
Guo-Feng Fang
College of Animal Science and Technology, Shandong Agricultural University, Tai`an, 271018 Shandong, People`s Republic of China
Wei Chen
College of Animal Science and Technology, Shandong Agricultural University, Tai`an, 271018 Shandong, People`s Republic of China
Shou-Dong Wang
College of Animal Science and Technology, Shandong Agricultural University, Tai`an, 271018 Shandong, People`s Republic of China
Yan-Dong Wang
College of Animal Science and Technology, Shandong Agricultural University, Tai`an, 271018 Shandong, People`s Republic of China
Chuan-Hao Li
College of Animal Science and Technology, Shandong Agricultural University, Tai`an, 271018 Shandong, People`s Republic of China
Hong-Lei Zhu
College of Animal Science and Technology, Shandong Agricultural University, Tai`an, 271018 Shandong, People`s Republic of China
Hui Wang
College of Animal Science and Technology, Shandong Agricultural University, Tai`an, 271018 Shandong, People`s Republic of China
Yong-Qing Zeng
College of Animal Science and Technology, Shandong Agricultural University, Tai`an, 271018 Shandong, People`s Republic of China
Asian Journal of Animal and Veterinary Advances
Year: 2017 | Volume: 12 | Issue: 3 | Page No.: 161-168







ABSTRACT
Background: Intramuscular fat (IMF) is one of the most important traits affecting meat quality such as tenderness, juiciness and flavor. Phosphotyrosine interaction domain containing 1 (PID1) gene has been reported as a candidate gene for fat deposition. Therefore, an overexpression of PID1 gene in pigs could be of great benefit to the swine industry and consumers. Materials and Methods: In this study, the PID1-transgenic pigs were produced by sperm mediated gene transfer and magnetic nanoparticles with high efficiency. Real-time fluorescence quantitative PCR and Southern blot methods were employed to identify PID1-transgenic pigs. The copy numbers of exogenous gene in transgenic pigs were estimated by Southern blot analysis. Results: Twenty four live piglets were born from the two sows by natural delivery. Eleven piglets were positive in the PCR amplifications (45.83%). These PCR-positive pigs were further identified by Southern-blot analysis, out of which 7 pigs were positive in both Southern blotting and PCR (29.16%), others were positive only in PCR. In addition, the PID1 expression levels in the liver and longissimus dorsi muscle tissue were higher than wild-type ones (p<0.05), whereas the levels in backfat and kidney were similar to wild-type ones in transgenic pigs. Conclusion: The results suggest that magnetic nanoparticles can be used to efficiently introduce a transgene into animals via sperm mediated gene transfer, the PID1-transgenic pig model could further serve to study obesity and the mechanism of IMF deposition.
PDF Abstract XML References Citation
How to cite this article
Guo-Feng Fang, Wei Chen, Shou-Dong Wang, Yan-Dong Wang, Chuan-Hao Li, Hong-Lei Zhu, Hui Wang and Yong-Qing Zeng, 2017. Generation of Transgenic Pigs Overexpressing PID1 Gene
Mediated by Magnetic Nanoparticles and Sperm. Asian Journal of Animal and Veterinary Advances, 12: 161-168.
DOI: 10.3923/ajava.2017.161.168
URL: https://scialert.net/abstract/?doi=ajava.2017.161.168
DOI: 10.3923/ajava.2017.161.168
URL: https://scialert.net/abstract/?doi=ajava.2017.161.168
INTRODUCTION
Pork is the most widely consumed meat accounting for more than half of meat consumption. With the global economy growth, the intensive swine production industry has been employed to meet the demand for a faster and larger growth of pigs with a high lean meat content. However, this type of production has brought problems related to poor meat quality and has decreased the intramuscular fat (IMF) content. Meat quality is an important economic trait in domestic animals. Improving the quality of fresh pork is significant importance to swine industry. The IMF also known as marbling has been revealed to be one of the most important traits affecting meat quality, such as meat tenderness, juiciness and taste1-3.
Phosphotyrosine interaction domain containing 1 (PID1) is an important mediator in the development of obesity-related insulin resistance (IR) in humans and animals. The PID1 gene has recently been discovered and reported that there was a higher level in obese subjects4. An increased expression of PID1 in 3T3-L1 adipocytes reduced insulin-stimulated glucose uptake and impaired insulin-stimulated PID1 translocation5. Further study has found that α-lipoic acid protects 3T3-L1 adipocytes from PID1-induced IR partially through increasing phosphorylation of Insulin receptor substrate 1 (IRS-1) and provide evidence that PID1 may be a potential target for the treatment of obesity and obesity-related IR6. A preliminary study of pig PID1 gene indicate that the expression of PID1 in laiwu pigs which have high level IMF content (12.78%) were much higher than large white pigs, implied that the expression of PID1 may be related to the IMF deposition7. All these results indicate that PID1 may be a potential candidate gene for fat deposition.
With the advances in biotechnology, transgenic techniques have been widely developed, which have become an essential tool in breeding genetically modified animals for research, agricultural production and disease resistance8-12. The previous methods for producing the transgenic animals have limitations such as the high cost of equipment’s, the need for skilled researchers and low efficiency. Therefore, new methods are still need to increase efficiency of transgenesis (gene integration and expression) with minimal host immune response. Nanotechnology describes the utilization of materials and systems through control of nanometer-sized materials and their application to physics, biology, medicine and other endeavors13. Magnetic nanoparticles (MNPs) have been studied extensively for the purpose of delivering drugs or genes into target cells or tissues14. Magnetofection is a simple, versatile and highly efficient method of gene transfer15-17. The basic principle of magnetofection is to associate MNPs with a transgene or virus, so that the complexes can be delivered into target cells using a magnetic field. This approach has a great advantage of significantly increasing the expression of a transgene into target cell, while reducing overall cell damage.
Here, the polyMAG-1000, polyethylenimine (PEI)-coated MNPs were used as a gene delivery agent to sperm cells. After that, the infected sperms were used as a carrier for transgene delivery. A transgenic pig model overexpression PID1 under the control of the cytomegalovirus (CMV) enhancer was obtained and examined.
MATERIALS AND METHODS
Construction of pIRES2-AcGFP1-PID1: The transgene plasmids, pIRES2-AcGFP1-PID1, which is constructed by Zhu et al.18 were expressed in Escherichia coli and then isolated and purified by an Endo Free Maxi Plasmid Kit (Tiangen, Beijing, China). It contains the aequorea coerulescens green fluorescent protein 1 (AcGFP1) reporter gene, cytomegalovirus (CMV) enhancer and phosphotyrosine interaction domain containing 1 (PID1).
Generation of transgenic pig: A large white boar and three large white sows (all were in their oestrum) were used as the experimental animals. Semen was collected from trained boar that had abstained for 3 days. The plasmids were diluted in autoclaved distilled water to get a final concentration of 800 mg mL–1 for polyMAG-1000 (Chemicell, Germany). About 500 μL of 800 mg mL–1 plasmids solution and an equal volume (500 μL) of polyMAG-1000 were added into the same eppendorf tube. The plasmids/nanoparticles mixture was gently pipetted up and down about five times and incubated at room temperature for 10 min. Then 1 mL mixture was added to 80 mL of semen (2×109 spermatozoa) in a 75 cm2 cell culture flask (Corning, USA). Sperm motility was calculated to be 0.8 before incubated with the plasmids/nanoparticles mixture. The sperm/plasmids/nanoparticles mixture was gently pipetted up and down about 10 times. After that, the flask was then placed on the MagnetoFACTOR-24 (Chemicell, Germany), incubated at room temperature for 30 min. Artificial insemination (80 mL sperm/plasmids/nanoparticles mixture/ sow) was performed and each sow was inseminated with 80 mL sperm/plasmids/nanoparticles mixture again the next day. Pregnancy was checked regularly about 21 days after artificial insemination using abdominal ultrasound examination. Piglets were delivered by natural birth after 115 days. The experimental protocols were conducted in compliance with Chinese guidelines for human care and use of animals in research.
Genotyping of transgenic pig: Piglets were born by natural delivery, which were subjected to PCR to confirm the presence of the transgene. The genomic DNA was extracted from 2 days old pig ear biopsies using a TIANamp Genomic DNA Kit (Tiangen, Beijing, China) following instructions provided in the attached protocol. Two pairs of primers were used for amplification of AcGFP1 and PID1, respectively. A 310 bp fragment of AcGFP1 was amplified with specific primer GFP-f (5'-CGAGGATGACGGCAACT-3') and primer GFP-r (5'-GGTGGACAGGTAGTGGTTATC-3'). The PCR was performed with the following parameters: 94°C for 5 min, followed by 35 cycles of 30 sec at 94°C, 20 sec at 61°C and 20 sec at 72°C, with a final extension step at 72°C for 5 min. A 247 bp fragment of PID1 was amplified with specific primer PID1-f (5'-TGGCTCGCATCGCCTACT-3') and primer PID1-r (5'-AACTCCTGGGACACCTCTT-3'). The PCR program initially started with a 94°C denaturation for 5 min, followed by 35 cycles of 30 sec at 94°C, 20 sec at 58°C and 20 sec at 72°C, then 72°C extension for 5 min. About 10 μL PCR products were electrophoresed through 1% agarose gel, stained with ethidium bromide and photographed by the JY04S-3B electrophoresis equipment (Junyi-dongfang, China). The amplified products were purified using a Universal DNA Purification Kit (Tiangen, China) and sequenced to confirm their identities.
Southern blot analysis: The PCR positive pigs were further confirmed by Southern blot analysis using DIG-High Prime DNA Labeling and Detection Starter Kit I (Roche, Germany) and DIG Wash and Block Buffer Set (Roche, Germany) as described in manuals. Genomic DNAs (20 μg) were digested with EcoR I (Thermo, USA) overnight and subjected to electrophoresis in 1% agarose gel with 40 V. The DNA was then transferred to a positively charged nylon membrane (Solarbio, China) by the capillary transfer method. During transfer, the DNA was nicked and denatured. Then the nylon membrane was pre-hybridized at 42°C for 1 h and then hybridized at 65°C overnight with a 663 bp PID1 probe. The probe was amplified using the primer pairs PID1p-f (5'-GGCTGGAAGATGTGGCAG-3') and PID1p-r (5'-TCAGCCATCATCGGATTCT-3') and labeled with digoxigenin. After hybridization, the hybridized probes were immunodetected with anti-digoxigenin-AP and fab fragments were then visualized with the colorimetric substrates NBT/BCIP.
Real-time fluorescence quantitative PCR: Two transgenic pigs, No. 12 and 14 and two control wild-type pigs were sacrificed when animals weighted 100±2 kg to investigate the functional expression of PID1 gene. Both the transgenic and wild-type pigs were fed with the same commercial standard diets in the same farm and reared in collective pens. Liver, backfat, longissimus dorsi muscle (LD) and kidney samples were harvested and frozen in liquid nitrogen jar for total RNA and total protein extraction. Part of LD muscle tissue of each pig was stored at 4°C for 24 h for IMF content measurement. Total RNA was isolated from triturated tissues of transgenic and wild-type pigs using trizol reagent (Invitrogen, USA) according to the manufacturer’s instructions. To remove genomic DNA contamination, total RNA was digested with DNase I, RNase-free (Thermo, USA). The concentration and quantification of RNA was estimated by BioPhotometer Plus (Eppendorf, Germany) and 1% agarose gel electrophoresis stained with ethidium bromide. Then, total RNA was reverse transcribed to cDNA according to the manufacturer’s instructions of PrimeScript RT reagent Kit with gDNA Eraser (TaKaRa, Dalian, China). Analysis of tissue-specific expression patterns of PID1 was performed by real-time fluorescence quantitative PCR (qRT-PCR) using the housekeeping gene GAPDH as an internal control. Primers for PID1 were PID1Q-F (5'-ATGTCTTTCCAGCCAATG-3') and PID1Q-R (5'-TAATCTCCCTGTAAACCCA-3'). The GAPDH was amplified using GAPDH-F (5'-ACTCACTCTTCTACCTTTGAT GCT-3') and GAPDH-R (5'-TGTTGCTGTAGCCAAATTCA-3')19. The qPCR amplification was conducted using an Mx3000P Real-Time PCR System (Stratagene, USA) and performed in a three repeats for each sample. Each reaction contains 7.5 μL 2x SYBR Premix Ex Taq (TaKaRa, Dalian, China), 0.3 μL 50x ROX Reference Dye II, 0.3 μL forward and reverse primers (10 μmol L–1), respectively, 1.5 μL cDNA and complementary water, to a final volume of 15 μL. The PCR amplification was carried out as follows: 95°C for 5 sec, then 40 cycles of 95°C for 5 sec, 59°C for 10 sec, 72°C for 15 sec, followed by 1 cycles of 95°C for 1 min, 61°C for 30 sec, 95°C for 30 sec to calculate the melting curve. The 2–ΔΔCt method was used to analyze the relative expression levels of PID1 gene20.
Western blot analysis: Total protein from tissues was extracted by homogenization in ice-cold RIPA buffer (Beyotime, China) supplemented with PMSF as protease inhibitor (Beyotime, China) for 2 h at 4°C. After centrifugation at 12,000xg for 20 min at 4°C, the supernatant was quantified using the Enhanced BCA Protein Assay Kit (Beyotime, China) in accordance with the manufacturer’s instructions. Equal amount of protein (10 μg lane–1) was separated by electrophoresis on 12% SDS-PAGE gels and transferred onto PVDF membranes (Solarbio, China) using the standard wet-tank transfer method (Junyi-Dongfang, China). After that, the membrane was blocked for 1 h at room temperature with blocking buffer (Beyotime, China). In order to detect the AcGFP1 protein, the LD muscle tissue protein on the PVDF membrane was hybridized with rabbit anti-GFP antibodies (1:1,000; Beyotime, China) and goat anti-rabbit IgG (1:1,000; Beyotime, China) were used as secondary antibodies. Finally, the target protein AcGFP was stained by TMB (Tiangen, China). For immunoblotting of PID1 and GAPDH (internal control) proteins, the membrane was cut into two sections. The top membrane section (size between 43 and 72 kDa) was incubated with rabbit anti-PID1 antibodies (1:1,000; Abcam, USA), the other membrane section (size between 10 and 43 kDa) was incubated with mouse anti-GAPDH antibodies (1:1,000; Beyotime, China) at 4°C overnight. Membranes were then washed three times with TBST for 10 min and subsequently incubated with either horseradish peroxidase-conjugated goat anti-rabbit IgG or goat anti-mouse IgG (1:1,000; Beyotime, China) as the secondary antibodies for 1 h at room temperature. After washing three times with TBST, the blots of the membranes were developed using the BeyoECL Plus chemiluminescence detection system (Beyotime, China). The membranes were exposed to film for 30 sec after incubation with ECL reagents. Densitometry was performed using Image-Pro Plus software.
Measurement of IMF content: After thawing at 4°C for 24 h, adhering adipose and connective tissue were removed from LD muscle. Then IMF content was measured by soxhlet petroleum-ether extraction method and expressed as the weight percentage of wet muscle tissue.
Statistical analysis: All the data were expressed as Means±SEM. Statistical analysis was performed using Student’s t test. The threshold of significance was defined as a p<0.05.
RESULTS AND DISCUSSION
Figure 1 shows the expression vector based on the pIRES2 backbone, successfully constructed as revealed by restriction profiles and sequencing analysis. After incubation at room temperature for 30 min, artificial insemination (80 mL sperm/plasmids/nanoparticles mixture/sow) was performed and all the sows became pregnant and one had a spontaneous abortion. Twenty four live piglets were born from the two sows by natural delivery. Eleven piglets were positive in the PCR amplifications (45.83%) with 310 and 247 bp fragments (Fig. 2a). These PCR-positive pigs were further identified by Southern-blot analysis, out of which seven pigs were positive in both Southern blotting and PCR (29.16%) (Fig. 2b), others were positive only in PCR. Sperm Mediated Gene Transfer (SMGT) is one of many transgenic techniques, which does not require embryo handling or expensive equipment and has been used more or less successfully in production of transgenic embryos and animals in a large number of species21. The exogenous DNA interacts with DNA-binding proteins (DBPs) that are present on the surface of spermatozoa head22. Previous study demonstrated that DNA uptake of spermatozoa by magnetofection is more efficient compared to that of DNA alone, because MNPs can bind over the entire surface of the spermatozoa, while exogenous DNA only binds the DBPs on the spermatozoa head17,23,24.
To investigate the functional expression of PID1, qRT-PCR was conducted to examine the tissues (liver, backfat, LD muscle tissue and kidney) from transgenic pigs (No. 12 and 14) and two wild-type pigs. High expression levels of PID1 were detected in liver and LD muscle tissue (p<0.05) in transgenic pigs, whereas the mRNA levels in other tissues were no difference. The PID1 expression levels in liver and LD muscle tissue in transgenic pigs were higher than wild-type pigs (p<0.05), whereas the levels in backfat and kidney were similar to wild-type ones (Fig. 3a).
Western blot analysis was performed to identify transgenic pigs and evaluate the difference in PID1 protein expression levels in transgenic and wild-type pigs. Western blot analysis further supported the results of Southern blot analysis with the anti-GFP antibody revealed a single band of 27 kDa specific to the LD muscle tissue of transgenic pigs (No. 12 and 14) and no corresponding band in wild type ones (Fig. 3b). Assessment of PID1 expression by western blot showed results similar to qRT-PCR (Fig. 3c).
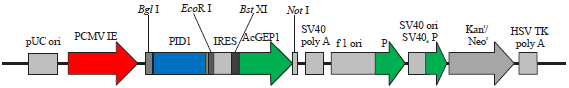 | |
| Fig. 1: | Schematic diagram of expression cassette of pIRES2-AcGFP1-PID1 vector |
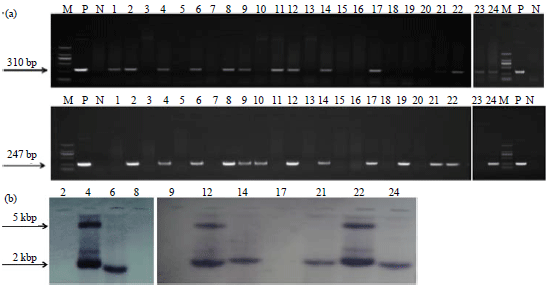 | |
| Fig. 2(a-b): | Identification of transgenic pigs by PCR and Southern blot, (a) The amplified product of AcGFP gene is 310 bp, the product of PID1 gene is 247 bp. Lane 1-24: Positive pig with two objective bands, negative pig with one or no objective band and (b) Identification of transgenic pigs by Southern blot. Genomic DNA of the PCR positive founders was digested with EcoR I and hybridized with a probe to detect the PID1 gene. M: DL2000 DNA marker, P: Positive plasmid control, N: Negative control using water as template |
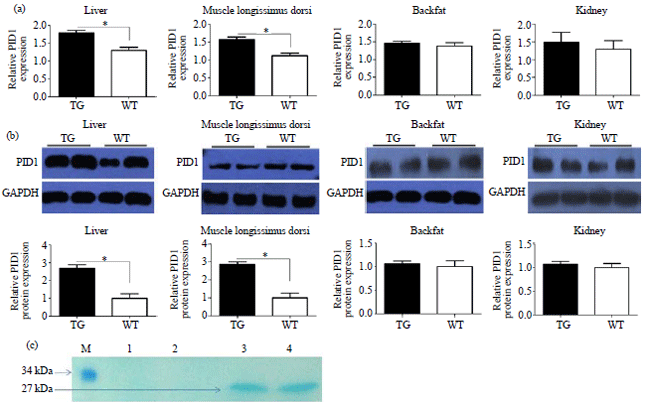 | |
| Fig. 3(a-c): | Expression of PID1 in transgenic pigs, (a) Analysis of relative transgene expression level in transgenic pigs and wild-type pigs by real-time fluorescence quantitative PCR. The GAPDH was used as an internal control. TG: Transgenic pigs, WT: Wild type pigs, *p<0.05, (b) Western blot analyses of PID1 using tissues collected from transgenic pigs and wild type pigs. Densitometry analysis of Western blot results were perform in triplicate (Date were presented as Mean±SEM). The GAPDH was used as an internal control. TG: Transgenic pigs, WT: Wild type pigs, *p<0.05 and (c) AcGFP protein analysis of the transgenic pigs. M: Prestained dual color protein molecular weight marker, lane 1 and 2: Wild-type pigs, lane 3: Transgenic pigs No. 12, lane 4: Transgenic pigs No. 14. A 27 kDa band of AcGFP is apparent in the LD muscle of transgenic pigs but no corresponding band in wild type pigs |
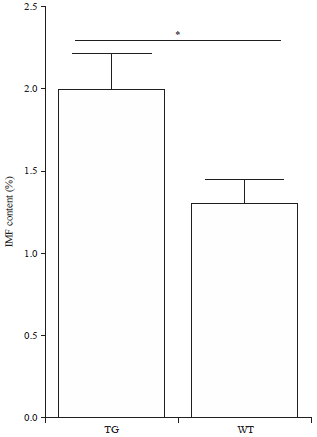 | |
| Fig. 4: | IMF content in the LD muscle of transgenic pigs and wild type pigs. TG: Transgenic pigs, WT: Wild type pigs. All assays were performed in triplicate and the results were given as Mean±SEM. *p<0.05 |
Figure 4 shows the IMF content was increased in transgenic pigs and it was higher than that in the wild type pigs.
Previous study of the PID1 silenced transgenic rabbits which was constructed by RNA interference showed that the mRNA and protein level of PID1 in the positive rabbits decreased significantly compared with the negative and blank groups25. The PID1 knockdown partly rescued the dissipation of mitochondrial mass, mitochondrial DNA and intracellular ATP synthesis occurred following the addition of cyanide p-trifluoromethoxyphenyl-hydrazone (FCCP) as well as inhibition of mitochondrial transmembrane potential in 3T3-L1 adipocytes incubated with FCCP, which might explain the involvement of PID1 in development of obesity-associated IR and mitochondrion function26. Further study showed that PID1 knockdown dramatically activate the insulin-stimulated phosphorylation of IRS-1, increases glucose transport in myocytes by activation of the IRS-1/PI3K/AKT insulin pathway27. In this study, qRT-PCR and western blot indicated that PID1 was up-regulated in transgenic pigs and the mRNA levels of PID1 of the transgenic pigs were much higher in liver than in other tissues. The high expression level of PID1 in pig liver indicates that the PID1 may be involved in lipid metabolism. No matter what the RNA level or protein level, the expression of exogenous gene-PID1 and the IMF content did not achieve the ideal of a higher level. This may be consistent with studies that have reported that foreign gene expression may be influenced by the copy number of the transgene28,29, transgene integration site and form of integration in the transgenic animal’s genome30,31. All these elements probably have an influence on the expression of the transgene in this study. These Southern-positive transgenic pigs showed one to three bands that hybridized with 663 bp fragment probe, indicating that copy number of exogenous gene in transgenic positive pigs is 1-3. The low IMF was likely caused by the low copy number of exogenous gene. In order to obtain an ideal efficiency expression of exogenous gene-PID1 and a higher IMF content, the transgene may should be integrated into the specific site in the genome with high copy numbers.
CONCLUSION
The PID1-transgenic pig model with increased transgene expression of PID1 could serve to develop transgenic pigs with higher IMF content and improve pork quality, which may also provide a good model to study obesity and the mechanism of IMF deposition.
ACKNOWLEDGMENTS
Both Guo-Feng Fang and Wei Chen contributed equally to this study and this study was supported by the National Animal Breed Resource Preservation Project of China (2130135), the National Project for Breeding Transgenic Pigs of China (2013ZX08006-002), Shandong Provincial Modern Pig Technology and Industry System Project (SDAIT-08-02) and Shandong Provincial Agricultural Animal Breeding Project of China (2013LZ02-015, 2014LZ03-016).
REFERENCES
- Fernandez, X., G. Monin, A. Talmant, J. Mourot and B. Lebret, 1999. Influence of intramuscular fat content on the quality of pig meat-1. Composition of the lipid fraction and sensory characteristics of m. longissimus lumborum. Meat Sci., 53: 59-65.
CrossRefDirect Link - Fortin, A., W.M. Robertson and A.K.W. Tong, 2005. The eating quality of Canadian pork and its relationship with intramuscular fat. Meat Sci., 69: 297-305.
CrossRefDirect Link - Cannata, S., T.E. Engle, S.J. Moeller, H.N. Zerby and A.E. Radunz et al., 2010. Effect of visual marbling on sensory properties and quality traits of pork loin. Meat Sci., 85: 428-434.
CrossRefDirect Link - Wang, B., M. Zhang, Y.H. Ni, F. Liu and H.Q. Fan et al., 2006. Identification and characterization of NYGGF4, a novel gene containing a phosphotyrosine-binding (PTB) domain that stimulates 3T3-L1 preadipocytes proliferation. Gene, 379: 132-140.
CrossRefPubMedDirect Link - Zhang, C.M., X.H. Chen, B. Wang, F. Liu and X. Chi et al., 2009. Over-expression of NYGGF4 inhibits glucose transport in 3T3-L1 adipocytes via attenuated phosphorylation of IRS-1 and Akt. Acta Pharmacol. Sin., 30: 120-124.
CrossRefPubMedDirect Link - Wang, Y.M., X.F. Lin, C.M. Shi, L. Lu and Z.Y. Qin et al., 2012. α-Lipoic acid protects 3T3-L1 adipocytes from NYGGF4 (PID1) overexpression-induced insulin resistance through increasing phosphorylation of IRS-1 and Akt. J. Bioenerg. Biomembr., 44: 357-363.
CrossRefDirect Link - Qian, Y., Y.Q. Zeng, J.F. Du, J.X. Cui and H. Li et al., 2010. CDS cloning and relationship between intramuscular fat content and mRNA expression of PID1 gene in pig. Hereditas, 32: 1153-1158.
PubMedDirect Link - Harvey, A.J., G. Speksnijder, L.R. Baugh, J.A. Morris and R. Ivarie, 2002. Expression of exogenous protein in the egg white of transgenic chickens Nat. Biotechnol., 20: 396-399.
CrossRefDirect Link - Webster, N.L., M. Forni, M.L. Bacci, R. Giovannoni and R. Razzini et al., 2005. Multi-transgenic pigs expressing three fluorescent proteins produced with high efficiency by sperm mediated gene transfer. Mol. Reprod. Dev., 72: 68-76.
CrossRefDirect Link - Cui, D., J. Li, L. Zhang, S. Liu and X. Wen et al., 2015. Generation of bi-transgenic pigs overexpressing human lactoferrin and lysozyme in milk. Trans. Res., 24: 365-373.
CrossRefDirect Link - Lin, Y.S., C.C. Yang, C.C. Hsu, J.T. Hsu, S.C. Wu, C.J. Lin and W.T.K. Cheng, 2015. Establishment of a novel, eco-friendly transgenic pig model using porcine pancreatic amylase promoter-driven fungal cellulase transgenes. Trans. Res., 24: 61-71.
CrossRefDirect Link - Cameron, E.R., 1997. Recent advances in transgenic technology. Mol. Biotechnol., 7: 253-265.
CrossRefDirect Link - Kami, D., S. Takeda, Y. Itakura, S. Gojo, M. Watanabe and M. Toyoda, 2011. Application of magnetic nanoparticles to gene delivery. Int. J. Mol. Sci., 12: 3705-3722.
CrossRefDirect Link - Scherer, F., M. Anton, U. Schillinger, J. Henke and C. Bergemann et al., 2002. Magnetofection: Enhancing and targeting gene delivery by magnetic force in vitro and in vivo. Gene Ther., 9: 102-109.
Direct Link - Wang, Y., H. Cui, K. Li, C. Sun and W. Du et al., 2014. A magnetic nanoparticle-based multiple-gene delivery system for transfection of porcine kidney cells. PloS ONE, Vol. 9.
CrossRefDirect Link - Farini, V.L., C.V. Camano, G. Ybarra, D.L. Viale, G. Vichera, J.S. Yakisich and M. Radrizzani, 2016. Improvement of bovine semen quality by removal of membrane-damaged sperm cells with DNA aptamers and magnetic nanoparticles. J. Biotechnol., 229: 33-41.
CrossRefDirect Link - Zaniboni, A., M. Spinaci, A. Zannoni, C. Bernardini, M. Forni and M.L. Bacci, 2016. X and Y chromosome-bearing spermatozoa are equally able to uptake and internalize exogenous DNA by sperm-mediated gene transfer in swine. Res. Vet. Sci., 104: 1-3.
CrossRefDirect Link - Zhu, H.L., Y. Shi, Y.Q. Zeng, W. Chen and Z.G. Xu et al., 2014. Generation of transgenic rabbits expressing swine PID1 by testicular injection. Acta Laborator. Anim. Scient. Sin., 22: 6-12.
Direct Link - Erkens, T., M. van Poucke, J. Vandesompele, K. Goossens, A. van Zeveren and L.J. Peelman, 2006. Development of a new set of reference genes for normalization of real-time RT-PCR data of porcine backfat and longissimus dorsi muscle and evaluation with PPARGC 1 A. Biol. Med. Chem. Biotechnol. Vol. 6.
CrossRefDirect Link - Livak, K.J. and T.D. Schmittgen, 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods, 25: 402-408.
CrossRefDirect Link - Simth, K. and C. Spadafora, 2005. Sperm-mediated gene transfer: Applications and implications. Bio. Essays, 27: 251-262.
CrossRefDirect Link - Zani, M., M. Lavitrano, D. French, V. Lulli, B. Maione, S. Sperandio and C. Spadafora, 1995. The mechanism of binding of exogenous DNA to sperm cells: Factors controlling the DNA uptake. Exp. Cell Res., 217: 57-64.
CrossRefDirect Link - Kim, T.S., S.H. Lee, G.T. Gang, Y.S. Lee and S.U. Kim et al., 2010. Exogenous DNA uptake of boar spermatozoa by a magnetic nanoparticle vector system. Reprod. Domestic Anim., 45: e201-e206.
CrossRefDirect Link - Hasanzadeh, R., M. Zandi, M.R. Sanjabi, K.H. Pajooh and H. Shabani, 2016. Expression of fluorescent reporter protein was not obtained in ovine embryos produced through in vitro fertilization-sperm mediated gene transfer (IVF-SMGT). Small Rumin. Res., 143: 24-28.
CrossRefDirect Link - Xu, Z.G., L. Yang, Y.Q. Zeng, Z. Zhang and W. Chen et al., 2014. Construction of transgenic rabbit model by RNA interferencing PID1 gene. Acta Vet. Zootech. Sin., 45: 750-756.
Direct Link - Shi, C.M., Y.M. Wang, C.M. Zhang, J. Qiu and Y.H. Shen et al., 2012. Knockdown of NYGGF4 (PID1) rescues insulin resistance and mitochondrial dysfunction induced by FCCP in 3T3-L1 adipocytes. Mitochondrion, 12: 600-606.
CrossRefDirect Link - Zeng, X.Q., C.M. Zhang, M.L. Tong, X. Chi and X.L. Li et al., 2012. Knockdown of NYGGF4 increases glucose transport in C2C12 mice skeletal myocytes by activation IRS-1/PI3K/AKT insulin pathway. J. Bioenerg. Biomembr., 44: 351-355.
CrossRefDirect Link - Rahman, M.A., G.L. Hwang, S. Razak, F. Sohm and N. Maclean, 2000. Copy number related transgene expression and mosaic somatic expression in hemizygous and homozygous transgenic tilapia (Oreochromis niloticus). Transgenic Res., 9: 417-427.
CrossRefDirect Link - Kong, Q., M. Wu, Y. Huan, L. Zhang and H. Liu et al., 2009. Transgene expression is associated with copy number and cytomegalovirus promoter methylation in transgenic pigs. PLoS ONE, Vol. 4.
CrossRefDirect Link - Williams, A., N. Harker, E. Ktistaki, H. Veiga-Fernandes and K. Roderick et al., 2008. Position effect variegation and imprinting of transgenes in lymphocytes. Nucl. Acids Res., 36: 2320-2329.
CrossRefPubMedDirect Link - McBurney, M.W., T. Mai, X. Yang and K. Jardine, 2002. Evidence for repeat-induced gene silencing in cultured Mammalian cells: Inactivation of tandem repeats of transfected genes. Exp. Cell Res., 274: 1-8.
CrossRefPubMedDirect Link