
S.A. Abdel-Fattah
Department of Poultry Production, Faculty of Agriculture, Ain Shams University, Shoubra El-Kheima, 11241 Cairo, Egypt
LiveDNA: 20.8399
M.I. Shourrap
Department of Poultry Production, Faculty of Agriculture, Ain Shams University, Shoubra El-Kheima, 11241 Cairo, Egypt
LiveDNA: 20.22655
Mona A. Hemida
Department of Poultry Production, Faculty of Agriculture, Ain Shams University, Shoubra El-Kheima, 11241 Cairo, Egypt
LiveDNA: 20.22791
Asian Journal of Animal and Veterinary Advances
Year: 2018 | Volume: 13 | Issue: 4 | Page No.: 369-376







ABSTRACT
Background and Objective: Manipulating the incubation temperature, particularly during the late stage of embryonic development can strongly affect the performance of hatching chicks at later ages. This study was conducted to evaluate the influences of thermal conditioning during embryonic development and at an early age post-hatching on growth performance, metabolic hormones and the skeletal muscle profile of broiler chicks. Materials and Methods: A total of 510 hatching broiler breeder eggs were obtained from Cobb-500 broiler breeder flock at 48 weeks of age. At embryonic day 16 (E16) post candling and infertile exclusion, the eggs were randomly divided into two main groups, each of 240 eggs. The first group was set at optimal incubation conditions (T0), while the second (T1) was exposed to thermal conditioning (TC) at 39.5±0.1°C for 5 h/day at E16, E17 and E18. On day 4 post-hatching the (T0 and T1) groups were each randomly sub-divided into two sub-groups. One sub-group of each main group was exposed in a controlled chamber to TC at 38.0±0.1°C and 42.0% RH for 5 h/day through the 4th, 5th and 6th day of age, while the other sub-group was kept under optimal brooding conditions, resulting in four final sub-groups, T0T0 (optimal incubation and brooding conditions), T1T0 (embryonic TC), T0T1 (post hatch TC) and T1T1 (pre- and post-hatch TC), all were reared up to the age of 42 days. Data on growth performance, metabolic hormone concentrations and histomorphometric measurements of the myocytes of the pectoral muscles were analyzed with a one-way analysis of variance. Results: The results showed that all the TC groups, especially T1T1 had higher (p<0.01) growth and carcass performance than the non TC group (T0T0). With pre- and postnatal TC, whether separately or in combination, the IGF-I level was increased, T4 was reduced, while the T3 level did not change significantly at 42 days. In addition, the major pectoralis muscles of T1T0 and T1T1 chicks possessed greater number and histometric measurements of myocytes compared to those of the other groups. Conclusion: Pre- and postnatal TC both separately or in combination could enhance the productive performance and breast muscle development of broiler chicks.
PDF Abstract XML References Citation
Received: October 13, 2017;
Accepted: April 23, 2018;
Published: June 15, 2018
How to cite this article
S.A. Abdel-Fattah, M.I. Shourrap and Mona A. Hemida, 2018. Effect of Pre- and Post-hatch Thermal Conditioning on Productive Performance, Some Metabolic Hormones and Breast Muscles Growth of Broilers Chicks. Asian Journal of Animal and Veterinary Advances, 13: 369-376.
DOI: 10.3923/ajava.2018.369.376
URL: https://scialert.net/abstract/?doi=ajava.2018.369.376
DOI: 10.3923/ajava.2018.369.376
URL: https://scialert.net/abstract/?doi=ajava.2018.369.376
INTRODUCTION
In birds as in mammals, growth and muscle weight is a compiled function of several aspects, including prenatal muscle development and postnatal growth. During embryogenesis, the number of muscle fibers continues to increase; this increase stops shortly after hatch/birth and further muscle growth was mainly due to the increase in its fiber length and diameter, (hypertrophy). In this, any technology that could enhance skeletal muscle growth during this period would help to maintain the progress of growth performance in the future1. Moreover, changes in muscle growth associated with environmental or nutritional manipulations during the first days post-hatch (time of satellite cells activation) had an immediate stimulatory effect on satellite cells proliferation and differentiation, as well as increased body weight and percentage of breast muscle out of total body weight on 42 days of age2.
Halevy et al.1 showed that thermal conditioning (TC) of chick embryos from 8-10 or from 16-18 days of incubation at 38.5°C for 3 h/day, enhanced myoblast proliferation in post-hatching chicks. Therefore, incubation temperature efficiently affects not only the embryonic and the hatchability traits, but also the post-hatch growth and performance of chicks, in this respect Abdel-Fattah and Shourrap3 illustrated that, compared to chicks from optimal incubation conditions, chicks delivered from TC eggs during embryogenesis had significantly higher live body weight, body weight gain and better feed conversion ratio at hatch and all ages thereafter. Also, Piestun et al.4 found that successful TC at 38.1°C from E0-E5 significantly increased the market body weight of broiler chicks. Additionally, similar effects could be established with early post-hatching TC5,6 during which body temperature regulation is still immature, this may lead to greater adaptability to high temperatures in the final rearing period. Also, Abd El Azim7 showed that the body weight, weight gain and feed conversion ratio of Ross broiler chicks improved after they were subjected to 40°C for 4 h/day during the first 14 days of age (DOA).
To date, most studies evaluating the effect of thermal conditioning have conducted the pre-hatching and pos-thatching experiments independently of each other. Therefore, the definite effectiveness of each of these protocols remains unknown. Furthermore, this study aimed to establish whether a combination of pre- and post-hatching thermal conditioning may offer further benefits regarding either productive performance or related metabolic hormones.
MATERIALS AND METHODS
This study was carried out at the Agricultural Experiments and Research Station, Faculty of Agriculture, Ain Shams University, from May-September, 2016.
Experimental procedures: A total of 510 broiler breeder eggs with an average weight of 68.54±0.79 g were obtained from a commercial broiler breeder flock (Cobb-500) at 48 weeks of age (WOA). All the eggs were incubated under optimal conditions at 37.6±0.1°C and 65% relative humidity (RH) in a locally manufactured automatic incubator.
Prenatal thermal conditioning procedures: At embryonic day 16 (E16), the eggs were candled, so that infertile eggs and early dead embryos could be identified and excluded. Then, the remaining 480 eggs were randomly divided into two equal main groups, each of 240 eggs. The first group of eggs was set under the optimal incubation conditions and served as a control (T0), while the second group (T1) was transferred to another automatic incubator where the incubation temperature was raised to 39.5±0.1°C (thermal conditioning) and held constant for 5 h/day from 10.00 am to 3.00 pm through three successive embryonic days (E16, E17 and E18). After each of the daily thermal conditioning eggs were returned back to the main incubator, where they were kept under the optimal incubation conditions.
At the 19th day of incubation, all eggs were transferred to the hatchery and kept till hatching at 37.5±0.1°C and 70% RH.
Postnatal thermal conditioning procedures: After hatching and full feather drying (approximately 8 h post-hatch), chick weight at hatch were measured. The hatchability and late embryonic mortality percentage values were calculated based on the number of eggs for each group. Chicks were maintained under optimal brooding conditions at 34.0°C and 57.7% RH during the first 3 days of age (DOA). On the 4th DOA, 90 chicks from each of the main two groups (T0 and T1) with an overall average weight of 47.16±0.95 g were randomly subdivided into two sub-groups, each of 45 chicks. One of each sub-group (three replicates each) was transferred to a temperature controlled chamber, where they were exposed to a thermal condition at 38.0±0.1°C that was held constant for 5 h/day from 10.00 am to 3.00 pm and 42% RH on each of three consecutive days, the 4th, 5th and 6th DOA. After the end of the thermal conditioning period chicks were daily returned back to the main brooding room. The other sub-group was let under the optimal brooding conditions, resulting finally in four experimental sub-groups (T0T0, T1T0, T0T1 and T1T1).
Chicks’ management: All chicks were let for ad libitum water and feed consumption (FC). They were fed on commercial starter and grower crumbled diets. The starter diet (23% CP and 3020 kcal M.E) was offered during the starter period from day old till 21 DOA, while the grower diet (21% CP and 3100 kcal M.E) was offered until the end of the experiment at 42 DOA.
Birds were exposed to a continuous photoperiod (24 h light) during the 1st week of age (WOA) and then to a light cycle of 18 L: 6 D till the age of six weeks. They were vaccinated during the grow out period against the most common viral diseases (Newcastle, Gamboro and IB).
Collected data: The chicks were individually weighed at hatch and then weekly till the age of six weeks and their live body weights (LBW) were recorded. The body weight gain (BWG), feed consumption (g/bird) and feed conversion ratio (FCR) were calculated.
Different breast muscles (pectorals major and pectorals minor) and carcass yield weights at 42 DOA were recorded and proportionated to LBW at the corresponding age.
At 42 DOA, 15 chicks from each of the four sub-groups were slaughtered by severing the carotid arteries and jugular veins. Blood samples were collected, during their exsanguination in clean heparinized tubes and centrifuged at 4000 rpm for 15 m, the obtained plasma samples were used for the biochemical analyses. The radio immunoassay (RIA) method was used for the determination of triiodothyronine (T3), thyroxine (T4) and insulin like growth factor-I (IGF-I). Plasma T3 and T4 were determined by RIA technique using commercial RIA kits (Immunotech, Beckman Coult. Company) as reported by Britton et al.8. Insulin-like growth factors (IGF) was determined with commercial kits (Gammacoat, kits, clinical assay, Cambridge, Medical Diagnostics, Boston, MA) as reported by Houston and O’Neill9. The eviscerated carcasses were individually weighed and the dressing percentage or carcass yield was recorded. The breast muscles (pectorals major and pectorals minor) were dissected and weighed. All recorded weights were proportionated to the live body weight at the corresponding age.
Histometric and cytometric measurements: At 42 DOA, representative samples (0.5×0.5 cm) from the left major pectoralis muscle of the slaughtered chicks were carefully dissected during the slaughter time and immediately fixed in adequate volume of 10% formalin solution. Thin sections (4-5 micron) were cut and mounted on glass slides (three sections/sample/slide), then stained with the ordinary hematoxylin and eosin stain (H and E) procedures.
Histological observations and cytological measurements: Muscle sections were examined by using a Trinocular light microscope (Labomed, LX 400. Labo America, Inc. USA). Photography shots from the selected specimens were prepared for better illustration of the results. The histomorphometric measurements of myocytes of the pectoral muscles were performed by a software, Image J 1.50b (Wayne Rasband, National Institute of Health, USA) which was calibrated to the micrometer unit (μm) at different magnification powers using a micrometric ruler (PZO-WARS ZAWA- Made in Poland). The myocytes numbers in the pectoral muscle sections were counted in several different fields (3/section) for all treatments, under magnification power of 10X. Large (LD) and small (SD) diameters of 100 myocytes/slide/treatment were recorded to obtain some cytometric and histometric indexes according to Radu-Rusu et al.10 using the following equation:
Where:
| LD | = | Myocytes’ large diameter |
| SD | = | Myocytes’ small diameter |
| π | = | 3.1416 |
Statistical analysis: Data were subjected to a one-way analysis of variance using the general linear model (GLM) procedure11. The differences among means were tested using Duncan’s multiple range test12. All percentages were subjected to arc-sin transformation of the square root before analysis, however, actual percentage means were presented.
RESULTS AND DISCUSSION
The data in Table 1 showed the effect of pre- and postnatal TC on hatchlings body weight and all productive performance traits up to the age of marketing at 42 DOA. The presented data generally illustrated a beneficial effect of pre- and postnatal TC on LBW at 21 and 42 DOA.
It was cleared that, chicks of pre- and postnatal TC (T1T1) had significantly the best LBW, BWG and FCR values, followed by prenatal TC group (T1T0) comparable to group of postnatal TC (T0T1) and non-treated control group (T0T0).
| Table 1: | Effect of pre and post-hatch thermal conditioning on live body weight (LBW), body weight gain (BWG), feed consumption (FC) and feed conversion ratio (FCR) of broiler chicks |
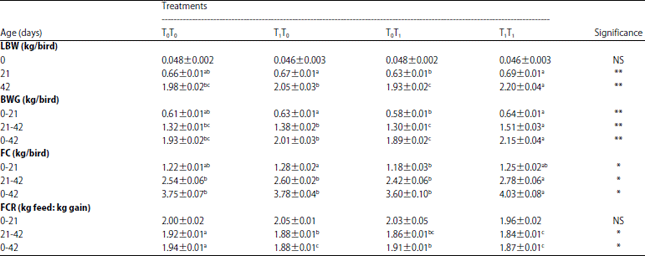 | |
a,b,cMeans within the same row with no common superscript differ significantly. T0T0: Normal incubation and optimal brooding conditions, T1T0: Prenatal thermal conditioning and optimal brooding condition, T0T1: Normal incubation and postnatal thermal conditioning, T1T1: Pre and postnatal thermal conditioning. *p<0.05, **p<0.01, NS: Non-significant | |
However, the T0T1 group had lower LBW and BWG when compared with T0T0 group, it showed better FCR due to its lowest FC.
The results concluded that prenatal TC procedure has a more potent effect than postnatal in enhancing growth performance parameters at a later age. Moreover, the magnitude of this effect was achieved when both procedures were applied (T1T1). This result might plausibly be attributed to better acquisition of heat stress tolerance, throughout the life span of chicks, via stimulating the thermoregulation system. Furthermore, prenatal TC was conducted during a period of development at which myoblasts hyperplasia occurs that may be one of the considered reasons for enhancing body weight and carcass yield thereafter. Additionally, embryonic TC may also, accelerated the metabolic reaction which in turn helped the developed chicks to grow faster. In this respect, Collin et al.13 stated that TC during embryogenesis gave the best results for initiating post-hatching growth when applied for 6-12 h at 39.5°C from E16-E18.
These results are in harmony with Abdel-Fattah and Shourrap3, who found that prenatal TC at 39.0°C for 6 h/day from E16-E18 had significantly enhanced LBW, BWG and FCR at 28 and 42 DOA. However, Willemsen14 concluded that continuous TC at 39.6 or 40.6°C declined body weight from hatch till marketing age. On the other hand, Al-Zhgoul et al.15 illustrated that exposing hatching Hubbard eggs to 38.8°C for 6 or 18 h/day from E10-E18 had no significant effect on body weight for the first 4 WOA.
The observed reduction in LBW and BWG associated with T0T1 group was in full agreement with the finding of Elsayed et al.16, who showed that post-hatch TC of Cobb and Ross strains at 36-37°C or 38-39°C for 6 h/day at the 1st, 3rd or 5th DOA reduced body weight gain during the first WOA.
The slightly observed growth retardation due to early post-hatch thermal challenge (T0T1), compared with T1T0, was associated with reduced enterocyte proliferation after heat exposure during the 1st week of growth as was previously suggested17. However a period of growth compensation was achieved, thus regained the weight of chicks and this is highly correlated with better nutrient utilization and absorption6.
The effects of pre- and post-hatch thermal conditioning on the relative weight of the carcass and breast muscles were shown in Table 2. It was obvious that, chicks subjected to both pre- and postnatal TC treatment (T1T1) had the highest (p<0.01) dressing relative weights at 42 DOA followed by the T1T0 group compared with the other groups (T0T0 and T0T1).
A similar trend was nearly achieved for the relative weight of breast muscles, though the superiority was accomplished for the T1T0 then for the T1T1, with lack of significance, compared with the optimally incubated chicks (T0T0), while intermediate values were recorded for chicks of T0T1 treatment.
The current findings were coincided with Loyau et al.18, they indicated that TC at 39.5°C for 12 h/day from E7-E16 significantly enhanced breast muscle yield at 35 DOA. However, Willemsen14 found no significant changes in the relative weights of breast muscle of four day old broiler chicks hatched from TC eggs at 40.6 or 39.6°C during E16-E18.
| Table 2: | Effect of pre- and post-hatch thermal conditioning on percentages of dressing and breast muscles relative weight of broiler chicks at 42 DOA |
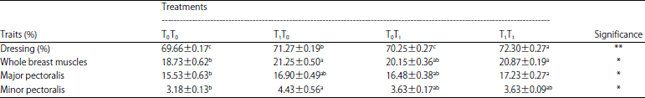 | |
a,b,cMeans within the same row with no common superscript differ significantly. T0T0: Normal incubation and optimal brooding conditions, T1T0: Prenatal thermal conditioning and optimal brooding condition, T0T1: Normal incubation and postnatal thermal conditioning, T1T1: Pre and postnatal thermal conditioning. *p<0.05, **p<0.01 | |
| Table 3: | Effect of pre- and post-hatch thermal conditioning on plasma T3, T4 and IGF-I levels at 42 DOA |
 | |
a,bMeans within the same row with no common superscript differ significantly. T0T0: Normal incubation and optimal brooding conditions, T1T0: Prenatal thermal conditioning and optimal brooding condition, T0T1: Normal incubation and postnatal thermal conditioning, T1T1: Pre and postnatal thermal conditioning. *p<0.05, **p<0.01, NS: Non-significant | |
It’s noteworthy that, these favorable results achieved in carcass trait values could be account for the growth acceleration associated with TC, particularly the prenatal TC (Table 1).
Plasma concentration of metabolic hormones as affected by pre or postnatal TC was illustrated in Table 3. Data showed insignificant differences among all groups concerning the T3 level, while T4 level was elevated (p<0.05) for chicks of optimal incubation conditions (T0T0), followed by those of either single pre (T1T0) or post-hatch (T0T1) TC, compared with chicks hatched from both embryonic and early age TC (T1T1). It was well known, that thyroid gland hormones are important in heat stress adaptation through their crucial role in regulating the metabolic rate. Thus, one of the possible mechanisms of thermotolerance was the ability to reduce heat production that is regulated by thyroid hormones secretion and activity, in particular T3. Consequently, the anticipated induction of thermotolerance through thermal conditioning either pre or postnatal will be associated with the modulation of thyroid hormone activities.
In this respect, Al-Zhgoul et al.15 reported that raising the ambient temperature to 41.0°C for 6 h/day at either 3 or 7 DOA, caused a significant reduction in plasma T3 levels at 42 DOA, when Hubbard eggs were exposed to 38.8°C for 6 or 18 h/day from E10-E18 compared with chicks hatched from eggs of normal incubation. Similarly, T3 level was decreased significantly at 42 DOA in Cobb broiler groups of post-hatch TC and pre with post-hatch TC compared to control and pre hatch TC ones19.
Also, Arjona et al.20 found no significant effect of post-hatch TC at 35.0-37.8°C for 24 h at 5 DOA on the level of both T3 and T4 when chicks were exposed to heat stress at 35-37.8°C at 43 DOA.
It could be suggested that, decreased plasma T4 levels, may be pointed out that broiler chicks not only respond to hyperthermia by increasing evaporative cooling through panting, but also decreased their metabolism and thereby heat production.
The worst (p<0.01) concentration of plasma IGF-I level was attained for chicks of non-treated control group (T0T0) compared to the remaining TC groups, whether pre or postnatal (Table 3).
The superiority of TC groups in achieving better marketing LBW, when compared with the non-conditioned group was associated with increased insulin-like growth factor-I (IGF-I) level and its immediate stimulation of satellite cell myogenic processes, as well as the enhanced enterocyte proliferation, expression and activity of brush-border membrane enzymes21.
Therefore, it is postulated that either embryonic or early age heat modulation and subsequently acclimation could help birds to neutralize hot summer stress resulted in a predictable upward shift of thyroid gland activity, which was reflected in regulating IGF-I.
In this respect, Christensen et al.22 found that the control line of growth selected turkey embryos had elevated IGF-I concentrations in response to higher temperatures at 37.2°C during the last three days of incubation.
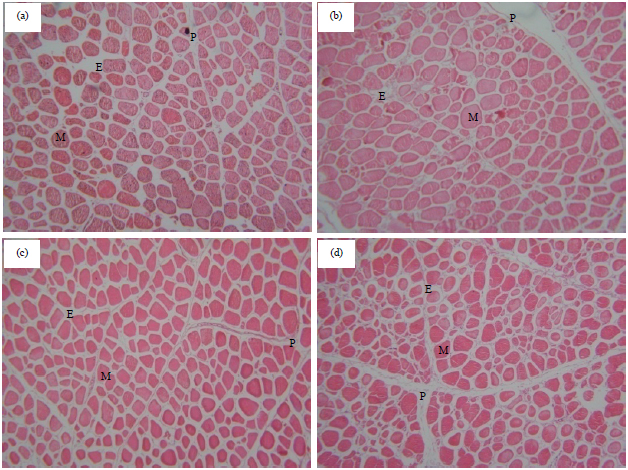 | |
| Fig. 1(a-d): | Transverse section through breast muscle from birds of different thermal treatments at 42 DOA, (a) T0T0, (b) T0T1, (c) T1T0 and T1T1. P: Perimysium, E: Endomysium, M: Myocytes (H and E 100X) |
T0T0: Normal incubation and optimal brooding conditions, T1T0: Prenatal thermal conditioning and optimal brooding condition, T0T1: Normal incubation and postnatal thermal conditioning, T1T1: Pre and postnatal thermal conditioning | |
| Table 4: | Effect of pre and post-hatch thermal conditioning on major pectoralis muscle myocytes numbers, thickness and cross-section area of the myocytes at 42 DOA |
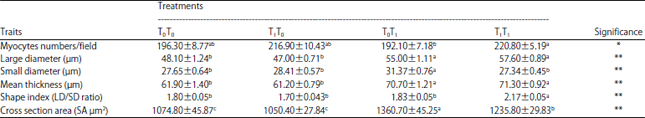 | |
a,b,cMeans within the same row with no common superscript differ significantly. T0T0: Normal incubation and optimal brooding conditions, T1T0: Prenatal thermal conditioning and optimal brooding condition, T0T1: Normal incubation and postnatal thermal conditioning, T1T1: Pre and postnatal thermal conditioning. *p<0.05, **p<0.01 | |
Moreover, Piestun et al.23 showed that raising the incubation temperature of Cobb broiler eggs to 39.5°C (with an increase of 1.7°C from normal conditions) at E16-E18 for 3 or 6 h/day increased IGF-I levels in muscles of the thermally manipulated groups than in control one.
The microscopic structure of major pectoralis muscles from different treatment groups were shown in Fig. 1. From the histological observation, it is noteworthy that, sections from different treatments showed typical skeletal muscle structure. Thereby, inspecting and measuring cytological differences was a better procedure to distinguish between different experimental groups.
The cytological measurements of the major pectoralis form chicks of different thermal conditioning treatments at 42 DOA were illustrated in Table 4. Results showed that prenatal heat conditioning either singly (T1T0) or in combining with the postnatal (T1T1) had significantly increased the myocytes number of major pectoralis muscles/field, 216.90 and 220.75, respectively, vs 192.1 and 196.3 for the postnatal thermal conditioning (T0T1) and non-treated groups (T0T0), respectively.
Concerning the myocytes dimensions (large and small diameters), thickness (Dχ) and cross section area (SA), the current estimated results revealed that chicks exposed to postnatal heat challenge (T0T1) had the highest (p<0.0) values, followed by those of T1T1 group compared to the other two groups, particularly the T0T0 one. Therefore, no treatment was higher than the T1T1 in the values of myocytes shape index (SI).
It is well known that the muscle fibers number, size and fiber-type composition are closely related to each other24. The performance of muscle in the adult animal largely depends on the muscle fiber number and type and therefore on fiber size. The muscle weight is a function of the total number of myofibers, myofiber cross-section area and length25.
From the histological observation and cytological measurements of pectoralis muscles, it was clear that TC conducted either during the late stage of embryonic development or at the early age post-hatch increased the myocytes amount of broiler pectoral muscles at the marketing age. In this respect, previous studies declared that TC during E16-E18 stimulated myonuclei proliferation consequently, increased the numbers of breast myonuclei of hatchling broiler chicks. This myonuclei hyperplasia resulted in a hypertrophy of pectoralis muscles by increasing the numbers of muscle fibers (myocytes). Thereby, increasing the breast muscles yield as accomplished herein at the marketing age3. In a similar manner, Werner and Wicke26 showed that raising the incubation temperature beyond the normal range might either directly influence molecular steps necessary for the proliferation and differentiation of myoblasts or indirectly affects myogenesis by general stimulation of the metabolism and the in ovo movement of the embryo. Likewise, exposure of late-term embryos to high temperature, 38.5 or 39.5°C for 3 or 6 h/day improved both diameter of myofibers and myoblast proliferation thus, enhanced absolute muscle growth at marketing1,13,23.
Halevy et al.2 postulated that post-hatch thermal conditioning caused an increase in satellite cell proliferation, necessary for further muscle hypertrophy. This was accompanied by a marked induction of insulin-like growth factor-I (IGF-I), but not hepatocyte growth factor in the breast muscle. Moreover, earlier reports affirmed that post-hatch TC of chicks at an early age results in transient growth arrest, followed by immediate compensatory growth. This leads to higher body and breast muscle weights of the TC chicks vs. their untreated counterparts at later ages27,28, which is in close agreement with the present findings of growth and carcass results (Table 1, 2).
CONCLUSION
It could be concluded that pre and/or postnatal TC treatments are a practical application to enhance productive performance and muscle growth of broiler chicks. Moreover, superior benefit could be achieved by the combination of both procedures, pre and post-hatch thermal conditioning as established for the T1T1 group.
SIGNIFICANCE STATEMENT
The present study discovers the possible synergistic effect of pre and postnatal thermal conditioning on the metabolic, hormonal makeup of broiler chicks and in enhancing their muscular proliferation. This study will help researchers to uncover the possible role of pre and postnatal thermal conditioning, particularly their combination to stimulate the thermotolerance acquisition as well as muscular accretion of broiler chicks.
REFERENCES
- Halevy, O., S. Yahav and I. Rozenboim, 2006. Enhancement of meat production by environmental manipulations in embryo and young broilers. World's Poult. Sci. J., 62: 485-497.
CrossRefDirect Link - Halevy, O., A. Krispin, Y. Leshem, J.P. McMurtry and S. Yahav, 2001. Early-age heat exposure affects skeletal muscle satellite cell proliferation and differentiation in chicks. Am. J. Physiol. Regul. Integr. Comp. Physiol., 281: R302-R309.
CrossRefPubMedDirect Link - Piestun, Y., S. Druyan, J. Brake and S. Yahav, 2013. Thermal manipulations during broiler incubation alter performance of broilers to 70 days of age. Poult. Sci., 92: 1155-1163.
CrossRefDirect Link - Tan, G.Y., L. Yang, Y.Q. Fu, J.H. Feng and M.H. Zhang, 2010. Effects of different acute high ambient temperatures on function of hepatic mitochondrial respiration, antioxidative enzymes and oxidative injury in broiler chickens. Poult. Sci., 89: 115-122.
CrossRefDirect Link - Fernandes, J.I.M., L.B. Scapini, E.T. Gottardo, A.M. Burin Junior, F.E.D.S. Marques and L. Gruchouskei, 2013. Thermal conditioning during the first week on performance, heart morphology and carcass yield of broilers submitted to heat stress. Acta Scient. Anim. Sci., 35: 311-319.
CrossRefDirect Link - Abd El Azim, A.M., 2012. Improve the heat tolerance of broilers through heat treatment during the first two weeks. Egypt. Poult. Sci., 32: 483-495.
Direct Link - Britton, K.E., V. Quinn, B.L. Brown and R.P. Ekins, 1975. A strategy for thyroid function tests. Br. Med. J., 3: 350-352.
CrossRefDirect Link - Houston, B. and I.E. O'Neill, 1991. Insulin and growth hormone act synergistically to stimulate insulin-like growth factor-I production by cultured chicken hepatocytes. J. Endocrinol., 128: 389-393.
CrossRefPubMedDirect Link - Collin, A., M. Picard and S. Yahav, 2005. The effect of duration of thermal manipulation during broiler chick embryogenesis on body weight and body temperature of post-hatched chicks. Anim. Res., 54: 105-111.
CrossRefDirect Link - Al-Zhgoul, M.B., A.E.S. Dalab, M.M. Ababneh, K.I. Jawasreh, K.A.A. Busadah and Z.B. Ismail, 2013. Thermal manipulation during chicken embryogenesis results in enhanced Hsp70 gene expression and the acquisition of thermotolerance. Res. Vet. Sci., 95: 502-507.
CrossRefDirect Link - Elsayed, N.A.M., A.E. Elkomy, Amina, S. El-Saadany and E.Y. Hassan, 2009. New suggested schemes for incubation temperature and their effect on embryonic development and hatching power. Asian J. Poult. Sci., 3: 19-29.
CrossRefDirect Link - Uni, Z., O. Gal-Garber, A. Geyra, D. Sklan and S. Yahav, 2001. Changes in growth and function of chick small intestine epithelium due to early thermal conditioning. Poult. Sci., 80: 438-445.
CrossRefDirect Link - Loyau, T., C. Berri, L. Bedrani, S. Metayer-Coustard and C. Praud et al., 2013. Thermal manipulation of the embryo modifies the physiology and body composition of broiler chickens reared in floor pens without affecting breast meat processing quality. J. Anim. Sci., 91: 3674-3685.
CrossRefDirect Link - Tona, K., O. Onagbesan, V. Bruggeman, V. Collin and C. Berri et al., 2008. Effects of heat conditioning at d 16 to 18 of incubation or during early broiler rearing on embryo physiology, post-hatch growth performance and heat tolerance. Arch. Geflugelk, 72: 75-83.
Direct Link - Arjona, A.A., D.M. Denbow and J.W. Weaver Jr., 1990. Neonatally-induced thermotolerance: Physiological responses. Compa. Biochem. Physiol. Part A: Physiol., 9: 393-399.
CrossRefPubMedDirect Link - Lin, H., H.C. Jiao, J. Buyse and E. Decuypere, 2006. Strategies for preventing heat stress in poultry. World Poult. Sci. J., 62: 71-86.
CrossRefDirect Link - Christensen, V.L., J.P. McMurtry, W.E. Donaldson and K.E. Nestor, 2001. Incubation temperature affects plasma insulin-like growth factors in embryos from selected lines of turkeys. Poult. Sci., 80: 949-954.
CrossRefDirect Link - Piestun, Y., M. Harel, M. Barak, S. Yahav and O. Halevy, 2009. Thermal manipulations in late-term chick embryos have immediate and longer term effects on myoblast proliferation and skeletal muscle hypertrophy. J. Applied Physiol., 106: 233-240.
CrossRefPubMedDirect Link - Ryu, Y.C., M.C. Rhee and B.C. Kim, 2004. Estimation of correlation coefficients between histological parameters and carcass traits of pig longissimus dorsi muscle. Asian-Aust. J. Anim. Sci., 17: 428-433.
CrossRefDirect Link - Werner, C. and M. Wicke, 2008. Increasing the temperature at a certain period during the incubation of broiler eggs improves the carcass and breast yield without a negative impact on the meat quality. Arch. Tierz. Dummerstorf, 51: 11-16.
Direct Link - Yahav, S. and S. Hurwitz, 1996. Induction of thermotolerance in male broiler chickens by temperature conditioning at an early age. Poult. Sci., 75: 402-406.
CrossRefPubMedDirect Link