
George John
Department of Zoology, Periyar E.V.R. College (Autonomous), 620 023 Tiruchirappalli, Tamil Nadu, India
Razack Rafi Mohamed
Department of Zoology, C. Abdul Hakeem College (Autonomous), Melvisharam, Tamil Nadu, India
Pachaan Kolanchinathan
Department of Zoology, Periyar E.V.R. College (Autonomous), 620 023 Tiruchirappalli, Tamil Nadu, India
LiveDNA: 91.22434
Athmanathan Balasundaram
Department of Zoology, Periyar E.V.R. College (Autonomous), 620 023 Tiruchirappalli, Tamil Nadu, India
LiveDNA: 91.16997
Asian Journal of Animal and Veterinary Advances
Year: 2018 | Volume: 13 | Issue: 3 | Page No.: 282-294







ABSTRACT
Background and Objective: Formulation of probiotic mixtures for marine shrimps are still not perfect, the concept of probiotics is totally unfamiliar for freshwater-cultured prawns. This study was an attempt to evaluate the nutritional efficacy of two putative probiotic strains Bacillus subtilis RCMB21 and Pseudomonas fluorescens RCMB39. Materials and Methods: Bacillus subtilis RCMB21 and Pseudomonas fluorescens RCMB39 were separately in a commercial formulated feed for shrimps. (1) Coating the feed, with agar as the binder and (2) Fermentation of the feed with bacteria. Feeding experiments were conducted for 60 days on Macrobrachium malcolmsonii (M. malcolmsonii) with probiotic incorporated diet and growth and nutritional quality improvement of the prawn were assessed. Data were analyzed using two-way analysis of variance (ANOVA) using SPSS. Results: Bacillus subtilis coated diet, the increased weight was 3.5 times over the initial and 2.7 times for Pseudomonas fluorescens coated feed at the end of 60 days. Similar results were obtained for protein, carbohydrate and lipid. Electrophoresis of muscle showed dense intense polypeptide fractions of 200, 70, 40 and 20 kD. Fatty acid profile of prawns showed 47.43% saturated fatty acids, 13.71% monoenes and 38.8% of polyunsaturated fatty acids. Conclusion: Based on growth and biochemical studies, it could be assumed that bacteria coated diets performed better over diets fermented with bacteria. Between the bacteria incorporated, Bacillus subtilis had more beneficial properties.
PDF Abstract XML References Citation
Received: October 15, 2017;
Accepted: January 04, 2018;
Published: April 15, 2018
How to cite this article
George John, Razack Rafi Mohamed, Pachaan Kolanchinathan and Athmanathan Balasundaram, 2018. Nutritional Value of Two Bacterial Strains Bacillus subtilis RCMB21 and Pseudomonas fluorescens RCMB39 as Feed Supplement for Freshwater Prawn Macrobrachium malcolmsonii. Asian Journal of Animal and Veterinary Advances, 13: 282-294.
DOI: 10.3923/ajava.2018.282.294
URL: https://scialert.net/abstract/?doi=ajava.2018.282.294
DOI: 10.3923/ajava.2018.282.294
URL: https://scialert.net/abstract/?doi=ajava.2018.282.294
INTRODUCTION
The development of probiotics applicable to cultured organisms involves a multi-step process in which efficiency evaluation experiments are of fundamental importance. Rather than depending on the positive traits of candidate bacteria alone the use of target organism itself in the evaluation procedure provides a stronger basis for in vivo applications. One of the recognized modes of action of probiotics is providing positive growth factors1.
In addition to the obvious probiotic traits like antipathogenic nature, bacterial role in nutrition as SCPs has long been highlighted by scientists involved in shrimp and fish nutrition research2-5. Besides whole cells, bacterial spores are also put into use as food additives6. Nutritive value of two bacterial strains, Brevibacterium lactofermentum and Corynebacterium glutamaticum was assessed by Kiessling and Askbrandt7 using them as partial replacement for fish meal in the diet of rainbow trout Oncorhyncus mykiss. Substantial replacement of micro-algae in the larval diet of Penaeus monodon with the marine heterotrophic bacteria, Bacillus (2 strains), Pseudomonas (2 strains) Moraxella and Micrococcus, was successfully carried out by Mohamed3.
Instead of using as a direct feed for the cultured species, bacteria are also used for enriching live feed. Intriago and Jones8 used Flexibacter sp. as an exclusive diet for Artemia. The rotifer, Brachionus plicatilis has also been reared using bacteria as their feed6,9. Oral administration of probiotics led to their adherence in shrimp digestive tract and enhancement on the growth, survival, biochemical parameters, immune expression and disease resistance in Penaeus monodon was studied by Kolanchinathan et al.10.
With a view to assess the benefits of the two bacterial strains, Bacillus subtilis-RCMB21 and Pseudomonas fluorescens-RCMB39 as nutritional probiotics, they were incorporated in a commercial prawn feed and were fed the juveniles of M. malcolmsonii for 60 days. Hence, this study was carried out to assess the nutritional augmentation of chosen probionts and selection of suitable mode of entry into the animals.
MATERIALS AND METHODS
Experimental animal and design: Juveniles of M. malcolmsonii measuring from 7.0-9.5 cm were collected from the Lower Anaicut (Anaikarai) region of the river Coleroon and acclimated to the laboratory condition in Periyar E.V.R. College (Autonomous), Tiruchirappalli-620 023, Tamil Nadu, India, (2013-2015). The animals were washed in 0.1% benzalkonium chloride solution and kept in sufficient quantity of sterile water. For sterilization, water was purified in a commercial water purifier which had a physical filter assembly, activated charcoal column and a U.V irradiation unit. The juveniles were starved for 2 days in sterile water prior to the feeding schedule, with the intention of removing majority of bacteria from the gut of the animal.
The experimental animals were divided into 6 groups, comprising of 25 animals each. Animals of the first group were fed with reconstituted commercial feed (‘control’) and those of the second group with agar coated (‘control coated’). Animals of the third group were fed with feed pellets coated with Bacillus subtilis RCMB21 and the ones of the fourth group, with feed fermented with B. subtilis RCMB21. Similarly prawn juveniles of the fifth group were maintained on feed pellets coated with Pseudomonas fluorescens-RCMB39, while animals of the sixth group were maintained on feed fermented with P. fluorescens. Daily feed rationing was done at 3% b.wt., of animals in each tub: In two doses, one at 10 h and the second at 18 h. Feed not consumed if any and the excreted materials were siphoned out in the morning daily, prior to water replenishing. Total duration of the feeding experiments was 60 days. Animals in the tubs were frequently observed for any change in their swimming behaviour, colouration, moulting frequency and the occurrence of mortality.
At the end of each 15 days period, 5 animals from each tub were selected at random and were subjected to body measurements and biochemical analysis. Fatty acid composition of the muscles of prawn was analyzed only on the 45th day of the experiment. Electrophoresis of muscle and hepatopancreas proteins was done at the culmination of 15 and 30 days of feeding.
Probiotic bacteria: Putative probiotic strains of bacteria Bacillus subtilis RCMB21 and Pseudomonas fluorescens RCMB39 were isolated from the gut of wild freshwater prawn M. malcolmsonii. Based on the positive results of the screening procedures, Bacillus subtilis RCMB21 and Pseudomonas fluorescens RCMB39 was selected and were aseptically grown on 10 mL aliquots of nutrient broth (18 h at room temperature: 28±2°C). Five milliliters each of the culture broth was transferred into 250 mL nutrient broth and incubated on a rotary shaker at 150 rpm for 18 h at room temperature. The cells were harvested by centrifugation (Remi cooling microfuge, at 10,000 rpm for 15 min at 4°C). The microbial biomass was diluted with 0.85% NaCl to attain a final concentration of 0.150 g mL–1 of bacterial cells . The bacteria saline mixture was stored at 4°C and within 24 h period they were incorporated in a commercial prawn feed.
Scanning probe microscope (SPM) studies were conducted to observe the morphology of Bacillus subtilis RCMB21 and Pseudomonas fluorescens RCMB39. Freshly harvested bacteria were washed in distilled water. Samples for SPM analysis were transferred on to cover glass and mounted on specimen stubs using double adhesive tape. All analyses were made using Shimadzu scanning probe microscope model SPM 9500 using a 30ja×30ji scanner in contact mode.
Feed formulations|
Control: Commercial prawn feed (Naza Ltd., Philippines), which is widely used in Tamil Nadu for farming the marine shrimp, Penaeus monodon and the freshwater giant prawn Macrobrachium rosenbergii (Scampi) was used as the control diet, feed of starter grade was autoclaved to remove the existing bacteria (if any), incorporated 2% (w/w) cassava as binder and was reconstituted into pellets with a 1 mm diameter extruder. The pellets were oven dried at and stored in sterile air tight containers.
Bacteria coated feed: Bacillus subtilis RCMB21 and Pseudomonas fluorescens RCMB39 cells were incorporated separately at 3% weight of the feed. Twenty one milliliters of the stored bacterial cell-saline mixture was centrifuged at 10,000 rpm for 15 min at 4°C. Hundred grams of the re-pelleted feed was warmed up to 45°C. The bacterial biomass was mixed thoroughly in molten agar (35°C) and blended with the pre-warmed feed pellets using sterile glass rods to make a uniform coating over the pellets. The pellets were cooled to room temperature and stored in airtight containers, at 4°C for 4-5 days. On every 4th day, a fresh batch of bacteria-coated feed was prepared following the same procedure.
Feed fermented with bacteria: The re-pelleted feed was fermented with Bacillus subtilis RCMB 21 and Pseudomonas fluorescens RCMB39 separately, following the procedure by Ramesh and Lonsane11, suggested for alpha amylase production with Bacillus megaterium. Necessary modifications were however, made to suit the changed substratum.
Known quantity of the commercial feed was made moist and the pH was adjusted to 8.0. The feed was autoclaved for 60 min and cooled down to 37°C. Bacillus subtilis RCMB21 and P. fluorescens RCMB39 stored at 4°C were brought to room temperature and mixed thoroughly with the warm feed taken in a beaker kept on a water bath at 37°C. The mouth of the beaker was covered with tinfoil with a provision for inserting a sterile glass rod for periodic mixing. The feed-bacteria mixture was moistened at 24 h interval with sterilized water, to maintain a steady fermentation action. The fermentation assembly was maintained for 48-72 h period, depending upon the intensity of the vinegarish smell emanating.
At the termination of the fermentation period, the feed-bacterial mixture was mixed with 2% w/w cassava (binder) and squeezed through the diets of an extruder to get pellets of 1 mm diameter. The fermented feed pellets were dried in an oven 60°C for 24 h and stored in airtight containers. Each bacterium was administered at 3% weight of the feed, initially.
Growth parameters: Several growth parameters such as weight gain (%), length gain (%), specific growth rate (SGR) and feed efficiency were calculated for M. malcolmsonii fed on formulated diets supplemented with following chosen probiotic bacterial species, either 1) B. subtilis 2) P. fluorescens :

Where:
| W1 | = | Weight at time T1 |
| W2 | = | Weight at time T2 |
Feed efficiency (FE) by Kiessling and Askbrandt7:
![]()
Biochemical analysis: Biochemical analysis were carried out in bacteria incorporated feeds and in the abdominal muscles determined by moisture12, ash12, total protein13, carbohydrate14 and lipid15 of M. malcolmsonii, at various feeding stages.
Fatty acid composition: Muscle tissue of prawns was rapidly frozen at -25°C in a deep freezer, until analyzed. Then, 1.5 g of the tissue was homogenized and the sample was trans-esterified using a mixture of acetyl chloride and methanol (1/20 v/v). The resulting fatty acid methyl esters (FAME) were then injected on capillary column (25 mL×0.32 mm i.d.) employing a flame ionization detector (FID) in a Hewlett Packard 5890 gas chromatograph. The reported fatty acid compositions were based on the average of two injections and were expressed as percentage (chromatogram area) of total fatty acids.
Electrophoretic protein profiling: Protein profiling was performed using SDS-polyacrylamide gel electrophoresis as described by Laemmli16. Linear slab polyacrylamide gels of 12% were used for the analysis of peptides in hepatopancreas and abdominal muscles of M. malcolmsonii fed on bacteria incorporated diets till the 15th and 30th day of experiment. Electrophoresis was carried out for about 4 h at a constant current of 100 V. Molecular weight standards (MBI Fermentas: Protein molecular weight markers No. SM0431) of (3-Galactosidase (116.0 kD), bovine serum albumin (66.2 kD), ovalbumin (45.0 kD), lactate dehydrogenase (35.0 kD), restriction endonuclease BSP981 (25.0 kD), p-lactoglobulin (18.4 kD) and lysozyme (14.4 kD) were run concurrently. After electrophoresis, the gels were stained for 24 h with 0.2% Coomassie Brilliant Blue R Solution and de-stained by a de-stainer containing methanol, acetic acid and water (7:50:63 v:v:v). Densitometric quantification of the bands was also carried out.
Statistical analysis: All the reported values were expressed as mean±standard deviation. The statistical significance was evaluated by two-way analysis of variance (ANOVA) using SPSS version 10.5 (SPSS, Cary, NC, USA) and the individual comparisons were obtained by Post hoc analysis ‘Student Newman Keuls“ test (SNK), a significance level of p<0.05 was used17.
RESULTS
Bacteria: Scanning photomicrograph studies revealed the morphology of the bacteria B. subtilis-RCMB21 and P. fluorescens-RCMB39. Laser scanned magnified images of the bacteria are given in Fig. 1a, b. Rod shaped nature with smooth surfaces of their body wall were evident in the pictures.
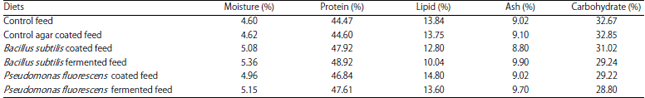
Biochemical composition of feeds: The results of the proximate composition analysis for the control and bacteria incorporated feeds are presented in Table 1.
The moisture content varied between 4.6-5.36% of the formulated feed. Control feed had lesser moisture value compared to the rest. Moisture content of the bacteria-coated feed was analyzed, prior to refrigeration. Percentage of ash varied between 8.8 and 9.9%. Any defined pattern in variation was not apparent.
Protein content of the feeds varied between 44.47 and 48.92%. Bacteria incorporated feeds, both B. subtilis and P. fluorescens, had higher percentage of proteins than the control and agar-coated feeds. Fermented feeds had an edge in their protein content over the corresponding bacteria-coated feeds (Table 1).
Control feed and agar coated feed had 13.84 and 13.75% lipids, respectively. Pseudomonas fluorescens-coated feed had 14.8% lipid while the feed fermented by the same bacteria had slightly lesser level of lipids. Lipid content of the B. subtilis-coated feed was 12.8% only while the B. subtilis-fermented feed had least amount of lipid (10.04%).
Carbohydrates were prominent component of the feeds. ‘Control’, ‘control agar coated’ and the ‘B. subtilis-coated’ feeds had high amounts of carbohydrates (31.02-32.67%). Bacillus subtilis-fermented feed and the feeds, coated and fermented with P. fluorescens had lesser amount of carbohydrates (Table 1).
Physical growth of prawns fed with bacteria-incorporated diets
Length-increase: There was a steady progress in body length for all the groups of animals, fed on diverse diets formulated for this study.
| Table 1: | Proximate composition of bacteria incorporated diets (% of dry matters) |
 | |
 | |
| Fig. 1(a-b): | (a) Bacillus subtilis RCMB21 (SPM) and (b) Pseudomonas fluorescence RCMB39 (SPM) |
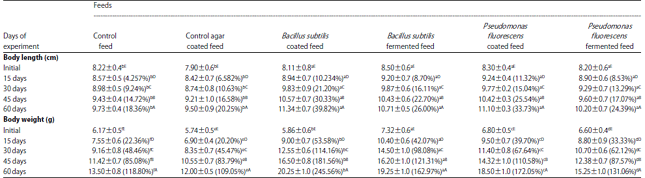
The animals which had an initial average length of 8.22 cm, when fed on the control (normal) feed, showed an increase of 9.73 cm at the end of the 60th day-an increase of 18.37% over the initial length (Table 2). The prawn juveniles fed on agar coated normal feed, also showed a close similar rate of length increase (20.25% increase over the initial). Compared to these the length-increase in animal groups fed on either B. subtilis or P. fluorescens coated feeds was more. The animals, which were initially 8.11 and 8.3 cm on the average, could grow in the end to average lengths of 11.34 and 11.1 cm, respectively. At the end of 60 days, animals fed on B. subtilis coated feed registered length increase of 39.82%, while the ones fed on P. fluorescens coated feed showed a length increase of 33.73%. Although, not up to the extent of coated feeds, bacteria-fermented feeds also could elicit good growth response. Prawns fed on B. subtilis fermented feeds showed an increase of 26% increase in length while those fed on P. fluorescens-fermented feed could gain in length by 24.39% (Table 2).
Two-way ANOVA and SNK tests showed that there were significant differences in length increase between the control groups (normal and normal-agar coated diet fed) and the animal groups fed on bacteria coated as well fermented diets.
| Table 2: | Body length and body weight variation of M. malcolmsonii fed on bacteria incorporated feeds |
 | |
ANOVA (p<0.05), SNK test: Identical lower case superscripts denote similar values vertically, Identical upper case superscripts denote similar values horizontally | |
With respect to length-increase, there was no significant difference among the bacteria-incorporated diet fed groups. The progression of growth in terms of body length for each 15 days segment of the 60 days experiment duration was also significantly distinct for each diet group as evidenced by the five significantly different harmonic mean subsets corresponding to each experiment day-segment.
Weight increase
Gross body weight: For the prawns fed on normal feed and normal-agar coated feed, body mass increase was less compared to the prawns fed on bacteria incorporated diets. At the end of 60 days, the prawns that got ‘control’ feed attained 118.8% increase in body weight, while those which were fed on agar coated normal feed could register only 109.05% growth (Table 2). Body mass increase in B. subtilis coated diet fed prawns was immense. Animals of 5.86 g average body weight could grow to body weight of 20.25 g (245.56%) over the initial weight. Body weight increase in P. fluorescens coated diet fed prawns was apparently to a lesser extent (from 6.8 g initial to a final weight of 18.5g-172.05%). Among the fermented feeds B. subtilis fermented feed could elicit greater body mass increase (7.32 g initial weight to 19.25 g final weight-162.98%) in prawns, than P. fluorescens fermented feed (6.6 g initial weight to 15.25 g final weight-131.06%) increase over the initial weight). Results of ANOVA and SNK tests also revealed the same trend. Prawn-body weight increase was distinctly different for each kind of feed. The growth rate in the order of increase was: Agar coated diet, normal diet P. fluorescens fermented diet, P. fluorescens coated diet, B. subtilis coated diet and B. subtilis fermented diet. Significantly different harmonic mean subsets of SNK table revealed substantial rate of weight increase for all the diet-groups with respect to the progression of experiment duration. The magnitude of weight increase as calculated for a unit animal 1 g b.wt.-initial). At the end of the 60th day of experiment, body weight was doubled in the two ‘control-diet’ groups (normal and agar coated feeds), while increase was 3.5 times for the prawns fed on B. subtilis coated feed and 2.7 times for those fed on P. fluorescens coated feed. Fermented feeds could bring forth slightly lesser degree of growth than the bacteria coated feeds (2.63 times weight increase for B. subtilis fermented feed and 2.3 times for P. fluorescens fermented feed).
Dry weight: The percentage dry weight values of the muscles of prawns fed on various bacteria combined diets are presented in Table 3. The dry matter percentage showed a range of 22.5-28.5%.
Specific growth rate (SGR): Specific growth rate (SGR) in terms of body weight increase, for prawns fed on various diets is presented in Table 3. From Table 3, it was evident that the SGRs were more or less uniform for the ‘control’ and ‘agar coated feeds’ for most of the experimental segments. Bacteria incorporated diets (coated, fermented feeds) had comparatively higher SGRs at least for the initial 30 days of experiment, after which the indices seemed to level off.
| Table 3: | Dry weight of muscle, specific growth rate (SGR) and Feed efficiency of the M. malcolmsonii fed on bacterial incorporated feeds |
 | |
| Table 4: | Biochemical variation of tissue-protein, carbohydrate and lipids of M. malcolmsonii fed on bacteria incorporated feeds |
 | |
ANOVA (p<0.05), SNK test: Identical lower case superscripts denote similar values vertically, Identical upper case superscripts denote similar values horizontally | |
Feed efficiency (FE): Indices depicting the efficiency of various feeds, in terms of their conversion as prawn body mass are presented in Table 3. Though there were un-ascribable fluctuations, maximum efficiency could be assigned to B. subtilis coated feed followed by P. fluorescens coated feed. Normal feed and the feeds fermented by B. subtilis and P. fluorescens were further down in their efficiency, while the agar coated normal feed was the least efficient, comparatively.
Growth related biochemical variations
Protein
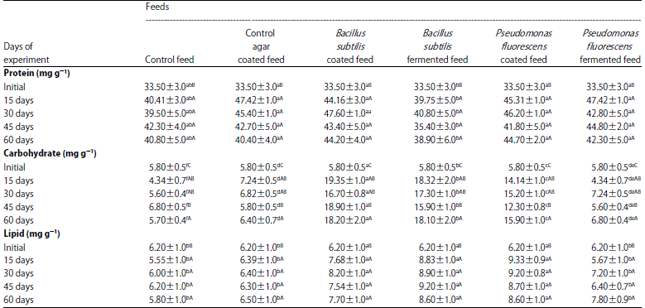
Soluble protein: The TCA soluble protein fractions of the abdominal muscles of M. malcolmsonii, fed on bacteria incorporated diets are presented in Table 4. The values are presented as mg/100 mg muscle dry weight. Though there was a definite increasing trend in the protein values in the first half of the experiment (till 30 days), the values become more or less steady towards the latter half the range of protein values was 35.5-47.60 mg g–1. Protein increase in the prawns was evident, when the values were assigned to the ‘unit animal’ of 1 g initial weight. The increase in prawn weight could be assigned significantly to the buildup of abdominal muscles and protein being the chief constituent of muscles, should also be on the ascent. As evidenced from the increase in protein content was about 5 fold for B. subtilis-coated diet fed prawns and about 4.5 times for P. fluorescens-coated diet fed prawns. For fermented diet fed prawns, the increase was 3.4 times. For the ‘control fed’ groups the increase was approximately 2.7 times.
Analysis of variance and the post hoc tests revealed that the feeds coated with B. subtilis and P. fluorescens elicited maximum increase in protein, while the feeds fermented with the same bacteria, were effective to a lesser extent. Table 4 revealed that the first 15 days of the feeding experiment could bring about maximum extent of protein increase, followed by the next 15 days and then the rest of the experiment period, in a descending order.
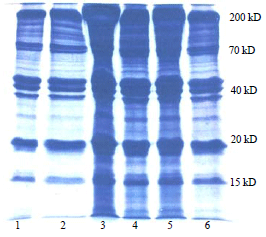
Electrophoresis of muscle and hepatopancreatic proteins: The denatured gel electrophoresis of total soluble proteins of hepatopancreas collected from M. malcolmsonii fed on various diet formulations showed several polypeptide fractions. In untreated sample (control) the polypeptides were electrophoresed in the molecular weights ranging from 200-20 kD. Based on the order of migration, various polypeptides (in the case of prawns fed with bacterial diet formulations till the 15th day), which were intensely stained, were classified as 200 kD polypeptide, 70 kD polypeptide (there were two sub-units in this fractions), 40 kD polypeptide and a 20 kD polypeptide (Fig. 2, lane-2).
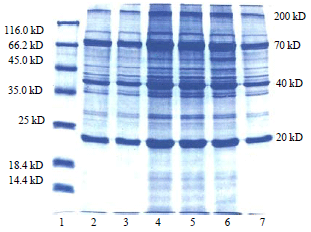 | |
| Fig. 2: | Electrophorogram of proteins in the hepatopancreas of M. malcolmsonii fed on bacteria incorporated diets for 15 days |
Lane 1: Marker, 2: Control diet fed prawns, 3: Control agar coated diet fed prawns, 4: Bacillus subtilis coated diet fed prawns, 5: Bacillus subtilis fermented diet fed prawns, 6: Pseudomonas fluorescens coated diet fed prawns and 7: Pseudomonas fluorescens fermented diet fed prawns | |
 | |
| Fig. 3: | Electrophorogram of proteins in the hepatopancreas of M. malcolmsonii fed on bacteria incorporated diets for 30 days |
Lane 1: Marker, 2: Control diet fed prawns, 3: Control agar coated diet fed prawns, 4: Bacillus subtilis coated diet fed prawns, 5: Bacillus subtilis fermented diet fed prawns, 6: Pseudomonas fluorescens coated diet fed prawns and 7: Pseudomonas fluorescens fermented diet fed prawns | |
In addition to these major polypeptides, there were several minor fractions also in all the profiles. Among various diet formulation treatments, maximum accumulation of all the polypeptide fractions was seen in the hepatopancreas samples of prawn, P. malcolmsonii fed with "Bacillus subtilis coated" feed. Less accumulation of polypeptide was noted in the case of samples from prawns fed with "Pseudomonas fluorecens fermented" and "control agar coated" diet formulations (Fig. 2. lanes-3 and 7).
 | |
| Fig. 4: | Electrophorogram of proteins in the muscle of M. malcolmsonii fed on bacteria incorporated diets for 15 days |
Lane 1: Marker, 2: Control diet fed prawns, 3: Control agar coated diet fed prawns, 4: Bacillus subtilis coated diet fed prawns, 5: Bacillus subtilis fermented diet fed prawns, 6: Pseudomonas fluorescens coated diet fed prawns and 7: Pseudomonas fluorescens fermented diet fed prawns | |
Almost a related pattern of polypeptide accumulation was seen with both "Bacillus subtilis-fermented" and "Pseudomonas fluorescens-coated" diets (Fig. 2. lanes-5, 6). A comparable pattern but with increased quantities of all the polypeptides was observed in the case of prawns fed till the 30th day with various diet formulations (Fig. 3). Densitometer scanning carried out for these samples also reported similar results.
For the prawn samples taken on the 15th day of feeding experiment, the electrophorogram of muscle protein profile showed few major polypeptide fractions and several minor polypeptide fractions (Fig. 4). Based on the staining intensities of various polypeptides, there were major fractions at 70 kD region (there were two sub-units in this fraction), 20 kD region and 15 kD regions in all the samples in the gel. As in the case of hepatopancreas, in muscle samples also, more accumulation of various polypeptides was observed in "Bacillus subtilis-coated" diet treated prawns followed by "Bacillus subtilis fermented", "Pseudomonas fluorescens-coated "Pseudomonas fluorescens-fermented" and "control coated" diet formulations (Fig. 4). A similar profile of electrophorogram of various polypeptides was observed for 30 days fed samples of prawn, with a pattern of increased accumulation of all polypeptides (Fig. 5). Densitometer scanning also revealed similar results.
Lipids
Total lipids: Lipid varied from 5.55-9.33 mg g–1, on a dry weight basis (Table 4). In general, lipids showed greater increase in rate till 30 days of feeding and there after the levels were more or less stabilized.
| Table 5: | Fatty acid composition of M. malcomsonii abdominal muscle fed on varied diets at 45th day |
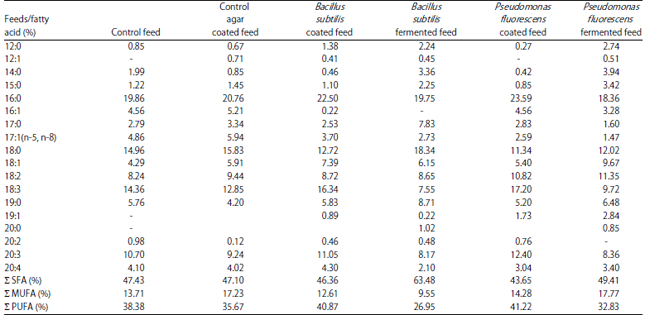 | |
Σ SFA: Saturated fatty acids, Σ MUFA: Mono unsaturated fatty acids, Σ PUFA: Poly unsaturated fatty acids | |
Analysis of variance and the related post hoc tests revealed that the lipid increase was maximum in animals fed with P. fluorescens coated feed and then to a lesser extent in prawns fed with B. subtilis coated feed and the feed fermented with the same strain. Lipid increase was least in ‘control’, control coated and P. fluorescens fermented feeds. SNK test confirmed that with the progression experiment days, the lipid content also increased-the maximum level by the 30th day.
When modified to suit the ‘unit animal’ (1 g weight initial), the lipid levels showed their magnitude of increase with respect to feeding. At the end of 60 days animals fed with B. subtilis coated diet registered a five-fold increase in lipid level, over the initial. P. fluorescens coated feed also elicited a similar magnitude of increase. Among the fermented feeds, B. subtilis fermented ones had an edge over the P. fluorescens fermented. Animals fed with ‘control“ and ‘control coated diets registered only a two-fold increase in lipid level.
Fatty acid profile: Fatty acid composition of the muscles of M. malcolmsonii fed for 45 days on diets bound with bacteria are presented in Table 5. Prawns, which were fed on control feed had 47.43% of unsaturated fatty acids with palmitic (16:0) and stearic (18:8) acids as the main components. Mono-unsaturated fatty acids were less in composition (13.71%).
 | |
| Fig. 5: | Electrophorogram of proteins in the muscle of M. malcolmsonii fed on bacteria incorporated diets for 30 days |
Lane 1: Marker, 2: Control diet fed prawns, 3: Control agar coated diet fed prawns, 4: Bacillus subtilis coated diet fed prawns, 5: Bacillus subtilis fermented diet fed prawns, 6: Pseudomonas fluorescens coated diet fed prawns and 7: Pseudomonas fluorescens fermented diet fed prawns | |
Polyunsaturated fatty acid concentration was considerable 38.38%). Similar fatty acid profile could also be observed in the prawns fed on normal feed coated with agar. Percentage fatty acid composition in bacteria-incorporated diet fed prawns did not show any prominent variation and retained the composition more or less the same as for ‘control-fed’ groups. Saturated fatty acid percentage was more in animals fed on B. subtilis-fermented feed (63.48%), while polyunsaturated fatty acids were more in prawns fed with P. fluorescens coated feed (41.22%). Oleic (18:1), linoleic (18:2) and linolenic (18:3) acids constituted the unsaturated components, while highly unsaturated fatty acids were represented in meagre quantity by arachidonic acid (20:4).
Carbohydrates: Carbohydrate content of the prawns during the entire feeding period varied from 4.34-19.35 mg g–1 of muscle (Table 4). Increase in carbohydrate content in relation of feeding was most prominent in prawns fed with B. subtilis coated diet, followed closely by those fed on B. subtilis-fermented diet. This observation was further affirmed by the non-homogeneous subsets of harmonic mean values for these two diet groups. Carbohydrate increase of the "control", ‘control coated’ and P. fluorescens fermented diet-fed prawns was at a lesser extent, while the P. fluorescens coated diet-fed group registered a moderate increase.
Beyond the initial stage of the experiment, the increase in carbohydrate was substantial and significant for all the 15 days interval groups. Values when assigned to a unit animal undergoing growth. At the end of 60th day period, animals fed on P. fluorescens-fermented diets showed a steady increase of approximately three times, from the initial level. Prawns subsisted on B. subtilis coated feed, also could register an increase of 2.5 fold. However, the animals fed on diets such as ‘control“ coated B. subtilis fermented and P. fluorescens coated, could achieve a two fold increase or even less, at end of the 60th day.
DISCUSSION
Probiotics should have a positive influence on the health of the cultured animal. Their beneficial effects may be manifested through the act of modifying host-associated or ambient microbial community, by ensuring improved use of feed or enhancing its nutritional value and improving the host animal’s immunity against diseases. Besides their role as ‘biocontrol agents’ in aquaculture, probiotics may also perform in a nutritional way18. Hence, in the present evaluation study on probiotic candidates, their nutritional role also has been taken into consideration, in addition to their health-promoting features.
Although, B. subtilis-RCMB21 and P. fluorescens-RCMB39 were harmless as seen in the previous chapter, large scale exposure to a single species of bacteria might give rise to abnormal changes rarely in the target organisms-in their external features or behaviour. Hence, the prawns were exposed to B. subtilis-RCMB21 and P. fluorescens-RCMB39, separately, for 15 days. In both the cases, P. malcolmsonii behaved normally-regular in feeding and without any apparent change externally, thus confirming the non-pathogenic nature of the candidate probionts.
The scope of the present investigation being the use of bacteria as a nutritional probiotic, it was necessary to find a vehicle for their entry into the animals. Earlier investigators adopted several methods such as using exclusive live bacterial feed or their spores, where the intention was chiefly to utilize its nutritive value4. Several other methods have also been suggested, such as (i) Addition to the formulated feed, (ii) Addition to the culture water19, (iii) Bathing the animals in bacterial suspension20 and (iv) Addition through enriched or encapsulated live food21. Use of formulated feeds is the regular feeding method in prawn and marine shrimp farms and also a convenient method for regular monitoring. Hence, in the present study, for the probiont delivery into the gut of the prawn, the bacteria were incorporated in a commercially available diet, taking care to destroy the bacteria already existing in them. Gildberg et al.22 also followed such a method of fortifying commercial feed with probiotic candidates to feed the Atlantic Gadus morhua. Most of the earlier researches advise against the use of 100% bacterial biomass as food as the recipient species may be deprived of some essential nutrients like specific amino acids or fatty acids, which may be absent in the bacteria given. Also the high level of nucleic acids in bacteria may cause accumulation of uric acid in vertebrates23. Several acceptable dietary levels of bacterial SCP have been suggested earlier and these dietary levels range from 0-50%3,7. In studies involving Bacillus and Pseudomonas strains as replacement for regular micro algal diet for Penaeus monodon larvae, less than 10% substitution could give appreciable level of survival (> 60%) and good rate of growth3. Considering the enormous quantity bacterial mass to be generated on every fourth day, that too for two sets of experiments, the incorporation level of bacteria was restricted to 3% at present. Two methods of bacterial incorporation, followed in the present investigation were coating the bacteria over pellets and fermenting the diet with bacteria.
Coating was done with help of agar as a binder in the present study, several other binders including fish oil24, have been used for bacterial incorporation in the feed. However coating with agar was quick and convenient under laboratory circumstances and it in no way deteriorated the water quality. Even with 3% incorporation of bacterial cells, large number of bacteria could be bound to the feed (5.67×1023 cells g–1 feed in the case of B. subtilis and 1.4×1026 cells g–1 feed in the case of P. fluorescens).
For preparing fermented feed also same quantity of bacterial cells were administered to the feed and they were sufficient enough to bring about quick fermentation. The fermented product could be readily distinguished by their vinegarish smell and darkened appearance. The fermentation method followed in present study was the one followed by Ramesh and Lonsane11 for the enhancement of enzyme production (alpha-amylase) by Bacillus megaterium. Fermentation promotes bioconversion of the substrate’s chemical components, improves its flavour or aroma and improves the shelf life of the product. Health benefits are also attributed to fermented products. Fermentation of fishery products using Bacillus strains has been long in practice as a preservation measure23.
Though not to a large extent, a noticeable level of variation in the biochemical constituents was evident in the bacteria incorporated diets. There were higher amounts of protein in the feeds coated as well as fermented with B. subtilis and P. fluorescens. Lipid level was lesser in B. subtilis coated feed and in the feed fermented with the same strain. However in P. fluorescens coated feed the lipid was greater than the control (14.8%) (Table 4). Carbohydrate levels were particularly less in fermented feeds. Apparently bacteria mediated conversion or utilization might have contributed to these changes in constitution. Involvement of Pseudomonas and Bacillus strains in the conversion of hexose sugars has been documented25. Bacterial cellular proteins also might have contributed to the increase level of protein in the bacteria incorporated diets. There were no remarkable difference in the nutritive value of ‘control’ diets and ‘agar coated’ diets.
During feeding experiments, it could be observed that animals were quite active and showed no rejection of the bacteria-incorporated feeds. Animals moulted regularly, almost close to the established interval of 21 days26, showing regularity in their growth. Mortality was infrequent and could not be related to any specific experimental situation.
Looking into the results and the inferences coming through the statistical test (two-way ANOVA and Student-Newman Keul’s Test) it was evident that bacteria incorporated diet had significant influence in promoting the growth and nutritional quality of M. malcolmsonii and the effect with each feed was unique.
Live bacteria coated diet had a remarkable influence on the protein content: B. subtilis could bring forth a fivefold increase in the whole animal and P. fluorescens, a four and half times increase. Though not to be same extent the fermented diets also could induce a remarkable increase in protein content. These results underscore the concept of the use of bacteria as single cell protein, in addition to their other health promoting effects. According to Tacon et al.2 single cell proteins offer many advantages over conventional animal and plant proteins in that they are highly proteinaceous (40-80% on a dry weight basis), have short generation time, can be cultivated in limited space over available carbon sources and their nutritional composition can be controlled or altered by genetic manipulation.
Polyacrylamide gel electrophoresis study also revealed the increase of polypeptide fractions in the muscles and hepatopancreas of M. malcolmsonii, in relation to the uptake of both bacteria-coated and fermented feeds. Higher accumulation of polypeptides could obviously be attributed to the increased’ nutritional factors available to the prawns, through the bacteria-incorporated/-modified diets. Bacteria like Pseudomonas are known for their production of essential amino acids like methionine and arginine that are required for protein synthesis and for promoting growth4,23. Hepatopancreas, being an organ of storage and mobilization of proteins26, also reflected the effect of nutritional factors brought about by bacteria. The polypeptide profile of hepatopancreas observed for M. malcolmsonii in the present study was comparable with the observations of Sivakumar 27 in the same species.
As in the case of proteins, a diet related fat content increase was observed presently in M. malcolmsonii (Table-4). Compared to the control diets, P. fluorescens coated diet and B. subtilis incorporated (coated and fermented) diets showed higher level of lipids, which may be directly related to the lipid content of the bacteria. P. fluorescens was observed to have the highest amount of lipids, particularly unsaturated fatty acids. While observing the fatty acid composition of M. malcolmsonii (Table 5), the quantitative and qualitative diversity was more in prawns fed on the bacteria-included diets with obvious dominance of poly-unsaturated fatty acids in P. fluorescens-coated diets (41.22%). Generally, in animals, the most common saturated fatty acids are palmitic acid (6:0) and stearic acid (18:0)28. Along with these fatty acids, M. malcolmsonii also had linolenic acid (18:3) as the dominant polyunsaturated fatty acid-linolenic acid and linoleic acid (18:2) are not synthesized by animals Tacon et al.2 and the source of these can only be dietary, particularly plants. In M. malcolmsonii also the source of PUFA may be diet supplied, which is mainly of plant and animal origin. B. subtilis and P. fluorescens, the bacterial components of the diets, were also not rich in these PUFA. However, it is noteworthy that, many of the highly unsaturated fatty acids can be synthesized by fish and prawns through chain elongation and desaturation reactions2. Generally PUFA are important in maintaining the stability and permeability of biomembranes, particularly in the vascular and nervous systems29. Vergroesen, arachidonic acid has been observed as a HUFA in M. malcolmsonii in the present study. Arachidonic acid is the precursor for the biosynthesis of prostaglandins30, which have a definite role in structural integrity of phospholipids and membrane permeability31. The major saturated fatty acid component, palmitic acid (16:0) has been suggested as an energy source in M. malcolmsonii, particularly during embryogenesis32.
Carbohydrates, the primary energy source in animals was also found to be improved with the diets with bacteria as a component. The effect was more pronounced with B. subtilis incorporated (coated and fermented) diets. Often microbes catabolise external sources of carbohydrates. However, they also are known to have cellular starch and glycogen reserves23 and apparently these might have contributed to the high carbohydrate content observed in prawns fed on bacteria combined diets.
The attainment of better nutritional quality and well-being in M. malcolmsonii could also manifested in the increase of externally perceivable parameters like length and weight. Being a good nutrient source, the bacteria might have presented themselves as food to the prawn, also because of their rich exo-enzyme production traits, they might have converted the diet into easily absorbable form-through fermentation process externally or contribution to digestion internally. Being a source of macro-and micro-nutrients and also the contribution of enzymes for digestion are accepted criteria for probionts18. Hence, the two strains of bacteria, promoted here as probiont candidates, B. subtilis-RCMB21 and P.fluorescens-RCMB39, have met the required standard to an appreciable extent.
CONCLUSION
The beneficial effects of the incorporated bacteria may be attributed to their enzymes which aid in the digestion of the given diet and also the utilization of bacteria as food by the prawn. Beneficial bacteria may thus also be utilized as single cell proteins.
SIGNIFICANCE STATEMENT
The present investigation reveals the use of bacteria as nutritional probiotic and to find a vehicle for their entry into the animals. Thus this study help the researchers to uncover the critical area of using bacterial biomass as food for freshwater prawn Macrobrachium malcolmsonii, which being the source of macro-and micro-nutrients and also the contribution of enzymes for digestion. Thus, a new theory on using these two bacterial strains, Bacillus subtilis-RCMB21 and Pseudomonas fluorescens-RCMB39 as single cell protein (SCP), may be arrived at.
ACKNOWLEDGMENT
The grants from University Grant Commission (UGC) (2012-2015), MoES-OASTC (2011-2015), New Delhi, Government of India, were of great aid for the completion of the research and the authors acknowledge these agencies for their support.
REFERENCES
- Tinh, N.T.N., K. Dierckens, P. Sorgeloos and P. Bossier, 2008. A review of the functionality of probiotics in the larviculture food chain. Mar. Biotechnol., 10: 1-12.
PubMedDirect Link - Mohamed, K.S., 1996. Heterotrophic marine bacteria as supplementary feed for larval Penaeus monodon. Naga ICLARM Q., 19: 23-26.
Direct Link - Sahu, M.K., N.S. Swarnakumar, K. Sivakumar, T. Thangaradjou and L. Kannan, 2008. Probiotics in aquaculture: Importance and future perspectives. Indian J. Microbiol., 48: 299-308.
CrossRefDirect Link - Tseng, D.Y., P.L. Ho, S.Y. Huang, S.C. Cheng, Y.L. Shiu, C.S. Chiu and C.H. Liu, 2009. Enhancement of immunity and disease resistance in the white shrimp, Litopenaeus vannamei, by the probiotic, Bacillus subtilis E20. Fish Shellfish Immunol., 26: 339-344.
CrossRefPubMedDirect Link - Gatesoupe, F.J., 2008. Updating the importance of lactic acid bacteria in fish farming: Natural occurrence and probiotic treatments. J. Mol. Microbiol. Biotechnol., 14: 107-114.
PubMedDirect Link - Kiessling, A. and S. Askbrandt, 1993. Nutritive value of two bacterial strains of single-cell protein for rainbow trout (Oncorhynchus mykiss). Aquaculture, 109: 119-130.
Direct Link - Intriago, P. and D.A. Jones, 1993. Bacteria as food for Artemia. Aquaculture, 113: 115-127.
Direct Link - Gomez, G.D. and J.L. Balcazar, 2008. A review on the interactions between gut microbiota and innate immunity of fish. FEMS Immunol. Med. Microbiol., 52: 145-154.
CrossRefDirect Link - Kolanchinathan, P., P.R. Kumari, T.S. Gnanam, G. John and A. Balasundaram, 2017. Performance evaluation of two probiotic species, on the growth, body composition and immune expression in Penaeus monodon. J. Fish. Aquatic Sci., 12: 157-167.
CrossRefDirect Link - Ramesh, M.V. and B.K. Lonsane, 1987. Solid state fermentation for production of amylase by Bacillus megaterium 16M. Biotechnol. Lett., 9: 323-328.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Roe, J.H., 1955. The determination of sugar in blood and spinal fluid with anthrone reagent. J. Biol. Chem., 212: 335-343.
Direct Link - Folch, J., M. Lees and G.H.S. Stanley, 1957. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem., 226: 497-509.
CrossRefPubMedDirect Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Nimrat, S., P. Tanutpongpalin, K. Sritunyalucksana, T. Boonthai and V. Vuthiphandchai, 2013. Enhancement of growth performance, digestive enzyme activities and disease resistance in black tiger shrimp (Penaeus monodon) postlarvae by potential probiotics. Aquacult. Int., 21: 655-666.
Direct Link - Silva, E.F., M.A. Soares, N.F. Calazans, J.L. Vogeley, B.C. do Valle, R. Soares and S. Peixoto, 2012. Effect of probiotic (Bacillus spp.) addition during larvae and postlarvae culture of the white shrimp Litopenaeus vannamei. Aquacult. Res., 44: 13-21.
CrossRefDirect Link - Bruno, M.E.C. and T.J. Montville, 1993. Common mechanistic action of bacteriocins from lactic acid bacteria. Applied Environ. Microbiol., 59: 3003-3010.
PubMedDirect Link - Gildberg, A., H. Mikkelsen, E. Sandaker and E. Ringo, 1997. Probiotic effect of lactic acid bacteria in the feed on growth and survival of fry of Atlantic cod (Gadus morhua). Hydrobiologia, 352: 279-285.
CrossRefDirect Link - Robertson, P.A.W., C.O. Dowd, C. Burrells, P. Williams and B. Austin, 2000. Use of Carnobacterium sp. as a probiotic for Atlantic salmon (Salmo salar L.) and rainbow trout (Oncorhynchus mykiss, Walbaum). Aquaculture, 185: 235-243.
CrossRefDirect Link - Chand, R.K. and P.K. Sahoo, 2006. Effect of nitrite on the immune response of freshwater prawn Macrobrachium malcolmsonii and its susceptibility to Aeromonas hydrophila. Aquaculture, 258: 150-156.
Direct Link - Rautava, S., M. Kalliomaki and E. Isolauri, 2002. Probiotics during pregnancy and breast-feeding might confer immunomodulatory protection against atopic disease in the infant. J. Allergy Clin. Immunol., 109: 119-121.
PubMedDirect Link - Araujo, M.A. and A.L. Lawrence, 1993. Fatty acid profiles of muscle and midgut gland from wild Penaeus californiensis, P. occidentalis, P. setiferus, P. stylirostris and P. vannamei. J. World Aquacult. Soc., 24: 425-428.
CrossRefDirect Link - Mourente, G., G.J. Bell and D.R. Tocher, 2007. Does dietary tocopherol level affect fatty acid metabolism in fish. Fish Physiol. Biochem., 33: 269-280.
Direct Link