
Gulten Ates
Department of Physiology, Faculty of Istanbul Medicine, Istanbul University, Istanbul, Turkey
Elif Ozkok
Department of Neuroscience, Institute of Experimental Medicine, Istanbul University, Istanbul, Turkey
Hatice Yorulmaz
Department of Physiology, Faculty of Medicine, Halic University, Istanbul, Turkey
Irfan Aydin
Department of Medical Laboratory, Associate`s Degree Vocational School, Beykent University, Istanbul, Turkey
Sule Tamer
Department of Physiology, Faculty of Istanbul Medicine, Istanbul University, Istanbul, Turkey
Asian Journal of Animal and Veterinary Advances
Year: 2017 | Volume: 12 | Issue: 1 | Page No.: 17-23







ABSTRACT
Background: Sepsis is a life-threatening systemic inflammatory syndrome, that affects many organs or systems. Ghrelin is released in many tissues and organs and it has anti-inflammatory and antioxidant effects. Objective: In this study, the effects of exogenous ghrelin on Tumor Necrosis Factor-α (TNF-α), interleukin-10 (IL-10), superoxide dismutase (SOD), thiobarbituric acid reactive substances (TBARS) and ghrelin levels were aimed to investigate in liver tissue after inducing sepsis with lipopolysaccharide (LPS). Materials and Methods: Wistar albino rats were divided into four groups, control, LPS, ghrelin, ghrelin+LPS. The LPS group was established by giving LPS twice (5 mg kg–1, i.v. and i.p., respectively). Ghrelin (10 nmol kg–1) was given intravenously. Tissue levels of TNF-α, IL-10, ghrelin and the activities of SOD were mesured with using an ELISA kit. The levels of TBARS were determined spectrophotometrically. Results: The levels of TNF-α and IL-10 were found higher in the LPS group than controls (p<0.01 and p<0.05, respectively) and TBARS levels were increased in the LPS group compared with the controls, ghrelin and ghrelin+LPS groups (p<0.01, p<0.05 and p<0.01, respectively). The SOD and ghrelin levels were higher in the LPS group than in the others, ghrelin levels were also increased in the ghrelin group compared with the controls (p<0.05). According to histologic imaging, there were enlargements in sinusoids in the LPS group. In the ghrelin+LPS group, the same image was seen as in the controls. Conclusion: Ghrelin treatment attenuated anti- and pro-inflammatory cytokines, TBARS levels and SOD activities and also reduced liver tissue damage in a time and dose dependent manner during sepsis.
PDF Abstract XML References Citation
How to cite this article
Gulten Ates, Elif Ozkok, Hatice Yorulmaz, Irfan Aydin and Sule Tamer, 2017. Effects of Ghrelin on Inflammation and Oxidative Stress Parameters in Sepsis-induced Liver Tissue of Rat. Asian Journal of Animal and Veterinary Advances, 12: 17-23.
DOI: 10.3923/ajava.2017.17.23
URL: https://scialert.net/abstract/?doi=ajava.2017.17.23
DOI: 10.3923/ajava.2017.17.23
URL: https://scialert.net/abstract/?doi=ajava.2017.17.23
INTRODUCTION
Sepsis causes deterioration of the integrity of the host barrier, either physical or immunological, leading to direct penetration of the pathogens into the bloodstream, which creates the septic state1.
In experimental sepsis studies, lipopolysaccharide (LPS) is used to create an experimental sepsis model. The LPS is made up of macromolecules that constitute 75% of the outer membrane of Gram-negative bacteria, which are capable of causing lethal shock. The lipid A component of LPS promotes the expression of tissue factor and pro-inflammatory cytokines, which is sufficient to induce endothelial cell injury and apoptosis. The increments in the inflammatory response in sepsis are maintained by pro-inflammatory cytokines. Although, septic shock is often associated with pro-inflammatory responses, failure of the anti-inflammatory process also plays a role in the pathogenesis of this condition2-4.
Oxidative stress is occurred disruption of balance in favor of increased free radicals to antioxidants in macromolecules including atherosclerosis, hypertension, diabetes mellitus, ischemic diseases, neurodegenerative diseases and critical illness5. It has been shown that the mitochondrial electron transport chain, xanthine oxidase and the NADPH oxidase system cause a respiratory burst, which produces reactive oxygen species including superoxide (O2–), hydroxyl (OH–), Nitric Oxide (NO) radicals during neutrophil activation and arachidonic acid metabolism in sepsis6. Membrane permeability damage occurs through modification of protein, lipid and DNA macromolecules due to free radicals7. Under normal conditions, the effects of free radicals on macromolecules are abolished or weakened by constituents of the endogenous antioxidant enzyme system, such as superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX)7.
Thiobarbituric acid reactive substances (TBARS) are a biomarker of lipid peroxidation and contribute to the development of atherosclerosis, diabetes mellitus and cardiovascular diseases. The TBARS can cause damage to macromolecules such as proteins, lipids and DNA8. A recent study showed that increased plasma TBARS levels caused damage to three or more organs in patients with sepsis9.
Ghrelin, a peptide hormone with 28 amino acids has been isolated as a specific endogenous ligand for the growth hormone secretagogue receptor (GHS-R), which stimulates growth hormone secretion and energy metabolism and has also been reported to have effects on inflammation, osteoporosis, cancer, aging, learning and memory10-12.
In recent studies, ghrelin was demonstrated to be effective in inflammation by organizing inflammatory cytokines, increasing T-cell development and its receptors on many tissue cells and macrophages12-16. Exogenous administration of ghrelin suppresses inflammatory cytokines released from monocytes during inflammation, which is stimulated by LPS and regulates neutrophil-mediated immune response T-cells proliferation and tissue and oxidative damage12,15. Ghrelin treatment has been shown to have an antioxidant effect against systemic oxidative stress in many organs such as lung, liver, heart and kidney10,16.
In this study, the effects of exogenously administered Ghrelin on the pro-inflammatory cytokine TNF-α and anti-inflammatory cytokine IL-10 and oxidative stress parameters, such as TBARS and SOD levels were aimed to research in the liver tissue of male Wistar albino rats with sepsis induced with LPS.
MATERIALS AND METHODS
Ethic: This study was conducted at the Istanbul University Experimental Research Center (Resolution No. 2013/123).
Experimental groups: Male adult Wistar albino rats (210-240 g) were divided into four groups, each comprising 8 rats: (1) Control group, (2) LPS group, (3) Ghrelin group, (4) Ghrelin+LPS group. The animals were fed with a commercial diet and tap water ad libitum, housed in cages kept at a controlled temperature (22±2°C) and humidity (55-60%) with a 12 h light/dark cycle.
Experimental procedures: Lipopolysaccharide (LPS) from Escherichia coli O127: B8 (Sigma Aldrich, Product No. L5668) was dissolved in 1 mL of sterile saline solution. The LPS group was established by giving LPS twice (5 mg kg–1, in 1 mL of 0.9% NaCl, intravenous (i.v.) and intraperitoneal (i.p.), respectively) at 12 h intervals.
Ghrelin (10 nmol kg–1, dissolved in 1 mL of sterile saline solution) was administered i.v. in the ghrelin group. In the ghrelin+LPS group, 10 nmol kg–1 ghrelin was intravenously injected simultaneously with the first LPS injection. The rats were sacrificed 24 h after the 1st injection using sodium pentothal anesthesia (i.p., 30 mg kg–1).
Hematological and biochemical procedures: At the end of the experimental period, blood samples were taken to tube with EDTA determine the leukocyte count and dry tube to determine serum concentrations of glucose, aspartate aminotransferase (AST) and alanine aminotransferase (ALT) were analyzed using a autoanalyzer (Roche Cobass Integra Systems, Roche Diagnostic/Mannheim, Germany).
Enzyme-linked immunosorbent assay (ELISA) procedure: Liver tissues were homogenized with potassium phosphate buffer and centrifuged at 2500 rpm, +4°C for 10 min. Supernatants were removed for determining the levels of total ghrelin (Cat# EZRGRT-91K-EMD millipore corparation, Billerica), TNF-α (Cat# 865.000.096-Diaclone, France), IL-10 (Cat# 670.070.096-Diaclone, France) and the activities of SOD (ESOD-100, Bioassay system) using ELISA kits.
TBARS levels in liver tissue: The levels of lipid peroxidation were assessed by measuring TBARS using a thiobarbituric acid assay according to the method of Ohkawa et al.17. After a reaction with thiobarbituric acid (TBA-Sigma, Aldrich), the reaction product was extracted in butanol and its absorbance was determined spectrophotometrically (Shimadzu, Japan) at 535 nm. The TBARS concentrations were calculated using 1,3,3,3 tetra-ethoxypropane as a standard. The results were expressed as nmol g–1 tissue.
Histologic procedures: The evaluation of liver tissue using hematoxylin and eosin (H and E) staining was done by Medical Laboratory Department, Associate’s Degree Vocational School, Beykent University. The liver tissue samples were fixed in 10% buffered formalin and embedded in paraffin. Five-micrometer thick sections were placed on polylysine-coated slides and stained with H and E. The slides were viewed and evaluated under light microscopy (Olympus BX51, Olympus Corp., Tokyo, Japan) at 10× magnification.
Statistical analyses: Data are expressed as Mean±Standard Deviation. Overall statistical significances between the groups were tested with one-way ANOVA or Kruskal-Wallis tests, depending on the normality of distribution. In all cases, p<0.05 was set as the limit of significance.
RESULTS
Cytokine results: The TNF-α levels were found significantly high in the LPS group compared with the control, ghrelin and ghrelin+LPS groups (p<0.01, p<0.05 and p<0.05, respectively) (Fig. 1a).
The IL-10 levels were significantly increased in the LPS group compared with the other groups (p<0.05) (Fig. 1b).
Oxidative stress parameters: The SOD activities were higher in the LPS group than in the other groups (p<0.05) (Fig. 2a).
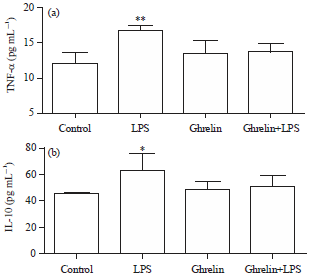 | |
| Fig. 1(a-b): | (a) TNF-α values in experimental groups. **Significant differences at p<0.01, LPS vs control and ghrelin groups and (b) IL-10 values in experimental groups, *Significant diffirences at p<0.05, LPS vs other groups |
| Table 1: | Biochemical parameters of serum glucose, AST and ALT for all groups |
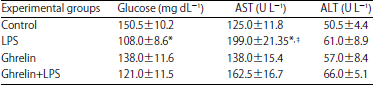 | |
| *p<0.01 vs control, ‡p<0.01 vs ghrelin | |
The TBARS values were increased in the LPS group compared with other groups (p<0.05) (Fig. 2b).
Ghrelin results: Ghrelin levels were significantly higher in the LPS and ghrelin groups than in the control group (p<0.05) (Fig. 2c).
Hematological and biochemical results: There were no differences in ALT levels in all experimental groups (p>0.05). There were highly significant differences in AST levels between the control and the LPS groups (p<0.01). Glucose levels were found highly significant in the LPS group compared with the control group (p<0.01) (Table 1).
The leukocyte numbers were increased in the LPS groups compared with all other groups (p<0.01) (Fig. 2d).
Histologic results: In the control group, there were normal liver parenchyma, hepatocytes were located radially around the central vein. In the sections of the LPS group, there were intense necrotic areas and were seen enlargements in sinusoids.
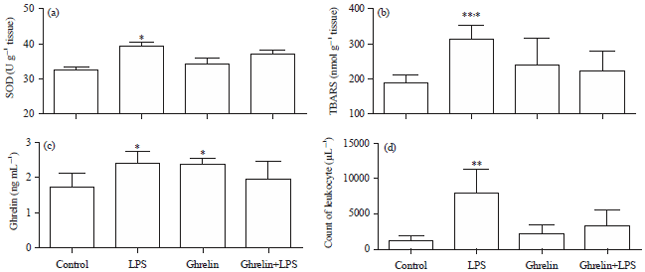 | |
| Fig. 2(a-d): | (a) SOD activities in experimental groups, *Significant diffirences at p<0.05, LPS vs other groups, (b) TBARS values in experimental groups, **Significant diffirences at p<0.01, LPS vs control and ghrelin+LPS groups, *Significant diffirences at p<0.05 LPS vs ghrelin groups, (c) Ghrelin values in experimental groups, *Significant differences at p<0.05, LPS and ghrelin vs control and ghrelin+LPS groups and (d) The leukocyte counts in experimental groups, **Significant diffirences at p<0.01, LPS vs other groups |
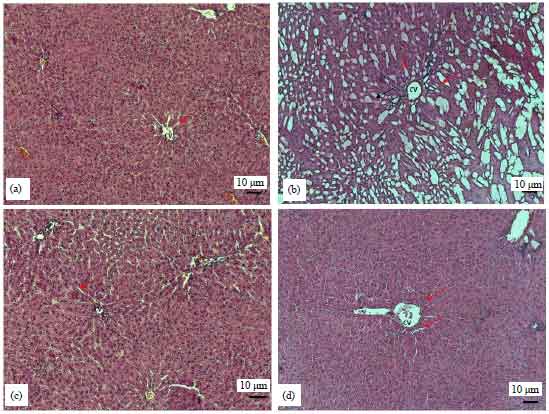 | |
| Fig. 3(a-d): | Section of liver tissue stained with H and E ×10 magnification, (a) Section of liver tissue from control group, (b) Section of liver tissue from LPS group, (c) Section of liver tissue from ghrelin group and (d) Section of liver tissue from ghrelin+LPS group, CV: Central vein. Arrows mentioned in enlargement of sinusoidal area |
In the ghrelin group, periportal areas in the region of the vena centralis had a neat appearance. Sinusoids and sinusoidal cells were the same as the controls. In the ghrelin+LPS group, the same image was seen as in the controls, regarding the sinusoids between hepatocytes cell cords and the periportal areas (Fig. 3a-d).
DISCUSSION
In this study, the effects of exogenous ghrelin on TNF-α, IL-10, SOD, TBARS and ghrelin levels were researched after inducing sepsis with lipopolysaccharide (LPS) in liver tissue. Exogenous ghrelin (10 nmol kg–1 i.v.) in the LPS-treated group was to determine which ghrelin had effects on inflammation, oxidative stress parameters and histologic structure in liver tissue.
In the LPS groups, the leukocyte numbers were significantly increased. The activation and accumulation of leukocytes in acute sepsis models have been shown to cause tissue damage in the liver due to endotoxin infusion with Gram-negative bacteria15,18,19.
There were found significantly increased levels of pro-inflammatory cytokine TNF-α and anti-inflammatory cytokine IL-10 in the LPS group compared with the controls, ghrelin and ghrelin+LPS groups. Consistent with our result, the increased amounts of cytokines in lipopolysaccharide induced sepsis has been demonstrated to cause metabolic and cellular changes in tissues, such as liver, kidney and intestine2-4.
Our findings of TNF-α levels in the ghrelin treated LPS group supported the results of other studies. In this study, GHRP-2, a synthetic analog of ghrelin had the effect of decreasing the inflammatory response to LPS-induced liver injury20. Wang et al.21 demonstrated that exogenous ghrelin was significantly decreased TNF-α levels in the acute kidney injury during endotoxemia by LPS administration in mice.
Decreased levels of IL-10 were observed in the ghrelin treated LPS groups depending on the dose and time that Ghrelin was given. Prenzler et al.22 showed increased levels of IL-10 in ghrelin treated normal weight endotoxemic rats but IL-10 levels decreased in obese rats.
It is known that IL-10 synthesis is important to maintain cytokine homeostasis in endotoxemia, which is characterized by dysregulation of pro and anti-inflammatory cytokine balances. The mechanism of ghrelin’s on IL-10 release is not clear. It is suggested that this decrement of IL-10 occured via its receptors on monocytes and macrophage in the ghrelin treated LPS group. Further studies are needed to clarify the effects of ghrelin on anti-inflammatory cytokine release in sepsis22.
In this study, SOD activities and TBARS levels were also examined as oxidative stress parameters. The activities of SOD and levels of TBARS in liver tissue were to increased in the LPS group. It was found that ghrelin treatment reduced SOD activities and TBARS levels.
Reactive Oxygen Species (ROS) are very important in the development of sepsis and organ damage23-25. Guerreiro et al.24 showed an increment in SOD activity following sepsis. In this study, TBARS is one of the indicators of lipid peroxidation, which is responsible for organ and tissue damage formed during sepsis with SOD23,26. Andrades et al.25 reported in their study that SOD/CAT and TBARS activities were related with organ damage development in sepsis.
Levels of TBARS have been reported to decrease in sepsis with treatment of exogenous ghrelin14,15,26. Previous studies reported that ghrelin has an antioxidant effect that decreases levels of TBARS and increases glutathione in organs, such as liver, heart and pancreas15,27. El Eter et al.28 showed in their in vitro and in vivo studies that the antioxidant activity of ghrelin reduced ROS production in parallel with our TBARS findings. Contrary to this, Li et al.14 reported that ghrelin had no effect on the decrement of ROS in cells.
Ghrelin levels were significantly increased in the LPS group. In addition, ghrelin levels were to approached that of the controls the ghrelin treated septic (Ghrelin+LPS) group. Consistent with our study, Granado et al.20 reported the protective effect of GHRP-2 on LPS-induced liver dysfunstion. Recent studies showed that ghrelin was released as an endogenous response owing to its tissue protective effects14,20.
Hepatic function may be affected to varying degrees in several infectious diseases. Aminotransferases have been shown to increase in diseases, such as ischemic liver injury, myocardial infarction, arrhythmias, congestive heart failure, stroke, sepsis and severe pathologic conditions23,29,30. There were found to increase ALT levels in the experimental group but these increments were not significant. The AST levels were significantly high in the LPS group compared with the other groups. In line with our result, Zapelini et al.31 reported that increased levels of ROS stimulated oxidative damage of lipid and protein macromolecules in early-stage sepsis and also increased AST levels in late stage sepsis.
Recent studies found that LPS decreased glucose levels in rats with sepsis. In this study, glucose levels were found highly decreased in the LPS group, but there were in the ghrelin-treated LPS group, glucose levels were found increased as same as control. Ghrelin improves serum glucose levels by increasing growth hormone levels in sepsis14,15,32,33.
Histopathologic changes have been shown in the liver of rats with sepsis34,35. In the present study, in histochemical evaluation of tissue sections was undertaken in all groups. There were intense necrotic areas and seen enlargements in sinusoids in the LPS group. In the ghrelin+LPS group, the histologic structure was found similar to the control group, regarding the sinusoids between hepatocytes cell cords and the periportal areas. In the ghrelin+LPS group, the same image was seen as in the controls, regarding the sinusoids between hepatocytes cell cords and the periportal areas.
CONCLUSION
In conclusion, ghrelin treatment attenuates pro-inflammatory cytokine TNF-α, TBARS levels, SOD activity and also impaired liver tissue damage in a time and dose dependent manner during sepsis.
ACKNOWLEDGMENT
This study was granted from Istanbul University Thesis Projects (Project No. 38846). We would like to thank Mr. David F. Chapman for editing the English of the manuscript.
REFERENCES
- Lever, A. and I. Mackenzie, 2007. Sepsis: Definition, epidemiology and diagnosis. Br. Med. J., 335: 879-883.
CrossRefPubMedDirect Link - Angus, D.C. and T. van der Poll, 2013. Severe sepsis and septic shock. N. Engl. J. Med., 369: 840-851.
CrossRefDirect Link - Ramachandran, G., 2014. Gram-positive and gram-negative bacterial toxins in sepsis: A brief review. Virulence, 5: 213-218.
CrossRefDirect Link - Aruoma, O.I., 1998. Free radicals, oxidative stress and antioxidants in human health and disease. J. Am. Oil Chem. Soc., 75: 199-212.
CrossRefDirect Link - Zimmerman, J.J., 1995. Defining the role of oxyradicals in the pathogenesis of sepsis. Crit. Care Med., 23: 616-617.
PubMedDirect Link - Peralta, J.G., S. Llesuy, P. Evelson, M.C. Carreras, B.G. Flecha and J.J. Poderoso, 1993. Oxidative stress in skeletal muscle during sepsis in rats. Circulat. Shock, 39: 153-159.
Direct Link - Choumar, A., A. Tarhuni, P. Letteron, F. Reyl-Desmars and N. Dauhoo et al., 2011. Lipopolysaccharide-induced mitochondrial DNA depletion. Antioxidants Redox Signal., 15: 2837-2854.
CrossRefDirect Link - Goode, H.F., H.C. Cowley, B.E. Walker, P.D. Howdle and N.R. Webster, 1995. Decreased antioxidant status and increased lipid peroxidation in patients with septic shock and secondary organ dysfunction. Crit. Care Med., 23: 646-651.
PubMed - Kojima, M., H. Hosoda, Y. Date, M. Nakazato, H. Matsuo and K. Kangawa, 1999. Ghrelin is a growth-hormone-releasing acylated peptide from stomach. Nature, 402: 656-660.
CrossRefPubMedDirect Link - Sato, T., Y. Nakamura, Y. Shiimura, H. Ohgusu, K. Kangawa and M. Kojima, 2012. Structure, regulation and function of ghrelin. J. Biochem., 151: 119-128.
CrossRefDirect Link - Majchrzak, K., K.M. Pawlowski, E.J. Orzechowska, I. Dolka, J. Mucha, T. Motyl and M. Krol, 2012. A role of ghrelin in canine mammary carcinoma cells proliferation, apoptosis and migration. BMC Vet. Res., Vol. 8.
CrossRefDirect Link - Wu, R., W. Dong, X. Cui, M. Zhou, H.H. Simms, T.S. Ravikumar and P. Wang, 2007. Ghrelin down-regulates proinflammatory cytokines in sepsis through activation of the vagus nerve. Ann. Surg., 245: 480-486.
CrossRefDirect Link - Li, W.G., D. Gavrila, X. Liu, L. Wang and S. Gunnlaugsson et al., 2004. Ghrelin inhibits proinflammatory responses and nuclear factor-κB activation in human endothelial cells. Circulation, 109: 2221-2226.
CrossRefPubMedDirect Link - Chang, L., J. Zhao, J. Yang, Z. Zhang, J. Du and C. Tang, 2003. Therapeutic effects of ghrelin on endotoxic shock in rats. Eur. J. Pharmacol., 473: 171-176.
CrossRefDirect Link - Suzuki, H., J. Matsuzaki and T. Hibi, 2011. Ghrelin and oxidative stress in gastrointestinal tract. J. Clin. Biochem. Nutr., 48: 122-125.
CrossRefDirect Link - Ohkawa, H., N. Ohishi and K. Yagi, 1979. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem., 95: 351-358.
CrossRefPubMedDirect Link - Rim, K.P., K. Kim, Y.H. Jo, J.H. Lee and J.E. Rhee et al., 2012. Effect of therapeutic hypothermia according to severity of sepsis in a septic rat model. Cytokine, 60: 755-761.
CrossRefDirect Link - Ozkok, E., H. Yorulmaz, G. Ates, P. Serdaroglu-Oflazer and A.S. Tamer, 2014. Effects of prior treatment with simvastatin on skeletal muscle structure and mitochondrial enzyme activities during early phases of sepsis. Int. J. Clin. Exp. Pathol., 7: 8356-8365.
PubMedDirect Link - Granado, M., A.I. Martin, M. Lopez-Mendiom, A. Lopez-Calderon and M.A. Villanua, 2008. GH-releasing peptide-2 administration prevents liver inflammatory response in endotoxemia. Am. J. Physiol.-Endocrinol. Metab., 294: E131-E141.
CrossRefPubMedDirect Link - Wang, W., S. Bansal, S. Falk, D. Ljubanovic and R. Schrier, 2009. Ghrelin protects mice against endotoxemia-induced acute kidney injury. Am. J. Physiol.-Renal Physiol., 297: F1032-F1037.
CrossRefDirect Link - Prenzler, N.K., C. Macke, R. Horn, G. Brabant, R. Pabst, M. Richter and H. Nave, 2007. Obesity influences the food consumption and cytokine pattern in ghrelin-treated endotoxemic rats. Life Sci., 81: 80-87.
CrossRefDirect Link - Ware, L.B., J.P. Fessel, A.K. May and L.J. Roberts, 2011. Plasma biomarkers of oxidant stress and development of organ failure in severe sepsis. Shock, 36: 12-17.
CrossRefDirect Link - Guerreiro, M.O., F. Petronilho, M. Andrades, L. Constantino and F.G. Mina et al., 2010. Plasma superoxide dismutase activity and mortality in patients with septic. J. Trauma Acute Care Surg., 69: E102-E106.
CrossRefDirect Link - Andrades, M., C. Ritter, M.R. de Oliveira, E.L. Streck, J.C.F. Moreira and F. Dal-Pizzol, 2011. Antioxidant treatment reverses organ failure in rat model of sepsis: Role of antioxidant enzymes imbalance, neutrophil infiltration and oxidative stress. J. Surg. Res., 167: e307-e313.
CrossRefDirect Link - Obay, B.D., E. Tasdemir, C. Tumer, H.M. Bilgin and M. Atmaca, 2008. Dose dependent effects of ghrelin on pentylenetetrazole-induced oxidative stress in a rat seizure model. Peptides, 29: 448-455.
CrossRefDirect Link - Nishi, Y., H. Mifune, A. Yabuki, Y. Tajiri and R. Hirata et al., 2013. Changes in subcellular distribution of N-octanoyl or N-decanoyl ghrelin in ghrelin-producing cells. Front. Endocrinol., Vol. 9.
CrossRefDirect Link - El Eter, E., A. Al Tuwaijiri, H. Hagar and M. Arafa, 2007. In vivo and in vitro antioxidant activity of ghrelin: Attenuation of gastric ischemic injury in the rat. J. Gastroenterol. Hepatol., 22: 1791-1799.
CrossRefDirect Link - Mathur, T., A.M. Manadan, S. Thiagarajan, B. Hota and J.A. Block, 2014. Serum transaminases are frequently elevated at time of diagnosis of idiopathic inflammatory myopathy and normalize with creatine kinase. J. Clin. Rheumatol., 20: 130-132.
CrossRefDirect Link - Sette, L.H.B.C. and E.P. de Almeida Lopes, 2014. Liver enzymes serum levels in patients with chronic kidney disease on hemodialysis: A comprehensive review. Clinics, 69: 271-278.
CrossRefDirect Link - Zapelini, P.H., G.T. Rezin, M.R. Cardoso, C. Ritter and F. Klamt et al., 2008. Antioxidant treatment reverses mitochondrial dysfunction in a sepsis animal model. Mitochondrion, 8: 211-218.
CrossRefDirect Link - Viana, M.V., R.B. Moraes, A.R. Fabbrin, M.F. Santos and F. Gerchman, 2014. Assessment and treatment of hyperglycemia in critically ill patients. Revista Brasileira Terapia Intensiva, 26: 71-76.
CrossRefDirect Link - Decaro, M.H. and N.E. Vain, 2011. Hyperglycaemia in preterm neonates: What to know, what to do. Early Hum. Dev., 87: S19-S22.
CrossRefDirect Link - Yang, S., M. Zhou, D.J. Koo, I.H. Chaudry and P. Wang, 1999. Pentoxifylline prevents the transition from the hyperdynamic to hypodynamic response during sepsis. Am. J. Physiol.-Heart Circ. Physiol., 277: H1036-H1044.
Direct Link - Thiemermann, C., H. Ruetten, C.C. Wu and J.R. Vane, 1995. The multiple organ dysfunction syndrome caused by endotoxin in the rat: attenuation of liver dysfunction by inhibitors of nitric oxide synthase. Br. J. Pharmacol., 116: 2845-2851.
CrossRefDirect Link