported as Mean±SE, n = 8-10, *p<05, **p<0.01, ***p<0.001, aSignificance difference between control group and CCL4 group, bSignificance difference between CCL4 group and curcumin co-treated group |
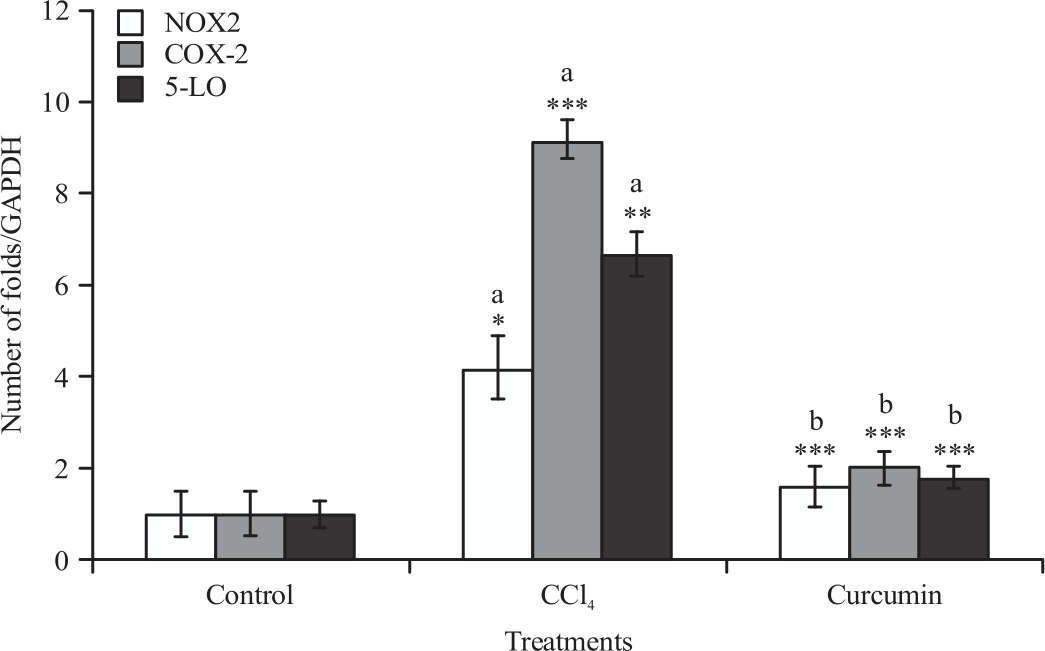 |
| Fig. 2: | Effect of CCL4 and curcumin on NOX2, COX-2 and 5LO mRNA expression, measured using real time PCR in BALB/c mice liver Data were reported as Mean±SE, n = 8-10, *p<05, **p<0.01, ***p<0.001, aSignificance difference between control group and CCL4 group, bSignificance difference between CCL4 group and curcumin co-treated group |
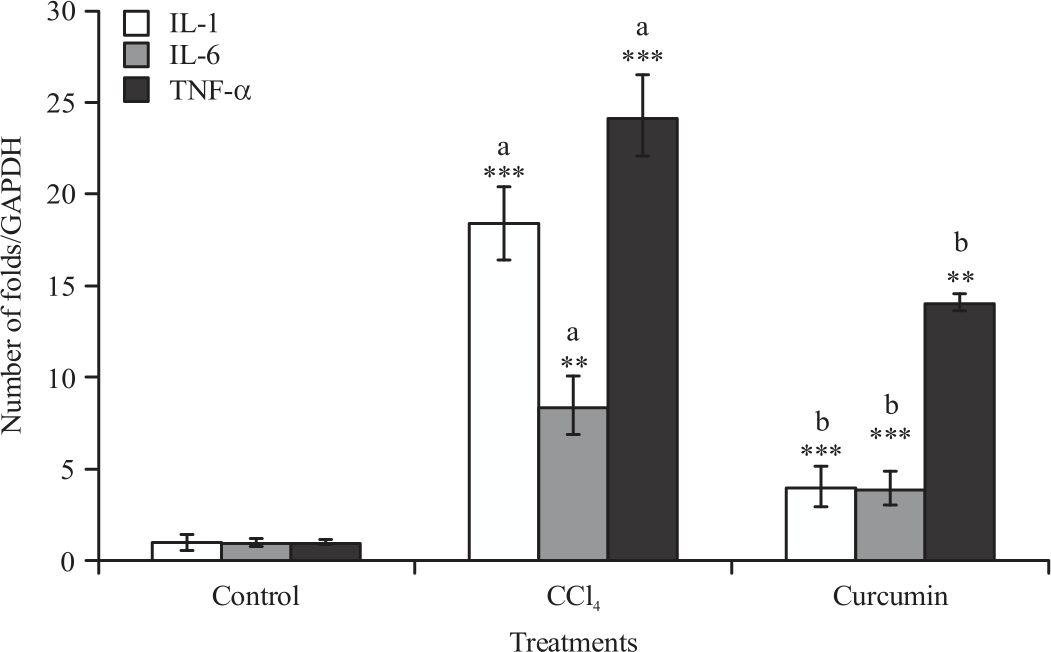 |
| Fig. 3: | Effect of CCL4 and curcumin on IL-1, IL-6 and TNF-α mRNA expression, measured using real time PCR in BALB/c mice spleen Data were reported as Mean±SE, n = 8-10, **p<0.01, ***p<0.001, aSignificance difference between control group and CCL4 group, bSignificance difference between CCL4 group and curcumin co-treated group |
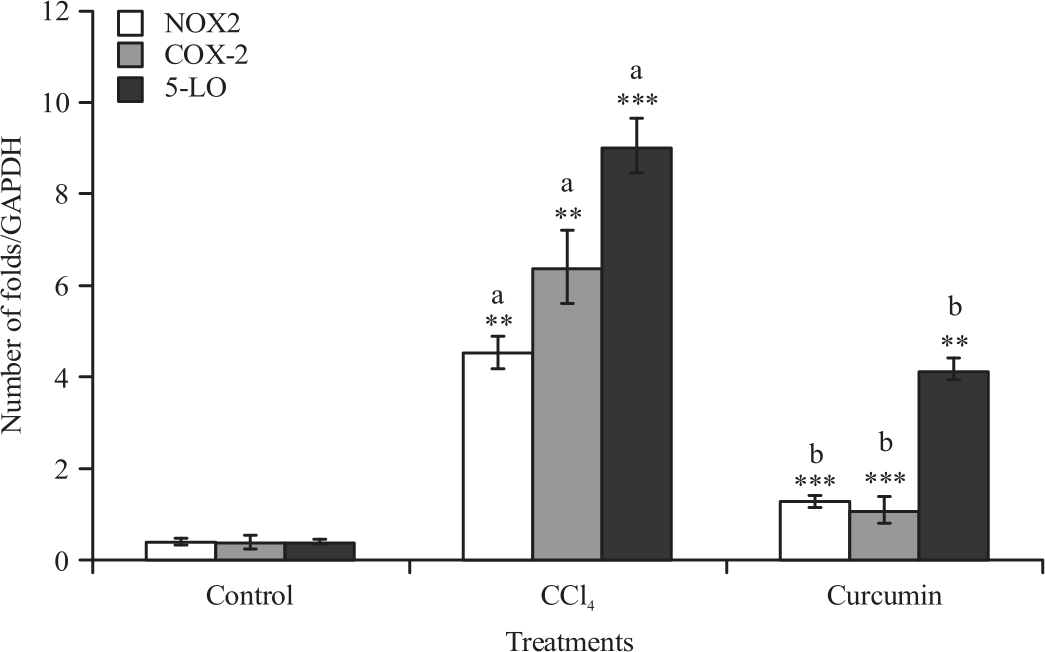 |
| Fig. 4: | Effect of CCL4 and curcumin on NOX2, COX-2 and 5-LO mRNA expression, measured using real time PCR in BALB/c mice spleen Data were reported as Mean±SE, n = 8-10, **p<0.01, ***p<0.001, aSignificance difference between control group and CCL4 group, bSignificance difference between CCL4 group and curcumin co-treated group |
Histopathological analysis: Histopathological findings of the liver were evaluated by light microscopy. Examination of the liver tissues of control rats had normal histoarchitecture of hepatocyte (Fig. 5a, b). Nevertheless, histopathological examination of HE-stained sections revealed that the treatment of male rats with CCL4 showed vascular and cellular changes. The vascular changes were in the form of congestion and dilatation of the central veins (Fig. 5c). The cellular changes were in the form of an infiltration by mononuclear inflammatory cells and diffuse hyperplasia of kupffer cells (Fig. 5d). Moreover, hepatocytes were showed by rarified areas of cytoplasm (Fig. 5e), while examination of liver sections of curcumin treated group showing normal histological appearance (Fig. 5f).
Examination of the spleen tissues of control and olive oil treated rats had normal histoarchitecture (Fig. 6a, b). Histopathological examination of male rats treated with CCL4 showed scattered necrotic areas in the parenchyma and increased number of macrophages were seen (Fig. 6c, d). In addition, there was an apoptotic change in megakaryocyte with fragmented nuclei (Fig. 6e), while examination of spleen sections of curcumin treated groups showed histological appearance nearly similar to the control (Fig. 6f).
DISCUSSION
In this study, the significant rise in serum AST, ALT and ALP activities in CCL4 group as compared with the control group is matched with the findings of Prakash et al.29, Kone et al.30. This outcome is in harmony with the extensive hepatocellular damage noted in the histopathological section of the current study because these hepatic enzymes are cytoplasmic in position and are released into circulation after loss of cell membrane integrity31. Results of the current study demonstrated that CUR caused a significant reduction in serum ALT, ALP and AST activities which return to normal along with healing of hepatic parenchyma and renewal of hepatocytes. Prevention of hepatic enzymes from leakage into the circulation can be attributed to the antioxidant, anti-inflammatory and anti-apoptotic properties of CUR32,33 which has an important role in maintaining the membrane integrity against the cytotoxicity of CCL4.
The CCL4 is metabolized in the endoplasmic reticulum of kupffer cells and hepatocytes involving two phases. In the first phase, it is metabolized by cytochrome P450 gives rise reactive trichloromethyl radicals which in presence of oxygen produces trichloromethyl peroxyl. These free radicals are unstable and immediately react with the membrane components initiating a chain reaction that leads to lipid peroxidation31 which is considered to be one of the main mechanisms of hepatic injury by CCL4. In this study, we found a clear increase in TBARS, an index of LPO, in the liver and spleen of CCL4 challenged mice as previously observed in liver by Ritesh et al.34 and in liver and spleen by Al-Zeiny and Abbas35. Given that GPX reduces lipid peroxides and other organic hydroperoxides, the reduction in this enzymatic antioxidant following exposure to CCL4 based on these findings might encourage overproduction of these cytotoxic mediators32.
CUR significantly reduced the CCL4-induced increase in hepatic and splenic LPO as appeared in the current study, suggesting that CUR could reduce the free radicals production or remove these pro-oxidant molecules36. In fact, the increase in hepatic SOD, CAT and GPX activities in this experimental model denotes improvement in the ability of antioxidant defense system to scavenge superoxide and hydrogen peroxide radicals32.
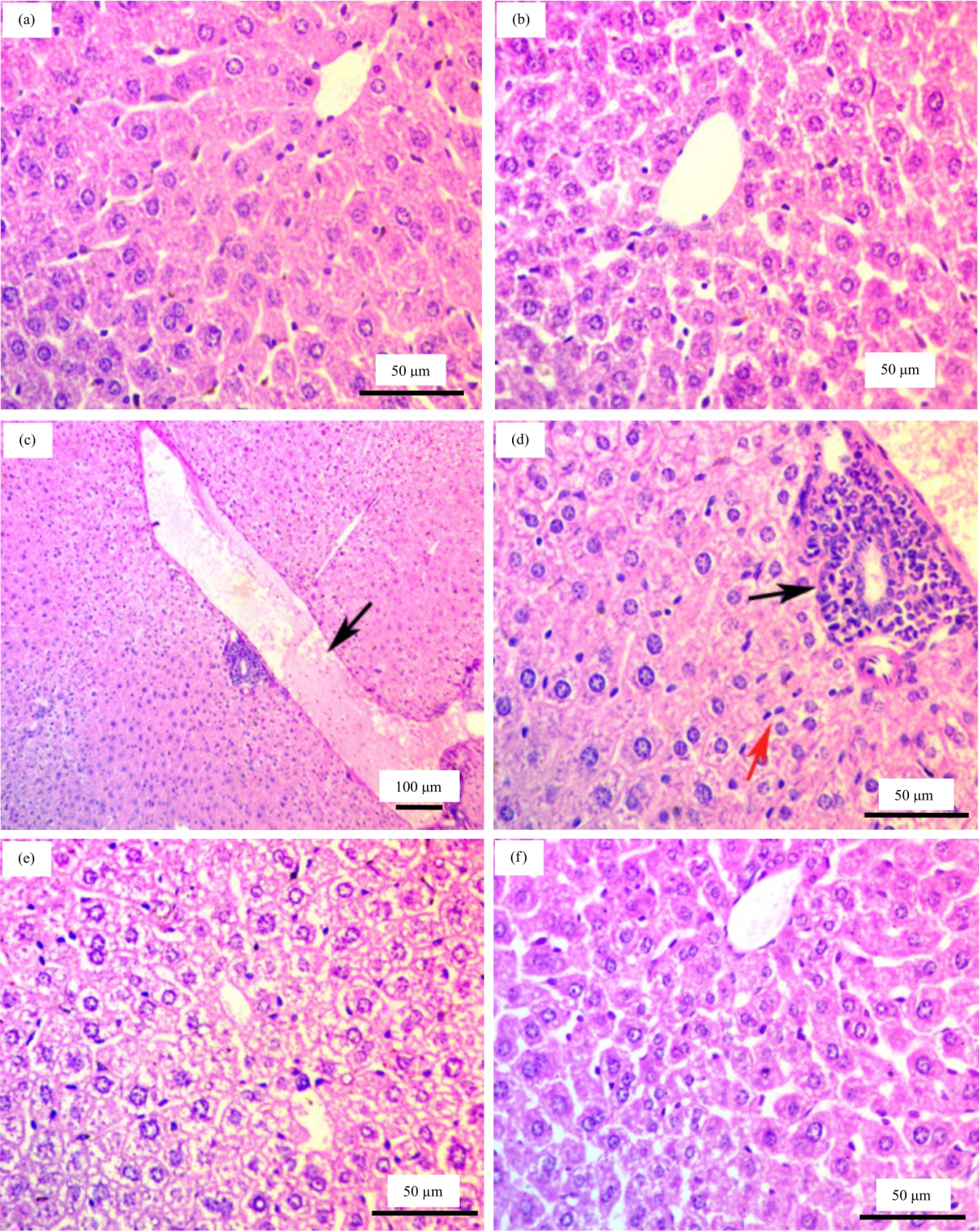 |
| Fig. 5(a-f): | (a, b) Liver of control rats showing normal histological architecture, (c-e) Liver of CCL4 treated group, (c) Showing congestion and dilatation of the central vein (arrow), (d) Magnified part of (c) showing mononuclear cell infiltration (black arrow) and hyperplasia of kupffer cells (red arrow), (e) Showing hepatic cells with positively stained cytoplasmic clumps separated by rarified areas of cytoplasm and (f) Liver of curcumin treated group showing normal appearance |
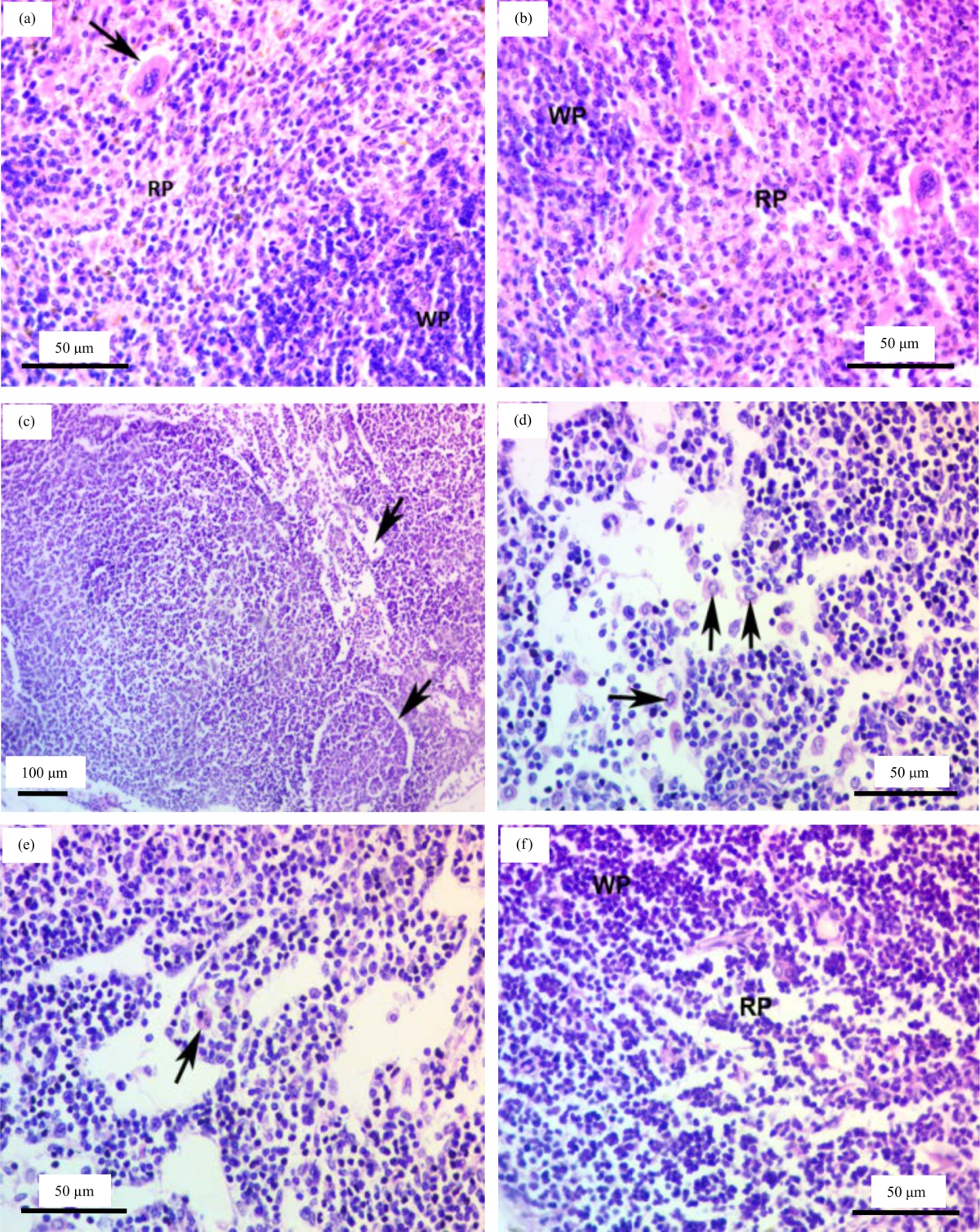 |
| Fig. 6(a-f): | (a, b) Spleen of control rats showing normal parenchyma (normal architecture) consists of white pulp (WP), red pulp (RP) and megakaryocyte with characteristic lobulated nucleus and acidophilic cytoplasm (arrow), (c-e) Spleen of CCL4 treated group showing scattered necrotic areas in the parenchyma (arrows), (d) Large number of macrophages are present (arrows), (e) Apoptotic changes in megakaryocyte with fragmented nuclei (arrow) and (f) Spleen of curcumin treated group showing cellular architecture nearly similar to the control |
Liver GSH was significantly reduced in CCL4 treated mice in the present study and this could be due to enhancing substrate utilization by glutathione peroxidase, as there is a direct correlation between GSH depletion and elevated lipid peroxidation37. A significant reduction in the activities of GST, GSH-Px, SOD and CAT had been recorded in the liver and spleen of CCL4 treated mice. it is well known that GST implicated in preserving the cells against ROS mediated injury by detoxification of lipid hydro peroxides formed due to oxidative damage38, it can be hypothesized that the reduction in its activity add another contributory factor in stimulating the lipid peroxidation in the current hepato- and splenotoxic mice model. Such alterations were also recorded by many studies39-41 and these results suggested that CCL4 administration make the liver and spleen more vulnerable to the attack of free radicals.
Consumption of CUR in the present study led to recovery of the redox balance state by reactivating the impaired activity of important antioxidant enzymes and restoring the levels of non-enzymatic antioxidants as observed in other hepatocellular toxicity models induced by cadmium42 or paracetamol43. This result may attribute to up-regulation of antioxidant enzyme gene expression as that found in aflatoxin B1-intoxicated rats treated with CUR35.
The second phase of CCL4 hepatotoxicity involved the inflammatory responses which play an important role. Some hepatic cells as kupffer cells, hepatic stellate cells (HSCs) and sinusoidal endothelial cells can be activated to release cytokines which moderate liver fibrogenesis, activate of HSCs, regulate expression and deposition of matrix proteins and modify the renewal of hepatocytes 44, 45.
The present study approved that the pro-inflammatory cytokines IL-1β, TNF-α and IL-6 were highly expressed in the liver and spleen of CCL4-treated mice as previously estimatesd46. These cytokines act both as a part of immune reactivity to hepatocellular damage and as a cytopathological mediator incriminated in inducing further hepatic histoarchitecture deteriorations47. In this study, the elevation in the expression levels of these cytokines is a clear biochemical footprint parallel to the perturbations in hepatic and splenic histopathological features. The potential increase in oxidants secondary to the suppression in antioxidant defense mechanism in the current experiment is suggested to induce the gene expression of pro-inflammatory cytokines, e.g., hydrogen peroxide induces the transcript levels of IL-1β and TNF-α48.
In our study, the significant inhibition in the expression of CCL4 –induced pro-inflammatory cytokines under investigation following CUR consumption is consistent with that found in C57BL/6 mice49. CUR down-regulates TNF- α by suppressing its various inducers50, modulating methylation of its promoter51, inhibiting p300/CREB-specific acetyl transferase52 and decreasing NF-kB53.
The expression of 5-LO in the liver is restricted to macrophages, that is required for the development of acute liver failure54. In our study marked elevation in mRNA 5-LO expression in liver and spleen have been found in CCL4-treated mice.
Inhibition of 5-LO gene expression by CUR supplementation in our study could play a fundamental role in arresting the growth and inducing the apoptosis in kupffer cell, thereby inhibits hyperplasia in these cells and subsequently prevents hepatotoxic necro-inflammatory injury induced by CCL455, 56.
The increase in the production of pro-inflammatory cytokines (e.g., IL-1β and TNF-α) may cause up-regulation of COX-2 expression57, following CCL4 exposure in our experimental model. The suppression in COX-2 gene expression by CUR administration might indicate its ability to excrete hepatoprotective effect on CCL4-induced liver damage via inhibition of prostaglandin synthesis and amelioration of inflammatory responses. Inhibition of nuclear factor-Kappa B is supposed to be one of the etiological factors implicated in down-regulation58 of COX-2.
In this study, over expression of NOX2 could enhance nuclear factor kappa B activation which explains the concomitant over expression of COX-2 in the liver and spleen of CCL4 challenged mice and resulted in ROS-mediated liver fibrosis59.
The obvious reduction in NOX2 gene expression in the liver and spleen of CCL4 intoxicated mice following administration of CUR might suppress free radicals generation and lipid peroxidation trying to protect against hepatic fibrosis60.
The histopathological changes in the hepatic features following CCL4 intoxication is in parallel with that observed by Ma et al.61 and Khedr and Khedr 62. Mononuclear cell infiltration in the liver and spleen of intoxicated mice reflects the non-specific immune response of leucocytes and macrophages to the foreign bodies. As soon as, tissue damaged, leukocytes rapidly move to the injury sites initiating an inflammatory response. Therefore, leukocyte infiltration was considered as a marker of inflammatory response63. The CUR efficaciously suppressed leucocyte infiltration, so it was suggested that CUR not only dampen the inflammatory response on the level of pro-inflammatory mediators but also on the level of cellular microenvironment64. Kupffer cell hyperplasia observed in the CCL4 challenged liver represents one of defensive tools of detoxification and is contributed to hepatic oxidative stress65. Activation of JNK, c-Jun and caspase-3, increased expression of pro-apoptotic proteins and reduced expression of anti-apoptotic protein (Bcl-2)66, are the potential underlying mechanistic ways behind appearance of apoptotic changes in megakaryocyte with presence of fragmented nuclei in the spleen of CCL4 exposed mice.
It was very clear in this study that he antioxidant, anti-inflammatory and anti-apoptotic properties of CUR played a pivotal role in returning the cellular architecture of liver and spleen of CCL4-induced toxic model nearly similar to the control as previously reported62. The CUR can be beneficial for preventing spleen toxicity by suppressing the cellular reactive oxidants generation, up-regulating the levels of the cellular antioxidant enzymes and protecting the cells against oxidative damage of proteins and lipids, protects cells from stress induced MAPK dependent and p53 mediated intrinsic apoptosis67. Inhibition of kupffer cells activation and hepatic oxidative stress and arresting the intrinsic apoptotic pathway, are believed to contribute to the ability of CUR to limit hepatocellular damage mediated by CCL468,69.
CONCLUSION
These findings provide compelling evidences that CCL4 disrupted the redox potential, caused up-regulation in pro-inflammatory cytokines and cytopathogenic inducers and histoarchitecture deteriorations in the liver and spleen of mice. CUR effectively limited the above mentioned disorders and therefore can be used as a promising therapeutic strategy offering new opportunities for treatment of hepatotoxic and spleenotoxic-associated CCL4 affections. However, further studies are highly recommended to investigate in depth the other molecular pathways underlying the antioxidant and anti-inflammatory effects of CUR in this experimental model.
REFERENCES
- Duval, F., J.E. Moreno-Cuevas, M.T. González-Garza, C. Rodríguez-Montalvo and D.E. Cruz-Vega, 2014. Protective mechanisms of medicinal plants targeting hepatic stellate cell activation and extracellular matrix deposition in liver fibrosis. Chin. Med., Vol. 9.
CrossRefDirect Link - Li, Z., J. Sun and X. Yang, 2015. Recent advances in molecular magnetic resonance imaging of liver fibrosis. BioMed Res. Int., Vol. 2015.
CrossRefDirect Link - Enomoto, M., H. Morikawa, A. Tamori and N. Kawada, 2014. Noninvasive assessment of liver fibrosis in patients with chronic hepatitis B. World J. Gastroenterol., 20: 12031-12038.
CrossRefPubMedDirect Link - Xuan, J., S. Chen, B. Ning, W.H. Tolleson and L. Guo, 2015. Development of HepG2-derived cells expressing cytochrome P450s for assessing metabolism-associated drug-induced liver toxicity. Chem. Biol. Interact., 5: 63-73.
CrossRefPubMedDirect Link - Birch-Machin, M. and A. Bowman, 2016. Oxidative stress and ageing. Br. J. Dermatol., 175: 26-29.
CrossRefDirect Link - Huang, Z.G., W.R. Zhai, Y.E. Zhang and X.R. Zhang, 1998. Study of heteroserum-induced rat liver fibrosis model and its mechanism. World J. Gastroenterol., 4: 206-209.
CrossRefDirect Link - Zhang, D.G., C. Zhang, J.X. Wang, B.W. Wang and H. Wang et al., 2017. Obeticholic acid protects against carbon tetrachloride-induced acute liver injury and inflammation. Toxicol. Applied Pharmacol., 314: 39-47.
CrossRefPubMedDirect Link - Li, L., M. Duan, W. Chen, A. Jiang, X. Li, J. Yang and Z. Li, 2017. The spleen in liver cirrhosis: Revisiting an old enemy with novel targets. J. Transl. Med., Vol. 15.
CrossRefDirect Link - Neto, Z., M. Machado, A. Lindeza, V. do Rosário, M.L. Gazarini and D. Lopes, 2013. Treatment of Plasmodium chabaudi parasites with curcumin in combination with antimalarial drugs: Drug interactions and implications on the ubiquitin/proteasome system. J. Parasitol. Res., Vol. 2013.
CrossRefDirect Link - O'Rourke, J.M., V.M. Sagar, T. Shah and S. Shetty, 2018. Carcinogenesis on the background of liver fibrosis: Implications for the management of hepatocellular cancer. World J. Gastroenterol., 24: 4436-4447.
CrossRefPubMedDirect Link - Morinaga, A., T. Ogata, M. Kage, H. Kinoshita and S. Aoyagi, 2010. Comparison of liver regeneration after a splenectomy and splenic artery ligation in a dimethylnitrosamine-induced cirrhotic rat model. HPB., 12: 22-30.
CrossRefPubMedDirect Link - Mitra, S.K., S.J. Seshadri, M.V. Venkantaranganna, S. Gopumaaaaaadhavan, U.V. Udapa and D.N.K. Sarma, 2000. Effect of HD-03-a herbal formulation in galactosamine-induced hepatopathy in rats. Indian J. Physiol. Pharmacol., 44: 82-86.
PubMedDirect Link - Eybe, V., D. Kotyzova and J. Koutensky, 2006. Comparative study of natural antioxidants-curcumin, resveratrol and melatonin-in cadmium-induced oxidative damage in mice. Toxicology, 255: 150-156.
CrossRefPubMedDirect Link - Phan, T.T., P. See, S.T. Lee and S.Y. Chan, 2001. Protective effects of curcumin against oxidative damage on skin cells in vitro: Its implication for wound healing. J. Trauma, 51: 927-931.
CrossRefPubMedDirect Link - Biswas, S.K., D. McClure, L.A. Jimenez, I.L. Megson and I. Rahman, 2005. Curcumin induces glutathione biosynthesis and inhibits NF-κB activation and interleukin-8 release in alveolar epithelial cells: Mechanism of free radical scavenging activity. Antioxid. Redox Signal., 7: 32-41.
CrossRefPubMedDirect Link - Reitman, S. and S. Frankel, 1957. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol., 28: 56-63.
CrossRefPubMedDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Ohkawa, H., N. Ohishi and K. Yagi, 1979. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem., 95: 351-358.
CrossRefPubMedDirect Link - Beutler, E., O. Duron and B.M. Kelly, 1963. Improved method for the determination of blood glutathione. J. Lab. Clin. Med., 61: 882-888.
PubMedDirect Link - Misra, H.P. and I. Fridovich, 1972. The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. J. Biol. Chem., 247: 3170-3175.
CrossRefPubMedDirect Link - Aebi, H., 1984. Catalase in vitro. In: Methods in Enzymology, Packer, L., Academic Press, Cambridge, Massachusetts, United States, ISBN: 9780121820053, pp: 121-126.
CrossRefDirect Link - Paglia, D.E. and W.N. Valentine, 1967. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med., 70: 158-169.
CrossRefPubMedDirect Link - Habig, W.H., M.J. Pabst and W.B. Jakoby, 1974. Glutathione S-transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem., 249: 7130-7139.
CrossRefPubMedDirect Link - Kornberg, A., S.R. Kornberg and E.S. Simms, 1956. Metaphosphate synthesis by an enzyme from Escherichia coli. Biochim. Biophys. Acta, 20: 215-227.
CrossRefDirect Link - Ismail, I.A., H.S. Kang, H.J. Lee, H. Chang and J. Yun et al., 2013. 2-Hydroxycinnamaldehyde inhibits the epithelial-mesenchymal transition in breast cancer cells. Breast Cancer Res. Treat., 137: 697-708.
CrossRefPubMedDirect Link - Livak, K.J. and T.D. Schmittgen, 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods, 25: 402-408.
CrossRefDirect Link - Prakash, O., G.N. Singh, R.M. Singh, S.C. Mathur, M. Bajpai and S. Yadav, 2008. Protective effect of a herbal formula against carbontetrachloride induced hepatotoxicity. Int. J. Pharmacol., 4: 282-286.
CrossRefDirect Link - Kone, S.D., F.A. Yapo, G. Gnahoue and F.H. Yapi, 2017. Evaluation of doses-response and combined preventive effects of zinc and vitamin D on liver toxicity induced by carbon tetrachloride in wistar rats. Int. J. Biochem. Biophys., 5: 46-52.
Direct Link - Recknagel, R.O., E.A. Glende, Jr., J.A. Dolak and R.L. Waller, 1989. Mechanisms of carbon tetrachloride toxicity. Pharmacol. Ther., 43: 139-154.
CrossRefPubMedDirect Link - El‐Bahr, S.M., 2015. Effect of curcumin on hepatic antioxidant enzymes activities and gene expressions in rats intoxicated with aflatoxin B1. Phytother. Res., 29: 134-140.
CrossRefDirect Link - Fan, Z., J. Yao, Y. Li, X. Hu, H. Shao and X. Tian, 2015. Anti-inflammatory and antioxidant effects of curcumin on acute lung injury in a rodent model of intestinal ischemia reperfusion by inhibiting the pathway of NF-Kb. Int. J. Clin. Exp. Pathol., 8: 3451-3459.
PubMedDirect Link - Ritesh, K.R., A. Suganya, H.V. Dileepkumar, Y. Rajashekar and T. Shivanandappa, 2015. A single acute hepatotoxic dose of CCl4 causes oxidative stress in the rat brain. Toxicol. Rep., 2: 891-895.
CrossRefDirect Link - Al-Zeiny, S.S.M. and D.A. Abbas, 2017. Comparative histological study of protective effect of oil and alcoholic extracts of dry palm dates and leaves (Phoenix dactylifera L) against CCL4 induced oxidative stress in rats. Kufa J. Vet. Med. Sci., 8: 79-89.
Direct Link - Soto-Urquieta, M.G., S. López-Briones, V. Pérez-Vázquez, A. Saavedra-Molina, G.A. González-Hernández and J. Ramírez-Emiliano, 2014. Curcumin restores mitochondrial functions and decreases lipid peroxidation in liver and kidneys of diabetic db/db mice. Biol. Res., Vol. 47.
CrossRefDirect Link - Al-Shabanah, O.A., K. Alam, M.N. Nagi, A.C. Al-Rikabi and A.M. Bekairi, 2000. Protective effect of aminoguanidine, a nitric oxide synthase inhibitor, against carbon tetrachloride induced hepatotoxicity in mice. Life Sci., 66: 265-270.
CrossRefPubMedDirect Link - Yang, Y., J.Z. Cheng, S.S. Singhal, M. Saini, U. Pandya, S. Awasthi and Y.C. Awasthi, 2001. Role of glutathione S-transferases in protection against lipid peroxidation: Overexpression of hGSTA2-2 IN K562 cells protects against hydrogen peroxide-induced apoptosis and inhibits Jnk and caspase 3 activation. J. Biol. Chem., 276: 19220-19230.
CrossRefDirect Link - Anilakumar, K.R., K.R.S. Krishna, G. Chandramohan, F. Khanum and A.S. Bawa, 2007. Bees wax polyphenols as suppressor of CCl4-induced oxidative stress in rats. Indian J. Physiol. Pharmacol., 51: 361-367.
PubMedDirect Link - Franco, R. and J.A. Cidlowski, 2009. Apoptosis and glutathione: Beyond an antioxidant. Cell Death Differ., 16: 1303-1314.
CrossRefPubMedDirect Link - Circu, M.L. and T.Y. Aw, 2012. Glutathione and modulation of cell apoptosis. Biochim. Biophys. Acta, 1823: 1767-1777.
CrossRefPubMedDirect Link - Shapiro, H., M. Ashkenazi, N. Weizman, M. Shahmurov, H. Aeed and R. Bruck, 2006. Curcumin ameliorates acute thioacetamide‐induced hepatotoxicity. J. Gastroenterol. Hepatol., 21: 358-366.
CrossRefDirect Link - Khorsandi, L., E. Mansouri, M. Orazizadeh and Z. Jozi, 2016. Curcumin attenuates hepatotoxicity induced by zinc oxide nanoparticles in rats. Balkan Med. J., 33: 252-257.
CrossRefPubMedDirect Link - Friedman, S.L., 1999. Cytokines and fibrogenesis. Semin. Liver. Dis., 19: 129-140.
CrossRefPubMedDirect Link - Shi, J., K. Aisaki, Y. Ikawa and K. Wake, 1998. Evidence of hepatocyte apoptosis in rat liver after the administration of carbon tetrachloride. Am. J. Pathol., 153: 515-525.
CrossRefDirect Link - Makni, M., Y. Chtourou, H. Fetoui, E.M. Garoui, T. Boudawara and N. Zeghal, 2011. Evaluation of the antioxidant, anti-inflammatory and hepatoprotective properties of vanillin in carbon Tetrachloride-treated rats. Eur. J. Pharmacol., 668: 133-139.
CrossRefDirect Link - Zhang, G.L., Y.H. Wang, H.L. Teng and Z.B. Lin, 2001. Effects of aminoguanidine on nitric oxide production induced by inflammatory cytokines and endotoxin in cultured rat hepatocytes. World J. Gastroenterol., 7: 331-334.
CrossRefPubMedDirect Link - Chen, M.P., S.H. Yang, C.H. Chou, K.C. Yang, C.C. Wu, Y.H. Cheng and F.H. Lin, 2010. The chondroprotective effects of ferulic acid on hydrogen peroxide-stimulated chondrocytes: Inhibition of hydrogen peroxide-induced pro-inflammatory cytokines and metalloproteinase gene expression at the mRNA level. Inflamm. Res., 59: 587-595.
CrossRefPubMedDirect Link - Huang, S.S., D.Z. Chen, H. Wu, R.C. Chen and S.J. Du et al., 2016. Cannabinoid receptors are involved in the protective effect of a novel curcumin derivative C66 against CCl4-induced liver fibrosis. Eur. J. Pharmacol., 779: 22-30.
CrossRefPubMedDirect Link - Jain, S.K., J. Rains, J. Croad, B. Larson and K. Jones, 2009. Curcumin supplementation lowers TNF-α, IL-6, IL-8 and MCP-1 secretion in high glucose-treated cultured monocytes and blood levels of TNF-α, IL-6, MCP-1, glucose and glycosylated hemoglobin in diabetic rats. Antioxidants Redox Signaling, 11: 241-249.
CrossRefDirect Link - Reuter, S., S.C. Gupta, B. Park, A. Goel and B.B. Aggarwal, 2011. Epigenetic changes induced by curcumin and other natural compounds. Genes Nutr., 6: 93-108.
CrossRefDirect Link - Balasubramanyam, K., R.A. Varier, M. Altaf, V. Swaminathan, N.B. Siddappa, U. Ranga and T.K. Kundu, 2004. Curcumin, a novel p300/CREB-binding protein-specific inhibitor of acetyltransferase, represses the acetylation of histone/nonhistone proteins and histone acetyltransferase-dependent chromatin transcription. J. Biol. Chem., 279: 51163-51171.
CrossRefPubMedDirect Link - Singh, S. and B.B. Aggarwal, 1995. Activation of transcription factor NF-κB is suppressed by curcumin (diferuloylmethane). J. Biol. Chem., 270: 24995-25000.
PubMedDirect Link - Li, L., Y.R. Liu, S. Gao, J.F. Li and S.S. Li et al., 2014. Inhibition of 5-lipoxygenase pathway attenuates acute liver failure by inhibiting macrophage activation. J. Immunol. Res., Vol. 2014.
CrossRefDirect Link - Titos, E., J. Clària, A. Planagumà, M. López-Parra and N. Villamor et al., 2003. Inhibition of 5-lipoxygenase induces cell growth arrest and apoptosis in rat Kupffer cells: Implications for liver fibrosis. FASEB J., 17: 1745-1747.
CrossRefPubMedDirect Link - Titos, E., J. Claria, A. Planaguma, M. Lopez-Parra and A. Gonzalez-Periz et al., 2005. Inhibition of 5-lipoxygenase-activating protein abrogates experimental liver injury: Role of Kupffer cells. J. Leukoc. Biol., 78: 871-878.
CrossRefPubMedDirect Link - Kuo, C.L., C.W. Chi and T.Y. Liu, 2004. The anti-inflammatory potential of berberine in vitro and in vivo. Cancer Lett., 203: 127-137.
CrossRefPubMedDirect Link - Surh, Y.J., K.S. Chun, H.H. Cha, S.S. Han, Y.S. Keum, K.K. Park and S.S. Lee, 2001. Molecular mechanisms underlying chemopreventive activities of anti-inflammatory phytochemicals: Down-regulation of COX-2 and iNOS through suppression of NF-κB activation. Mutat. Res./Fund. Mol. Mech. Mutagen., 480-481: 243-268.
CrossRefDirect Link - Lan, T., T. Kisseleva and D.A. Brenner, 2015. Deficiency of NOX1 or NOX4 prevents liver inflammation and fibrosis in mice through inhibition of hepatic stellate cell activation. PloS One, Vol. 10, No. 7.
CrossRefDirect Link - Paik, Y.H., K. Iwaisako, E. Seki, S. Inokuchi and B. Schnabl et al., 2011. The nicotinamide adenine dinucleotide phosphate oxidase (NOX) homologues NOX1 and NOX2/gp91phox mediate hepatic fibrosis in mice. Hepatology, 53: 1730-1741.
CrossRefPubMedDirect Link - Ma, J.Q., C.M. Liu, Z.H. Qin, J.H. Jiang and Y.Z. Sun, 2011. Ganoderma applanatum terpenes protect mouse liver against benzo(α)pyren-induced oxidative stress and inflammation. Environ. Toxicol. Pharmacol., 31: 460-468.
CrossRefPubMedDirect Link - Khedr, N.F. and E.G. Khedr, 2014. Antioxidant and anti-inflammatory effects of curcumin on CCl4-induced liver fibrosis in rats. Am. J. Biomed. Sci., 6: 191-200.
CrossRefDirect Link - Ma, J.Q., J. Ding, L. Zhang and C.M. Liu, 2014. Hepatoprotective properties of sesamin against CCl4 induced oxidative stress-mediated apoptosis in mice via JNK pathway. Food Chem. Toxicol., 64: 41-48.
CrossRefPubMedDirect Link - Abdelhalim, M.A.K. and B.M. Jarrar, 2011. Gold nanoparticles administration induced prominent inflammatory, central vein intima disruption, fatty change and Kupffer cells hyperplasia. Lipids Health Dis., Vol. 10.
CrossRef - Neyrinck, A., 2004. Modulation of Kupffer cell activity: Physio-pathological consequences on hepatic metabolism. Bull. Mem. Acad. R. Med. Belg., 159: 358-366.
PubMedDirect Link - Brar, S.S., T.P. Kennedy, A.B. Sturrock, T.P. Huecksteadt and M.T. Quinn et al., 2002. NADPH oxidase promotes NF-κB activation and proliferation in human airway smooth muscle. Am. J. Physiol.-Lung Cell. Mol. Physiol., 282: L782-L795.
CrossRefPubMedDirect Link - Rashid, K., S. Chowdhury, S. Ghosh and P.C. Sil, 2017. Curcumin attenuates oxidative stress induced NFκB mediated inflammation and endoplasmic reticulum dependent apoptosis of splenocytes in diabetes. Biochem. Pharmacol., 143: 140-155.
CrossRefPubMedDirect Link - Lee, H.Y., S.W. Kim, G.H. Lee, M.K. Choi and H.W. Jung et al., 2016. Turmeric extract and its active compound, curcumin, protect against chronic CCl4-induced liver damage by enhancing antioxidation. BMC Complement. Altern. Med., Vol. 16.
CrossRefDirect Link - Choudhury, S.T., N. Das, S. Ghosh, D. Ghosh, S. Chakraborty and N. Ali, 2016. Vesicular (liposomal and nanoparticulated) delivery of curcumin: A comparative study on carbon tetrachloride-mediated oxidative hepatocellular damage in rat model. Int. J. Nanomed., 11: 2179-2193.
CrossRefPubMedDirect Link