
Rony Marsyal Kunda
Department of Biochemistry, Faculty of Veterinary Medicine, Universitas Gadjah Mada, Yogyakarta, Indonesia
LiveDNA: 62.16743
Niken Satuti Nur Handayani
Department of Genetic, Faculty of Biology, Universitas Gadjah Mada, Yogyakarta, Indonesia
LiveDNA: 62.16385
Hery Wijayanto
Department of Anatomy, Faculty of Veterinary Medicine, Universitas Gadjah Mada, Yogyakarta, Indonesia
Rini Widayanti
Department of Biochemistry, Faculty of Veterinary Medicine, Universitas Gadjah Mada, Yogyakarta, Indonesia
LiveDNA: 62.16531
Asian Journal of Animal and Veterinary Advances
Year: 2017 | Volume: 12 | Issue: 5 | Page No.: 227-238







ABSTRACT
Background and Objective: The DNA barcode can be used as a basis for determining the objectives and direction of development of conservation activities and can clarify differences and kinship relationships between species in the population. This study aimed to determine nucleotides sequences as DNA barcode for investigation of genetic diversity and phylogenetic from cuscuses in Maluku and Papua as well as other biogeography on COX3 gene with sequencing method. Materials and Methods: The genome DNA from 25 samples, extracted according to the Qiamp DNA protocol is the Mini Blood Kit (Qiagen) and is used as a template for COX3 gene amplification using PCR method. Results: Nucleotides alignment of Phalanger members found 23 nucleotides distinguishing and Spilocuscus members found 8 nucleotides distinguishing, which can be used as barcode. The phylogram tree was using Neighbor joining classified cuscus from Maluku and Papua in clade A (Phalanger) and clade B (Spilocuscus), respectively. Clade A and clade B subdivided into clade A1 (Phalanger from Maluku), A2 (Phalanger from Papua) and clade B1 (Spilocuscus from Maluku), B2 (Spilocuscus from Papua). Clade B2 subdivided into one subclade B2' be under cuscus from Halmahera dan Sentani, Jayapura Islands. Conclusion: It was concluded that identification of Spilocuscus and Phalanger members from Maluku, Papua, Halmahera and Sentani could be distinguished by COX3 gene. Spilocuscus maculatus from Halmahera is derived from Papua, it introduced in the colonization of the Austronesian tribes on the Island. S. maculatus from Halmahera and Sentani Islands have one nucleotide distinguish, that is sites position 354 which distinguishes the cuscuses with cuscus from Papua and Maluku.
PDF Abstract XML References Citation
Received: June 12, 2017;
Accepted: August 01, 2017;
Published: August 15, 2017
How to cite this article
Rony Marsyal Kunda, Niken Satuti Nur Handayani, Hery Wijayanto and Rini Widayanti, 2017. DNA Barcoding of Cuscuses (Marsupialia: Phalangeridae) from Maluku and Papua. Asian Journal of Animal and Veterinary Advances, 12: 227-238.
DOI: 10.3923/ajava.2017.227.238
URL: https://scialert.net/abstract/?doi=ajava.2017.227.238
DOI: 10.3923/ajava.2017.227.238
URL: https://scialert.net/abstract/?doi=ajava.2017.227.238
INTRODUCTION
Mitochondrial DNA (mt-DNA) is the genetic material with a simple molecular structure, exists outside the nucleus in eukaryotic cells and have become a widely used tool for addressing phylogenetic relationships at various levels taxons1-3. The importance of using comprehensive amount of mitochondrial sequence data for inferring phylogenetic relationships was demonstrated by Kunda et al.2 and Widayanti et al.3. In fact, the rapid rate of change in the mt-DNA sequence results in differences between populations that have only been separated for long periods of time. In other words, sequence divergences are much larger among species than within species and thus mt-DNA genealogies generally capture the biological discontinuities recognized by taxonomists as species. Taking advantage of this fact, taxonomic revisions at the species level now regularly include analysis of mt-DNA divergences1. Species revision of cuscuses have been defined, in part, on the basis of divergences in their mt-DNA1-3.
Cuscuses are arboreal marsupials are divided into the 6 genera, Ailurops, Phalanger, Spilocuscus, Strigocusucus, Trichosurus and Wyulda. Based on morphological characters, there are 4 species of cuscuses in Maluku: Spilocuscus maculatus (spotted/white cuscus), Spilocuscus rufoniger (black spotted cuscus), Phalanger orientalis (northern common cuscus) and Phalanger vestitus (stein cuscus)2,3. The local people sometimes seem to recognise more than two species but this is probably due to sexual and developmental dimorphism in coat colour4. Both species taken for food by the local people5.
In Papua, there are more than six species of cuscuses based on morphological characters: Phalanger gymnotis (ground cuscus), P. orientalis (northern common cuscus), Spilocuscus maculatus (spotted/white cuscus), Spilocuscus rufoniger (black spotted cuscus), Spilocuscus papuensis (Waigeo cuscus) is endemic species of Waigeo Island, West Papua Province3 and Spilocuscus wilsoni (Black spotted cuscus) is endemic species to the islands of Biak and Supiori in the Cenderawasih Bay, Papua Province6.
The spotted cuscuses are a group of colorful, medium sized, arboreal frugivore-folivores endemic to tropical forests in the Australo-Papuan region. They are found in primary and secondary tropical moist forest. They also have been recorded in mangrove forest7. The female thought to carry one single young only5. Among Phalangeridae genera, Spilocuscus characterized by a unique combination of traits, including sexual dichromatism in pelage coloration and pronounced sexual dimorphism, with females larger than males. It is powerfully built and covered in thick woolly fur and weighs about 1-3 kg5. The males are usually spotted, but sometimes also completely white, whereas the larger females are usually non-spotted and grey-brown in colour. The young goes through a sequence of colour changes. Its solitary, nocturnal life style means that it hides in the 3 crowns or holes, or among masses of epiphytic vegetation during the day2,5.
Recent molecular studies have included of the Spilocuscus and Phalanger genera members from Maluku and Papua1,3, using 12S rRNA and NADH Dehydrogenase Sub-unit 1 (ND1) genes and using of Cyt b gene sequences for status affirmation of stein cuscus from Manipa Island2. It was asserted that cuscus from Manipa Island has genetic relationship with Spilocuscus maculatus (94%) than Phalanger members2.
If a short region of mt-DNA that consistently differentiated species can be found and accepted as a standard, a library of sequences linked to vouchered specimens would make this sequence an identifier for species, a "DNA barcode"8,9. Recent work suggests that a 648 bp region of the mitochondrial gene, cytochrome oxidase I (COXI), might serve as a DNA barcode for the identification of animal species9,10. The DNA barcoding usually consists of a fragment of the mitochondrial gene Cytochrome oxidase c subunit I (COX1) but other genes are also employed, sometimes with varying levels of success9. The method has many applications among which is an efficient means of identifying species because levels of divergence among individuals are usually much lower of the same species than between closely related species.
Past phylogenetic work often focused on mitochondrial genes encoding ribosomal (12S, 16S) DNA, but their use in broad taxonomic analyses is constrained by the prevalence of insertions and deletions (indels) that greatly complicate sequence alignments1. Cytochrome c oxidase subunit III gene (COX3) has an important advantage, COX3 gene appears because possess a greater range of phylogenetic signal besides COX1 gene. Sequences of COX3 gene is more divergent than COX1 and Cyt b gene sequences11. However, there is limited information on the suitability of the COX3 gene for the phylogenetic analysis within mammalian species. In fact, the evolution of these genes are slow enough to allow discrimination not only allied species, but also phylogeographic groups within a species12. This research will provide phylogeny and useful information providing a clear understanding of population history, evolutionary mechanisms and can be used as a genetic marker (barcodes) of the same genus or species.
The purposes of this study as preliminary study on mitochondrial genome based on COX3 sequences that declared as a DNA barcode for investigation of genetic diversity and phylogenetic from cuscuses in Maluku and Papua as well as other biogeography. Such tests require the analysis of phalangeridae members (Spilocuscus and Phalanger) from Maluku and Papua that have been before studied intensively so that declared COX3 gene as a DNA barcode. Cuscus conservation strategies will be more organized and useful if genetic marker can be known with certainty. It is hoped that the COX 3 gene sequences may act as a specific genetic marker (barcode) between species of cuscuses in the populations. Data from this study can be used to assist in the determination of conservation units as well as the determination of conservation strategies against these endemic protected animal.
MATERIALS AND METHODS
Samples collection: Twenty-five tissue biopsy samples were collected from two cuscuses of native habitat in Maluku (16 samples), Papua (9 samples) and, Halmahera Island (1 individual). All individuals ware identified on the basis of morphological characteristics and a sample tissues collected from each species and preserved in RNA Latter buffer (Qiagen) for the purpose of DNA isolation.
Molecular techniques and data analysis: Genomic DNA isolated from the tissues using DNA isolation kit (Qiagen) according to protocol method in combination with RNase treatment. The primer used to amplify COX3 gene was as follows: The PCR protocol ware: The DNA template 1 μL, Kappa 2G Ready Mix 25 μL, each forward and reverse primer 1 μL (10 pmol) and added H2O in a total of 50 μL. The PCR amplification was conducted using a Infigen Thermal Cycler according to the program, an initial denaturasi at 94°C for 5 min, denaturation at 94°C for 30 sec, annealing at 50°C for 30 sec and extension at 72°C for 1 min and 30 sec. A final extension at 72°C for 5 min was performed to completely extend the amplified product. The DNA quality assessed by electrophoresis on 1.5% agarose gel (3 μL PCR products addend with 1 μL of loading dye). Electrophoresis was run on 100 Volt condition for 30 sec and amplification result could be seen on the UV light. The purified PCR products were sequenced directly by 1st BASE Sequencing INT (Singapore).
The DNA sequences of COX3 gene were initially automatically alignments using Clustal W, followed by manual editing. Phylogenetic analysis of the aligned sequences was performed with the Molecular Evolutionary Genetics Analysis (MEGA) program version. 6.0. Distance analysis was conducted using the Neighbor-Joining (NJ) option employing the Kimura-2-parameter method with gap data treated as pair wise deletions. Molecular phylogeny analysis was conducted using heuristic searches, 1000 bootstrap replicates.
RESULTS
A total of 25 DNA samples have been isolated, then serves as a template for COX3 gene amplification by PCR method. The results PCR products 919 base pair (bp) after electrophoresis on a1% agarose gel with a 1 kilo base pair (kbp) DNA Ladder (1st base) are shown in (Fig. 1).
The PCR product size is 919 bp obtained after the primers specific that will be used as alignment using the BLAST program against mitochondrial genome P. vestitus (access numbers AB241057.1). In this study, 8 species were used for comparison to determine the genus and species of cuscuses from Maluku and Papua Island. The nucleotides sequences data were determined for the present study deposited in Genebank under accession numbers, NC_008137 (P. vestitus), NC_003039 (T. vulpecula), NC_868096 (A. ursinus), NC_868161 (S. celebensis), NC_868160 (S. maculatus), KJ_868165 (Wyulda squamicaudata), NC_003322 (Vombatus ursinus), KJ_868140 (Petrogale brachyotis).
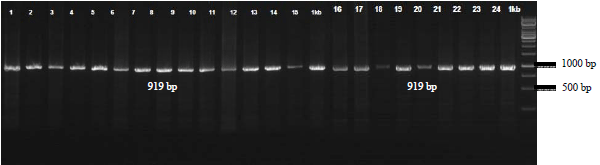 | |
| Fig. 1: | PCR Product COX3 gene of cuscuses on a 1.5% agarose gel, Description: 1-25: PCR product of cuscuses samples (919 base pair), *1 kilo base pair (kbp) = 1000 base pair (bp) |
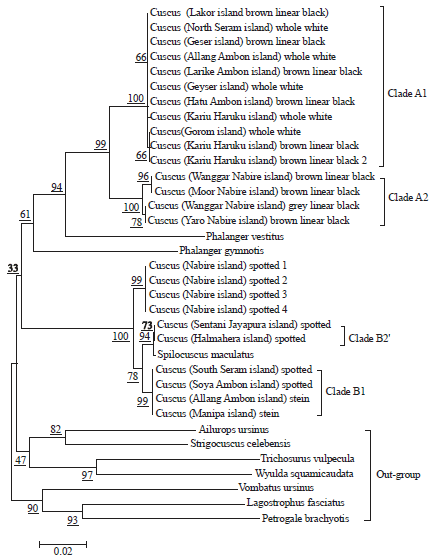 | |
| Fig. 2: | Phylogenetic relationship of cuscuses (Phalanger and Spilocuscus genera) from Maluku and Papua based on mitochondrial COX3 gene sequences |
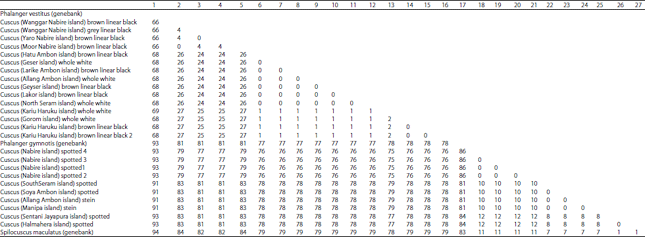
The result alignment COX3 gene was studied cuscuses obtained 783 nucleotides for subsequent analysis. There are differences in the nucleotides of 0-86 when inter cuscuses studied and 1-104 with cuscuses from Genebank (Table 1).
Analysis of the nucleotides matrix (Table 1), shows that cuscuses from Papua (Phalanger members) on the Yaro, Nabire (brown linear black), Moor Nabire (brown linear black) and Wanggar Nabire (Gray linear black) does not have different nucleotides (0 nucleotide). This also happens to cuscuses from Maluku (Phalanger members) on the Gorom island (whole white), Geser Island (whole white and brown linear black), North Seram (whole white), Lakor Island (brown linear black), Kariu village, Haruku Island (all samples) and Allang village, Ambon Island (whole white).
In this study, cuscuses from Papua (Spilocuscus members) on Nabire Island (total of four samples) does not have different nucleotides (0 nucleotide). The same thing seen in cuscuses from Maluku (Spilocuscus members) on the South Seram Island (spotted cuscus), Manipa Island (stein cuscus), Soya and Allan village, Ambon Island (spotted and stein cuscuses) does not have different nucleotides (0 nucleotide). Cuscuses collected from Halmahera Island (spotted cuscus) have 8 nucleotides (8 nucleotides) distinguish with Maluku samples, although being in one biogeographic regions, but it same as the Spilocuscus members from Papua, has no nucleotide differences (0 nucleotide) (Table 1). The NJ phylogenetic tree showed no conflicts between molecular and morphological identifications but the conflict is based on biogeography region (Fig. 2). Twenty three nucleotides distinguish can be used as a barcode among Phalanger members from Maluku and Papua and 4 nucleotides distinguish can be used as a barcode among Spilocuscus members from Maluku and Papua (Fig. 3).
| Table 1: | Number of differences (783 nucleotides) of mitochondrial COX3 gene of cuscuses with sequances comparison from Genebank database (basepair., bp) |
 | |
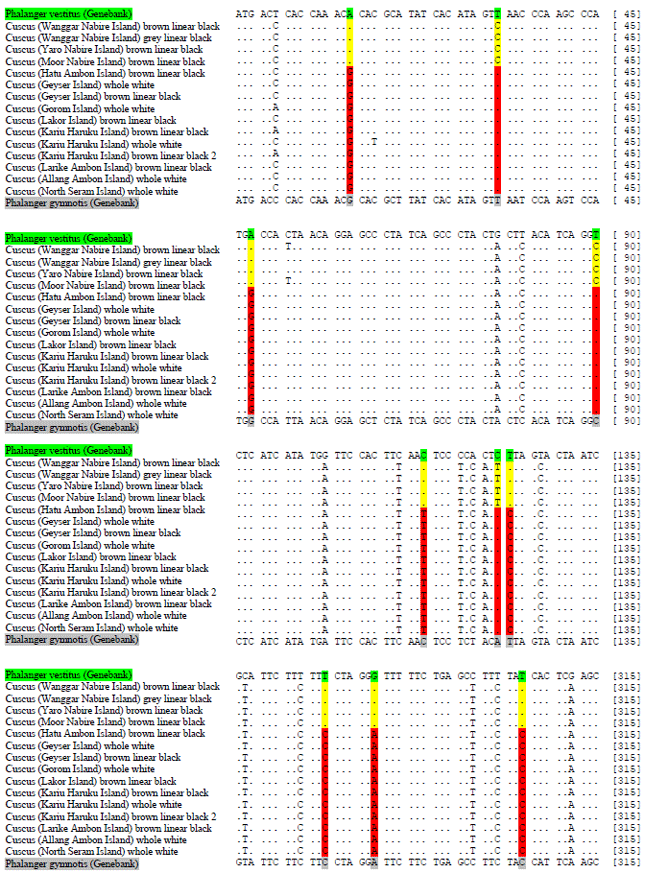 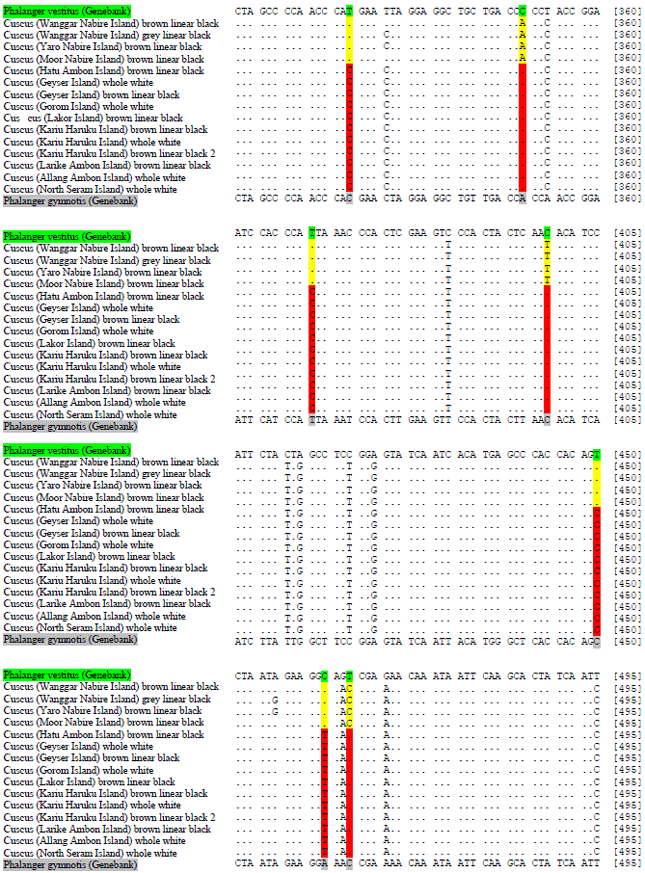 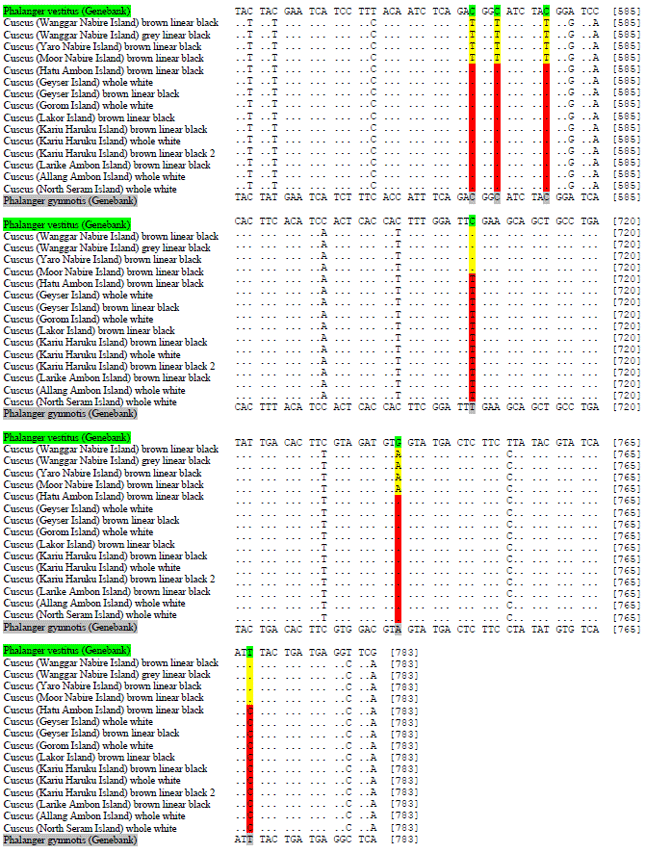 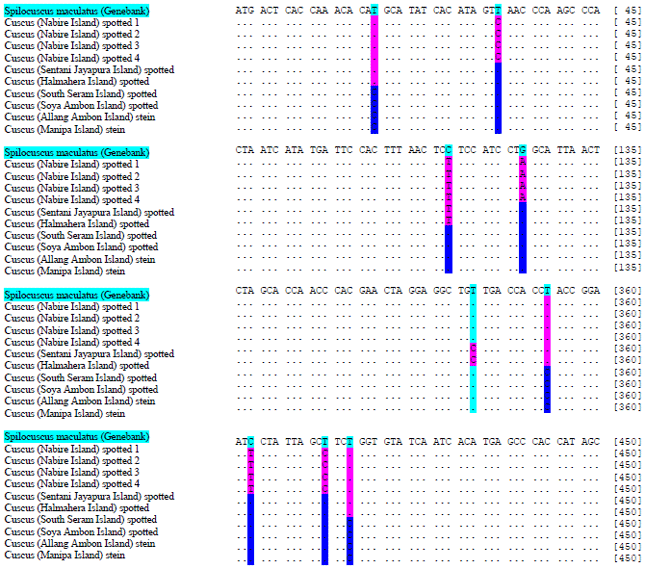 | |
| Fig. 3: | Sites nucleotides barcode of the COX3 gene of Phalanger and Spilocuscus genera from Maluku and Papua. Descriptions: *Yellow is Phalanger genera members from Papua, *Red is Phalanger genera members from Maluku, *Pink is Spilocuscus genera members from Papua and Halmahera Islands, *Dark Blue is Spilocuscus genera members from Maluku, *Green, Grey and Light Blue is Species compare from Genebank |
The occurrence of individuals groupings of Halmahera and Sentani populations Islands, in this study is possible due to the spread of cuscus from other locations causing the flow of genes between populations occurs regularly. Another possibility is that small populations around the site function as an intermediate population, so that sharp barriers such as mountains and seas that act as dispersal limiting factors do not play a role or inhibit gene flow. The Phalanger genera is composed of 2 separate clades of A1 (Maluku) supported by a bootstrap values of 100% and clade A2 (Papua) is supported by a 100% bootstrap values. Spilocuscus genera consists of 2 separate clades, B1 (Maluku) is supported by 99% bootstrap values and B2 (Papua) is supported by 100% bootstrap values. Two individuals from the Halmahera and Sentani Islands form a separate 1 sub-clade as part of the clade B2, that is clade B2' (Fig. 3).
DISCUSSION
Sites barcodes and biogeography of cuscuses from maluku and Papua: This study provides an input 23 new sequences (sites nucleotides) of Phalanger species from Maluku and Papua for the DNA barcode database and 4 new sites nucleotides of Spilocuscus species from Maluku and Papua (Fig. 3). Twenty three nucleotides distinguish can be used as a barcodes among Phalanger species from Maluku and Papua, that is sites positions 15-(G/A), 33-(T/C), 48-(G/A), 90-(T/C), 114-(T/C), 123-(C/T), 124-(C/T), 282-(C/T), 288-(A/G), 306-(C/T), 330-(C/T), 351-(C/A), 370-(C/T), 399-(C/T), 450-(C/T), 462-(T/C), 465-(T/C), 570-(C/T), 573-(C/T), 579-(C/T), 705-(T/C), 741-(G/A) and site 768-(C/T). Twenty three sites barcode is the results alignments of Phalanger members between the 2 populations (Maluku and Papua) (Fig. 3). The DNA barcode technique is a rapid way to identify and discover new species. Its success depends on the reciprocal monophyly of species and on the strength of the establishment of a "barcode gap", i.e., a clear delimitation between intra- and inter-specific sequence variability for genes sequences are studied13.
Results nucleotides alignment of Spilocuscus members from Maluku and Papua, were found 8 nucleotides distinguish between the two populations (Fig. 3), that is sites positions 18-(C/T), 33-(T/C), 117-(C/T), 126-(A/G), 354-(C/T), 408-(T/C), 417-(C/T) and site 420-(C/T). Eight nucleotides distinguish can be using as a barcodes among Spilocuscus members from Maluku and Papua. The results of alignment showed that there is substitution (transversion: Transition) in each genus of cuscuses (Phalanger and Spilocuscus). The majority of nucleotide substitutions that occur at the species level transition occurs, while at the genus level transversions occur. Substitution ratio differences at the level of species and genus can be caused by different biogeographic and metabolic rate14. In addition Kunda et al.2, it is reported that differences in nucleotide substitution ratio of a gene, can be attributed to differences in reproduction, life history, or the process of evolution and adaptation. The level of substitution of a population is interesting to study because it may vary in different organisms or genes in various organisms. Differences in nucleotide substitutions high very influential in percentage composition of nucleotides, which is in turn has implications for the phylogenetic analysis15. In addition Vali et al.15, is asserted that the scale of the difference deviation is so high nucleotide composition that will be a solid foundation and is instrumental in phylogenetic analyzes.
Changes in the composition of nucleotide substitution caused by both transversion and transition can be used as a molecular marker (barcode) to study natural populations1-3. Nucleotide substitution either transversion or transition can provide a clear understanding of the geographical evolutionary history of the population and can be used as a genetic marker (barcode)2,3. Not all of the genes can be used in the study of genetic markers and genes only with certain requirements which can be used as a barcode8.
The phylogram (Fig. 2) generated through NJ method using K2P distance was highly reliable as the out group used (Strigocuscus celebensis and Ailurops ursinus) segregated in separate clade as expected. The ability of COX3 gene in distinguishing 2 different genera was proved through the phylogram as 2 different clades (Clades A and B) distinguishable. Clade A as the Phalanger and clade B as the Spilocuscus. These results are in accordance with the declaration of Widayanti et al.1,3 and Fatem and Sawen4 that genus cuscus in Papua and Maluku is Phalanger and Spilocuscus. The results of studies using COX3 loci prove that S. maculatus origin Halmahera Island has a strong phylogeny with S. maculatus from Sentani Island. Mitochondrial sequences data from the ND1 gene supported the genetic association between S. maculatus species from Halmahera and Sentani Islands, but the study conducted by Widayanti et al.3, did not explain the correlation of genetic relationship between S. maculatus species from the 2 Islands based on biogeography.
Based on biogeography, S. maculatus from Halmahera Island, must have a kinship with S. maculatus from Maluku, but it raises a conflict, relations with the biogeographic distribution of species. The biogeographic conflict is clearly explained that about 6,500 years ago, human colonization occurred by the Austronesian tribes on the Halmahera Island. In that period they brought food crops and pets one of them S. maculatus. Large Island in Maluku, namely is Halmahera has ideal conditions and natural way to detect the translocation of animals, where irian act as the area of origin of the animals. This study proves that the results published by Widayanti et al.3, empirically proven have been demonstrated use different loci, but still maintain the stability of the phylogenetic tree between ND1 and COX3 loci. Phylogenetic signals are evident in the clade A the Phalanger of Papua segregated from the Phalanger of Maluku with few other related species (strong support by bootstrap value 100%). Similarly for phylogenetic signals are evident in the clade B the Spilocuscus of Papua segregated from the Spilocuscus of Maluku (strong support by bootstrap value 100%).
This study asserted that S. maculatus from Sentani and Halmahera Islands have a genetic kinship (94%) close to S. maculatus from Nabire. These phylogenetic 3 yields are consistent with the research reported by Widayanti et al.3 and supporting monophyly species from the Phalangeridae group. Ternate Island is far from Papua biogeographic regions, but both Spilocuscus has a very close relationship. Evidence suggests that cuscus species were transported to New Ireland (Papua New Guinea) as early as 10,000-20,000 years ago. Some species were introduced to the Solomons as early as 6600 years ago and to Timor as early as 6500 years ago. Phalanger orientalis may have been actively introduced to Seram, Buru, Sanana and Kei Islands but the initial introductory evidence is not known with certainty. The period when the spotted cuscus (S. maculatus) reached northern Maluku, so are the ornate cuscus (Phalanger ornatus) reached Northern Maluku remains enigmatic as well, though they likely arrived in Maluku well in advance of other large terrestrial mammalian species.
Historically, species-level systematics of Spilocuscus have been highly labile2,3. Within the last decade, however, taxonomy within the genus has stabilized. Recent research asserted that cuscus origin Manipa Island that is previously based on morphological characters classified into Phalanger group members, but based on genetic analysis (Cytb gene) sequences more appropriately classified into Spilocuscus group members5.
Study reported by Widayanti et al.1, examined phalangeridae relationships using mitochondrial 12S rRNA. Their results yield support for an association of Ailurops, Phalanger and Spilocuscus. Research by Raterman et al.16, asserted strong phylogenetic support found for both the S. celebensis sister taxon to Ailurops and the S. celebensis+Ailurops sister to Phalangerini relationships. First authors to include 3 genera (Ailurops from Sulawesi and Spilocuscus and Phalanger from Maluku and Papua islands) in a molecular study, used mitochondrial 12S rRNA gene1.
Results nucleotides alignment were found 3 sites and 13 sites that can be used as genetic markers between Spilocuscus and Phalanger members from Papua and Maluku1. Three sites positions of Spilocuscus genera that can be used a barcodes are 127-(G/A), 481-(C/T), 885-(T/C) and 13 sites positions of Phalanger genera are 67-(A/G), ke-89 (G/C), ke-137 (T/C), ke-285 (G/A), ke-468 (T/C), ke-595 (T/C), ke-598 (T/C), ke-647 (T/C), ke-654 (G/A), ke-665 (T/C), ke-769 (C/T), ke-874 (C/T), ke-876 (A/G)1. Added3 that cuscus from Manipa Island have genetic relationship with Spilocuscus maculatus (94%) than Phalanger members. Research conducted by Kunda et al.2 asserted that Stein cuscus from Manipa Island is closer to Spilocuscus maculatus than to Phalanger genera members. Previous molecular studies have not include specimens from Halmahera Island1. Research by Meredith et al.17 considered this taxon a close relative of S. celebensis, but retained both species in the genus Phalanger.
Research by Kunda et al.2, asserted that the colors and patterns of the hair can not be used as a basis for determining taxonomic cuscuses. Cuscuses from clade A are very varied from the colors and patterns of hairs (brown linear black, gray linear black and whole white) but the views of the nucleotides sequences from COX3 gene showed that all of them are the same species (Phalanger sp.) and have phylogenetic relationship with P. vestitus (100%) from genebank. The same thing happened at group B. They are many variations in colors and patterns of hair (spotted and stein) but it genetically the same species is Spilocuscus maculatus. The results of this study are different from the conventional taxonomy4. The availability of sequences in public reference database enables the effectiveness of DNA-based identification of species18, overcoming morphological identification difficulties.
This way, as the first broad data set of the COX3 barcoding for cuscuses species, it can potentially become an alternative or a complementary methodology to morphological identification. Accurate molecular identification to species-level can be a potential and valuable alternative to morphological identification, including phylogenetic relationships among species in the population. So, the COX3 barcoding sequences provided here can be helpful for processing phylogenetic traces from cuscuses. The molecular phylogenies presented in this study are considered more reflective on the true evolutionary relationships of Phalangeridae than previous morphological analyses, as the molecular phylogenies show that the high levels of congruence between them versus has been seen in the morphologically based on phylogenies.
CONCLUSIONS
The sequence of mt-DNA COX3 gene can be used as a DNA barcode for cuscuses from Maluku and Papua and found 23 nucleotides distinguish of Phalanger genera and 4 nucleotides distinguish of Spilocuscus genera from 2 populations (Maluku and Papua). It was proved that Spotted cuscus from Halmahera Island has the closest genetic relationship with S. maculatus from Papua (Sentani, Jayapura island) which is a species introduced from Papua when the human colonization of Austronesian tribes. Present result revealed that COX3 gene grouping cuscuses from Papua and Maluku become 2 clade A and clade B. Clade A is Phalanger genera and clade B is Spilocuscus genera and entry into Phalanger vestitus (and Spilocuscus maculatus species).
SIGNIFICANCE STATEMENT
This study discovers the possible S. maculatus from Halmahera have the closest genetic relationship with S. maculatus from Sentani, Jayapura Island which is a specie introduced from Papua when the human colonization of Austronesian tribes. This study will help researchers to uncover Phylogenetic Species Concept (PSC) that will provide an accurate understanding of the identity of a species in taxon and is an important framework for understanding biodiversity. Thus, a new theory on Phylogenetic species concept is very relevant to design a policy on what to protect, how to protect and how to facilitate biodiversity on an continuous basis.
ACKNOWLEDGMENT
This study is part of Dissertation was funded by the Directorate General of Higher Education (DGHE) Indonesia Grant No. 888/UN1-P.III/LT/DIT-LIT/2016 , March 01, 2016.
REFERENCES
- Widayanti, R., H. Wijayanto, W.D. Wendo and R.M. Kunda, 2015. [Identification of genetic diversity 12Sr RNA genes as genetic marker for determining species cuscus]. Jurnal Veteriner, 16: 227-235, (In Indonesian).
Direct Link - Kunda, R.M., N.S.N. Handayani, H. Wijayanto and R. Widayanti, 2016. Study of genetic marker of cuscuses (Marsupialia: Phalangeridae) from Maluku and Papua based on cytochrome b gene sequences. Pak. J. Biol. Sci., 19: 122-135.
CrossRefDirect Link - Widayanti, R., Agustianti, T. Suprayogi, R.M. Kunda and S. Pakpahan, 2016. Phylogenetic relationship of cuscuses (Marsupialia: Phalangeridae) from papua and maluku based on mitochondrial sequences of NADH dehydrogenase sub-unit 1 gene. Biotechnology, 15: 17-25.
CrossRefDirect Link - Fatem, S. and D. Sawen, 2007. [The species of cuscus in Northern coastal areas of Manokwari, Papua] Biodiversitas, 8: 233-237, (In Malay).
Direct Link - Zein, M.S.A. and D.M. Prawiradilaga, 2013. DNA Barcode Fauna Indonesia. Prenadamedia Group, Jakarta, Indonesia, ISBN: 9786027985261, pp: 16-18, (In Indonesian).
Direct Link - Xia, Y., H.F. GU, R. Peng, Q. Chen, Y.C. Zheng, R.W. Murphy and X.M. Zeng, 2012. COI is better than 16S rRNA for DNA barcoding Asiatic salamanders (Amphibia: Caudata: Hynobiidae). Mol. Ecol. Resour., 12: 48-56.
CrossRefDirect Link - Madeira, T., C.M. Souza, J. Cordeiro and P.J. Thyssen, 2016. The use of DNA barcode for identifying species of Oxysarcodexia Townsend (Diptera: Sarcophagidae): A preliminary survey. Acta Tropica, 161: 73-78.
CrossRefDirect Link - Hikosaka, K., Y.I. Watanabe, N. Tsuji, K. Kita and H. Kishine et al., 2010. Divergence of the mitochondrial genome structure in the apicomplexan parasites, Babesia and Theileria. Mol. Biol. Evol., 27: 1107-1116.
CrossRefDirect Link - Fadhlaoui-Zid, K., L. Knittweis, D. Aurelle, C. Nafkha and S. Ezzeddine et al., 2012. Genetic structure of Octopus vulgaris (Cephalopoda, Octopodidae) in the Central Mediterranean Sea inferred from the mitochondrial COIII gene. Comptes Rendus Biol., 335: 625-636.
CrossRefDirect Link - Meier, R., G. Zhang and F. Ali, 2008. The use of mean instead of smallest interspecific distances exaggerates the size of the “barcoding gap” and leads to misidentification. Syst. Biol., 57: 809-813.
CrossRefDirect Link - Nabholz, B., S. Glemin and N. Galtier, 2009. The erratic mitochondrial clock: Variations of mutation rate, not population size, affect mtDNA diversity across birds and mammals. BMC Evol. Biol., Vol. 9.
CrossRefDirect Link - Vali, U., M. Brandstrom, M. Johansson and H. Ellegren, 2008. Insertion-deletion polymorphisms (indels) as genetic markers in natural populations. BMC Genet., Vol. 9.
CrossRefDirect Link - Raterman, D., R.W. Meredith, L.A. Ruedas and M.S. Springer, 2006. Phylogenetic relationships of the cuscuses and brushtail possums (Marsupialia: Phalangeridae) using the nuclear gene BRCA1. Aust. J. Zool., 54: 353-361.
CrossRefDirect Link - Meredith, R.W., M. Westerman and M.S. Springer, 2009. A phylogeny of Diprotodontia (Marsupialia) based on sequences for five nuclear genes. Mol. Phylogenet. Evol., 51: 554-571.
CrossRefDirect Link - Frezal, L. and R. Leblois, 2008. Four years of DNA barcoding: Current advances and prospects. Infect. Genet. Evol., 8: 727-736.
CrossRefDirect Link