
B.C.J. De Silva
Veterinary Medical Center and College of Veterinary Medicine, Chungbuk National University, 28644 Cheongju, Korea
S.H.M.P. Wimalasena
Veterinary Medical Center and College of Veterinary Medicine, Chungbuk National University, 28644 Cheongju, Korea
Sabrina Hossain
Veterinary Medical Center and College of Veterinary Medicine, Chungbuk National University, 28644 Cheongju, Korea
H.N.K.S. Pathirana
Veterinary Medical Center and College of Veterinary Medicine, Chungbuk National University, 28644 Cheongju, Korea
Gang-Joon Heo
Veterinary Medical Center and College of Veterinary Medicine, Chungbuk National University, 28644 Cheongju, Korea
LiveDNA: 82.16116
Asian Journal of Animal and Veterinary Advances
Year: 2017 | Volume: 12 | Issue: 3 | Page No.: 152-160







ABSTRACT
Background and Objective: Infections of antibiotic-resistant bacteria in both human and animals own a great significance. The current study aimed to determine the quinolone susceptibility and the genetic characteristics of quinolone resistance of 20 strains of Pseudomonas aeruginosa isolated from pet Chinese stripe-necked turtles (Ocadia sinensis ). Methodology: Susceptibility of four antimicrobials including nalidixic acid, ciprofloxacin, ofloxacin and levofloxacin was examined. The PCR was carried out to amplify Quinolone Resistance Determining Region (QRDR) and to screen Plasmid Mediated Quinolone Resistance (PMQR) genes. Results: All tested isolates were resistant to nalidixic acid (100%), but none of the isolates show complete resistance to other tested antimicrobials. Two isolates showed intermediate resistance to ciprofloxacin (5%) and ofloxacin (5%), one each in Minimum Inhibitory Concentration (MIC) test. The qnrB gene was identified in one isolate (5%) and qnrS in three isolates (15%). The PCR assay could amplify aac(6’)-Ib gene from 8 isolates (40%) and none of them harbored aac(6’)-Ib-cr variant. Sequences obtained by amplifying gyrA and parC regions did not show any point mutation in QRDR. Neighbor-joining phylogenetic tree for gyrA indicated three distinct clads comprising first, current study isolates, second, clinical isolates of human and dogs and third, isolates from soil and water. Conclusion: All results suggest that studied strains of P. aeruginosa are less resistant to quinolones and are genetically more conserved with regards to gyrA gene region.
PDF Abstract XML References Citation
How to cite this article
B.C.J. De Silva, S.H.M.P. Wimalasena, Sabrina Hossain, H.N.K.S. Pathirana and Gang-Joon Heo, 2017. Characterization of Quinolone Resistance of Pseudomonas
aeruginosa Isolated from Pet Chinese Stripe-necked Turtles
(Ocadia sinensis). Asian Journal of Animal and Veterinary Advances, 12: 152-160.
DOI: 10.3923/ajava.2017.152.160
URL: https://scialert.net/abstract/?doi=ajava.2017.152.160
DOI: 10.3923/ajava.2017.152.160
URL: https://scialert.net/abstract/?doi=ajava.2017.152.160
INTRODUCTION
Ubiquitous environmental bacterium, Pseudomonas aeruginosa is a Gram-negative, aerobic bacterium which mostly thrives in moist or wet conditions. As a human opportunistic pathogen, P. aeruginosa causes infections ranging from minor gastrointestinal infections resulting in diarrhea, fever and headache to severe infections including meningitis, bacterial keratitis, endophthalmitis and necrotizing enterocolitis, especially in patients with impaired immune systems1. Despite Pseudomonas spp. is considered part of the indigenous flora of chelonian oral cavity and intestinal tract, they can cause opportunistic infections leading to ulcerative stomatitis, dermatitis, septicemia and pneumonia2.
Aminoglycosides and fluoroquinolones are commonly used for the systemic treatment of P. aeruginosa infections. However, resistance to these drugs is frequently detected in clinical practice3. The P. aeruginosa has been identified to possess an intrinsic antimicrobial resistance due to low outer membrane permeability, chromosomally encoded cephalosporinase and an extensive efflux pump system4. Besides, P. aeruginosa holds a prominent place in development of acquired resistance through horizontal transfer of genetic elements (integrons, transposons and plasmids) which contain resistance genes as well as by chromosomal mutations in DNA gyrase and topoisomerase IV5,6.
Plasmid mediated quinolone resistance develops mainly through the production of qnr proteins7. First qnr determinant was introduced as qnrA and the subsequent were known as qnrB, qnrC, qnrS and qnrD8. Furthermore, an enzyme called aac(6')-Ib causes aminoglycoside resistance by acetylation and a variant of this enzyme, aac(6')-Ib-cr was found to be another cause of plasmid-mediated quinolone resistance9. Mutations in DNA gyrase and topoisomerase IV arise most commonly in gyrA and parC subunits, respectively10. Because of the consistent occurrence of the mutations in gyrA region from amino acid position 67-106, it is commonly referred to as Quinolone Resistance Determining Region (QRDR)11.
Chelonians, comprised of turtles, terrapins and tortoises are kept in captivity for commercial purposes, particularly as pets. So far, turtle keeping has been continuously growing but it has not been matched with an appropriate increase in medical knowledge12. Pet turtles have been known to harbor a variety of zoonotic bacteria, especially in their digestive tract13. Among these, Salmonella spp., in particular have been well-studied as a source of salmonellosis in humans14,15. In the meantime, P. aeruginosa has been isolated from sea turtles and desert tortoises16,17. A MLST analysis has been conducted for the P. aeruginosa isolated from pet CSN turtles18. Moreover, P. aeruginosa isolated from pet turtles and turtle eggs exhibited a high-level aminoglycoside resistance19.
Chinese Stripe-Necked (CSN) turtle (Ocadia sinensis) is a medium-sized geoemydid turtle (up to 24 cm carapace length) with numerous dark-bordered, narrow yellow stripes on the head and neck. The CSN turtles are recognized as native inhabitants of Taiwan, Southern China and Northern Vietnam20. But, they are now being distributed worldwide and also imported to South Korea as a pet. The CSN turtle shows an omnivorous dietary habit which is surprisingly tolerant of harsh and even polluted aquatic systems20. Hence, ease of keeping could be the reason CSN to become more popular as a pet.
Current study sought to characterize the P. aeruginosa isolated from pet CSN turtles through the detection of quinolone susceptibility and screening the plasmid-mediated quinolone resistance genes (qnrA, qnrB, qnrS and aac(6')-Ib-cr) and chromosomal QRDR mutations in gyrA and parC for a better understanding of an opportunistic pathogen in pet turtles and potential zoonotic pathogen in human.
MATERIALS AND METHODS
Purchase of pet turtles: A total of 40 Chinese stripe-necked turtles were purchased from several pet shops and online markets in Korea. The turtles were under 4 weeks of age at purchase and studied immediately. All turtles were clinically inspected upon purchase and showed no signs of disease. Turtles were raised under laboratory conditions following the general husbandry method21.
Isolation of P. aeruginosa: Cloacal swabs from turtles were enriched by submerging them in Tryptic Soy Broth (TSB) and incubating at 37°C for 24 h. Enriched samples were streaked onto the selective culture medium cetrimide agar (CN) and incubated at 37°C for 24 h. Suspicious colonies were subcultured onto CN agar and incubated once more at 42°C for 24 h. The isolates that showed the positive growth were presumptively identified as P. aeruginosa.
Bacterial identification using 16S rRNA gene amplification and sequencing: Genomic DNA was extracted from presumptively identified P. aeruginosa isolates by Chelex 100 extraction method and PCR for 16S rRNA was performed using universal primers 12F and 1492R. Amplicons were sequenced and tested for the similarity using BLAST algorithm of NCBI database so as to confirm the species status.
Quinolone susceptibility testing: Twenty out of 31 identified isolates were randomly selected for the study. Susceptibility pattern of the P. aeruginosa isolates was investigated for nalidixic acid, ciprofloxacin, ofloxacin and levofloxacin by disk diffusion test on Mueller Hinton agar (MBcell Ltd., Seoul, Korea). Minimum Inhibitory Concentrations (MIC) of nalidixic acid, ciprofloxacin and ofloxacin were detected by broth microdilution method. All susceptibility testing were conducted according to the recommendations of Performance Standards for Antimicrobial Susceptibility Testing, Clinical and Laboratory Standards Institute22.
Detection of quinolone resistance genes: The presence of qnrA, qnrB and qnrS genes were detected using primers and PCR conditions listed in Table 1. Each PCR mixture was 20 μL total volume comprising of 10 μL of Quick Taq® HS DyeMix (Toyobo, Japan) 1 μL of 10 pmol μL–1 each primer and 1 μL of the template. The PCR products were checked in 1.5% agarose gel.
Detection of aac(6’)-Ib-cr variant: The PCR was performed to amplify the aac(6’)-Ib gene region by using the same protocol used for qnr genes (Table 1). Amplicons were purified using Expin™ PCR SV kit (GeneAll®, Korea) and were sent to Cosmogenetech Co., Ltd., Daejeon, Korea for sequencing. Acquired sequences were tested for the similarity in BLAST option in NCBI database in order to check the presence of aac(6’)-Ib-cr gene variant.
Amplification of Quinolone Resistance Determining Region (QRDR) and detection of mutations: The PCR for gyrA and parC regions were performed using primers and conditions described in Table 1. The PCR was conducted in 50 μL volumes consisting of 20 μL of Quick Taq® HS DyeMix (Toyobo, Japan), 2 μL of 10 pmol μL–1 each primer and 2 μL of the template. Amplicons were purified using Expin™ PCR SV kit (GeneAll®, Korea) and sent to Cosmogenetech Co., Ltd., Daejeon, Korea for sequencing. Acquired sequences were subjected to detection of mutations by comparison with published NCBI reference sequences (Accession No. L29417.1 and AB003428.1 for gyrA and parC, respectively) as described by Rubin et al.23. Analyzing and comparison of QRDR sequences were performed using Mutation Surveyor V5.0.1 (Softgenetics LLC, USA) software.
| Table 1: | Primers used to amplify QRD, qnr genes and conditions of each reaction |
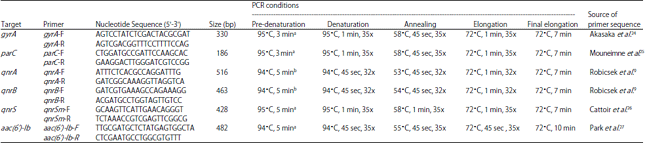 | |
| aPCR conditions were modified for this study, bPCR conditions were adapted from the reference source | |
| Table 2: | Details of the gyrA gene sequences of P. aeruginosa downloaded from NCBI database for the phylogenetic analysis |
 | |
| Table 3: | Antimicrobial susceptibility pattern and genetic characterization of quinolone resistance of Pseudomonas aeruginosa isolated from pet Chinese stripe-necked turtles |
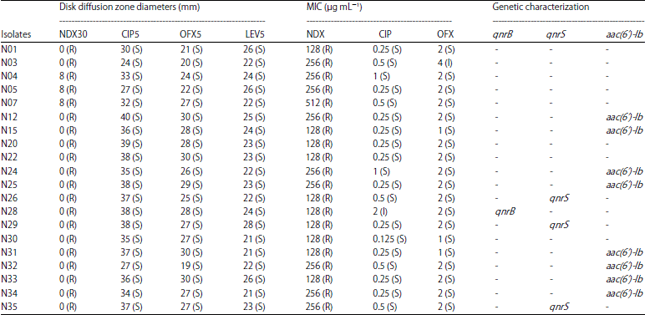 | |
NDX30: Nalidixic acid (30 μg), CIP5: Ciprofloxacin (5 μg), OFX5: Ofloxacin (5 μg), LEV5: Levofloxacin (5 μg), S: Susceptible, I: Intermediate, R: Resistant were designated using breakpoints described by the Clinical Laboratory Standards Institute22 | |
Phylogenetic comparison of gyrA sequences with published sequences of different origin: The sequences derived for gyrA gene region were analyzed and a neighbor-joining phylogenetic tree was obtained with 1000 bootstrap replications. For the analysis, 10 gyrA sequences of P. aeruginosa of different origin (i.e., clinical and environmental) were obtained from published sequences of the GenBank (Table 2 for the accession numbers and details) and MEGA6 sequence analyzing software was used for aligning and construction of the phylogenetic tree.
Submission of DNA sequences to NCBI: A total of nine representative sequences, including five gyrA and four aac(6’)-Ib, acquired from the current study were submitted to GenBank.
RESULTS
Isolation and identification of P. aeruginosa: Thirty-one of forty isolates showed positive growth on CN agar at 42°C. A subsequent BLAST search after 16S rRNA sequencing indicated a 99-100% match to P. aeruginosa sequences available in GenBank which confirmed their identity as P. aeruginosa.
Quinolone susceptibility testing: The results of the quinolone susceptibility testing are shown in Table 3. Consistent with the results of both disc diffusion test and MIC test, all 20 (100%) isolates of P. aeruginosa showed resistance to nalidixic acid. In disc diffusion test, no isolate showed intermediate resistance or resistance to other three quinolones, ciprofloxacin, ofloxacin and levofloxacin.
 | |
| Fig. 1: | Neighbor-joining phylogenetic tree derived by analyzing P. aeruginosa gyrA sequences obtained from the current study and downloaded from NCBI, sequences referred to as EU753434.1, EU753435.1, KP281423.1, KP281423.1, KU522119.1, KU380335.1, JQ844605.1, JQ844604.1, KJ920445.1 and KJ920446.1 were obtained from NCBI public database and the rest of the sequences were acquired from the current study, 1, 2, 3: Major clads |
| Table 4: | Accession numbers of the gyrA and parC sequences which were submitted to NCBI |
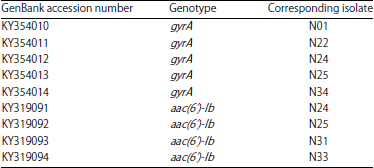 | |
In MIC test, two isolates, N03 and N28 showed intermediate resistance to ciprofloxacin (5%) and ofloxacin (5%) respectively (Table 3).
Detection of quinolone resistance genes (qnrA, qnrB, qnrS and aac(6’)-Ib-cr) and QRDR mutations: Table 3 shows that, none of the isolates possessed qnrA gene and only one isolate referred to as N28 harbored qnrB gene (5%). Three isolates showed qnrS gene (15%), while aac(6’)-Ib could be amplified from 8 isolates (40%) (Table 3). However, sequencing results could not identify any aac(6’)-Ib-cr variant in BLAST comparison. Neither gyrA nor parC sequences exhibited mutations in QRDR, within amino acid positions 67-106.
Phylogenetic comparison of gyrA sequences: Phylogenetic tree derived by analyzing and comparing the gyrA gene sequences obtained by current study and the published NCBI sequences is illustrated in Fig. 1. Neighbor-joining phylogenetic tree indicated three distinct clads comprising, clad 1 current study isolates, clad 2 clinical isolates of human and dogs and clad 3 isolates from soil and water.
Submission of DNA sequences to NCBI: Accession numbers of the gyrA and parC sequences which have been deposited in the GenBank database were listed in Table 4.
DISCUSSION
It is much reasonable to deem that P. aeruginosa is an opportunistic pathogen which is able to cause serious infections in animals and human1,2. In the meantime, it's intrinsic and acquired resistance to antimicrobials has also been well studied4,6–8. Due to higher significance and implications counting current health concerns, studies associated with quinolone resistance, particularly, genetics-based resistance of P. aeruginosa are more pronounced23,28-30.
Yonezawa et al.31 have studied the gyrA mutations of quinolone-resistant P. aeruginosa clinical isolates in the early era. Descending from such studies numerous similar studies have been conducted29,32,33. Parallel clinical studies related to animals were directed to canine infections of dogs and isolates from poultry23, 30,34. Meanwhile, a study focused on pet turtles encountered a range of antimicrobial-resistant bacteria including P. aeruginosa which showed a high-level aminoglycoside resistance, particularly to gentamicin19. However, to our knowledge, quinolone resistance studies regarding P. aeruginosa from pet turtles have not been conducted so far.
Being consistent with above, the current study was conducted to characterize the P. aeruginosa isolated from pet CSN turtles. The CSN turtle is a common species of pet turtles being kept by South Korean people, hence there is a possibility of getting opportunistic infections into humans who handle the turtles and on the other hand, might be a potential pathogen to the turtle itself when they are immunologically compromised.
In accordance with the disk diffusion and MIC outcomes, 100% resistance to nalidixic acid was observed. More or less similar outcome was observed with 96% (102/106 isolates) resistance to nalidixic acid from canine infected dog isolates23. In contrast, Al-Marjani35 reported P. aeruginosa clinical isolates showing 42.1 and 39.4% resistance to levofloxacin and nalidixic acid, respectively where environmental isolates showed the highest resistance rate (27.5%) to nalidixic acid. But, isolates of this study did not show any resistance to tested fluoroquinolones, ciprofloxacin, ofloxacin and levofloxacin. In line with this, a previous study reported P. aeruginosa isolated from a river which showed a 100% susceptibility to ciprofloxacin and levofloxacin36. Besides, multidrug-resistant clinical isolates from pediatric patients could show 94.8% (55/58) resistance to both ciprofloxacin and levofloxacin37. However, MIC results indicated two isolates showing intermediate resistance, isolate N03 to ofloxacin and isolate N28 to ciprofloxacin but only N28 could show qnrB gene. In the meantime, Michalska et al.28 have detected qnrB as the most prevalent in P. aeruginosa but Cayci et al.38 could not detect any of the qnr genes even in resistant strains. However, another study reported the presence of both qnrS and qnrA simultaneously in clinical samples (21% qnrS and 13.1% qnrA), while 2.5% (1/40) qnrS could be observed in sewage and tap water isolates35. In this study, three isolates which harbored qnrS were susceptible to all tested quinolones, except nalidixic acid. Even though it is controversial, a similar kind of observation was reported, in which Aeromonas spp., isolated from freshwater fishes harbored qnr genes and QRDR mutations, despite they are susceptible to quinolones39. Therefore, the necessity of investigating the organismal factors and mechanisms which could alter the gene expression owns significance. In the case of aac(6’)-Ib gene, none of the isolates could show the aac(6’)-Ib-cr variant, denoting that quinolone resistance is not mediated through aac(6’)-Ib-cr gene. Therefore, it could be a sign of aminoglycoside resistance as Diaz et al.19 and Michalska et al.28 reported but not plausible without further experiments.
A few isolates showed qnr genes but no QRDR mutations in both gyrA and parC regions were detected in any of the isolates. Although, every isolate showed resistance to nalidixic acid, a substantial resistance to fluoroquinolones was not detected. Because, plasmid-mediated qnr genes are not strong enough to show a phenotypic resistance unless it is facilitated by QRDR chromosomal mutations7. In Gram-negative bacteria, DNA gyrase seems to be the primary cellular target of quinolones, particularly fluoroquinolones. With regards to P. aeruginosa gyrA, the most common mutations that lead to drug resistance occur at Thr83 and Asp87 of the subunit11. In a human clinical study, 50% (19/38) of the ciprofloxacin-resistant strains harbored gyrA Thr83Ile mutation and parC Ser80Leu mutation40. Some other animal and human clinical studies also reported a range of QRDR mutations, majorly in gyrA and parC in which the isolates were showing a higher resistance to quinolones23,29. Although, some non-specific silent mutations of the amplified region were noticed, current study could not identify any of above mentioned specific point mutations within the amino acid positions 67-106. Therefore, it can be stated that QRDRs of every isolate in this study isolates is more conserved compared to highly resistant isolates.
According to the personal communication with pet shops, the studied group of turtles was very young and aged less than 4 weeks. All the individuals were purchased soon after hatched did not show any sign of disease and were confirmed not exposed to any antibiotics. It could be the reason P. aeruginosa isolates to become more susceptible to quinolone antibiotics. Nevertheless, a few isolates show qnr genes in their genome. Since those mediators are transferable from other organisms, two possible causes can be addressed. One is those plasmids might have horizontally transferred from other bacterial species while the newly hatched turtles were kept together with other turtles in the same cage till purchase. And the second is those strains could get plasmids from other bacterial species which thrive in the turtle's own body.
Phylogenetic analysis of gyrA sequences produced a neighbor-joining tree showing 3 major clads referred to as 1, 2 and 3. Interestingly, clad 1 comprised of isolates of the current study, clad 2 contained all clinical isolated both from human and pet dogs. The isolates from water and soil were separated as last clad, 3. This outcome clearly interprets the genetic similarity of P. aeruginosa isolated from CSN turtles and how they diverge from clinical and environmental isolates. As the phylogenetic tree portrays, the environmental isolates represent the basal clad and all other isolates separated arising from it. It might be caused by the source of bacteria separating animal isolates from environmental. A comparative genomic study also reported separated clusters in dendrogram revealing the genetic difference among human, bovine and environmental isolates of P. aeruginosa41. Although clinical and current study isolates are more or less closely related than with environmental isolates, still they keep in two distinct clads revealing that isolates from these young CSN turtles are genetically more unique and conserved than quinolone-resistant clinical isolates. Bootstrap node support for the separation of clinical isolates and current study isolates were 59 and 36% respectively. Furthermore, it is important to observe the 99% bootstrap separation of isolates from soil samples and water in clad 3, revealing the genetic diversity of P. aeruginosa even between water and land.
CONCLUSION
It is plausible that the P. aeruginosa isolated from CSN turtles are not resistant to tested fluoroquinolones and their QRDRs were genetically more conserved than that of quinolone-resistant strains. So it can be a plus point for the people who keep pet CSN turtles but further studies regarding more turtle species and more antimicrobials are highly recommended for a better understanding.
ACKNOWLEDGMENT
This study was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2015R1D1A1A01060638).
REFERENCES
- Porras-Gomez, M., J. Vega-Baudrit and S. Nunez-Corrales, 2012. Overview of multidrug-resistant Pseudomonas aeruginosa and novel therapeutic approaches. J. Biomater. Nanobiotechnol., 3: 519-527.
CrossRefDirect Link - Ciocan, O.A., M. Carp-Carare, C. Rimbu, A.P. Cozma, C. Carp-Carare, E. Guguianu and C.V. Panzaru, 2015. The incidence of dog recurrent otitis caused by strains of Multidrug-Resistant (MDR) Pseudomonas aeruginosa. Lucrari Stiintifice-Medicina Veterinara, 48: 26-31.
Direct Link - Lister, P.D., D.J. Wolter and N.D. Hanson, 2009. Antibacterial-resistant Pseudomonas aeruginosa: Clinical impact and complex regulation of chromosomally encoded resistance mechanisms. Clin. Microbiol. Rev., 22: 582-610.
CrossRefDirect Link - Linder, J.A., E.S. Huang, M.A. Steinman, R. Gonzales and R.S. Stafford, 2005. Fluoroquinolone prescribing in the United States: 1995 to 2002. Am. J. Med., 118: 259-268.
CrossRefDirect Link - Livermore, D.M., 2002. Multiple mechanisms of antimicrobial resistance in Pseudomonas aeruginosa: Our worst nightmare? Clin. Infect. Dis., 34: 634-640.
CrossRefDirect Link - Martinez-Martinez, L., A. Pascual and G.A. Jacoby, 1998. Quinolone resistance from a transferable plasmid. Lancet, 351: 797-799.
CrossRefDirect Link - Strahilevitz, J., G.A. Jacoby, D.C. Hooper and A. Robicsek, 2009. Plasmid-mediated quinolone resistance: A multifaceted threat. Clin. Microbiol. Rev., 22: 664-689.
CrossRefDirect Link - Robicsek, A., J. Strahilevitz, G.A. Jacoby, M. Macielag and D. Abbanat et al., 2006. Fluoroquinolone-modifying enzyme: A new adaptation of a common aminoglycoside acetyltransferase. Nat. Med., 12: 83-88.
CrossRefDirect Link - Di Ianni, F., P.L. Dodi, C.S. Cabassi, I. Pelizzone and A. Sala et al., 2015. Conjunctival flora of clinically normal and diseased turtles and tortoises. BMC Vet. Res., Vol. 11.
CrossRef - Warwick, C., P.C. Arena, C. Steedman and M. Jessop, 2012. A review of captive exotic animal-linked zoonoses. J. Environ. Health Res., 12: 9-24.
Direct Link - Bosch, S., R.V. Tauxe and C.B. Behravesh, 2015. Turtle-associated salmonellosis, United States, 2006-2014. Emerg. Infect. Dis., 22: 1149-1155.
CrossRefDirect Link - Dickinson, V.M., T. Duck, C.R. Schwalbe, J.L. Jarchow and M.H. Trueblood, 2001. Nasal and cloacal bacteria in free-ranging desert tortoises from the Western United States. J. Wildlife Dis., 37: 252-257.
CrossRefDirect Link - Warwick, C., P.C. Arena and C. Steedman, 2013. Health implications associated with exposure to farmed and wild sea turtles. JRSM Short Rep., Vol. 4.
CrossRefDirect Link - Wendt, M. and G.J. Heo, 2016. Multilocus sequence typing analysis of Pseudomonas aeruginosa isolated from pet Chinese stripe-necked turtles (Ocadia sinensis). Lab. Anim. Res., 32: 208-216.
CrossRefDirect Link - Diaz, M.A., R.K. Cooper, A. Cloeckaert and R.J. Siebeling, 2006. Plasmid-mediated high-level gentamicin resistance among enteric bacteria isolated from pet turtles in Louisiana. Applied Environ. Microbiol., 72: 306-312.
CrossRefDirect Link - Rubin, J., R.D. Walker, K. Blickenstaff, S. Bodeis-Jones and S. Zhao, 2008. Antimicrobial resistance and genetic characterization of fluoroquinolone resistance of Pseudomonas aeruginosa isolated from canine infections. Vet. Microbiol., 131: 164-172.
CrossRefDirect Link - Akasaka, T., M. Tanaka, A. Yamaguchi and K. Sato, 2001. Type II topoisomerase mutations in fluoroquinolone-resistant clinical strains of Pseudomonas aeruginosa isolated in 1998 and 1999: Role of target enzyme in mechanism of fluoroquinolone resistance. Antimicrob. Agents Chemother., 45: 2263-2268.
CrossRefDirect Link - Mouneimne, H., J. Robert, V. Jarlier and E. Cambau, 1999. Type II topoisomerase mutations in ciprofloxacin-resistant strains of Pseudomonas aeruginosa. Antimicrob. Agents Chemother., 43: 62-66.
Direct Link - Cattoir, V., L. Poirel, V. Rotimi, C.J. Soussy and P. Nordmann, 2007. Multiplex PCR for detection of plasmid-mediated quinolone resistance qnr genes in ESBL-producing enterobacterial isolates. J. Antimicrob. Chemother., 60: 394-397.
CrossRefDirect Link - Park, C.H., A. Robicsek, G.A. Jacoby, D. Sahm and D.C. Hooper, 2006. Prevalence in the United States of aac(6′)-Ib-cr encoding a ciprofloxacin-modifying enzyme. Antimicrob Agents Chemother., 50: 3953-3955.
CrossRefDirect Link - Michalska, A.D., P.T. Sacha, D. Ojdana, A. Wieczorek and E. Tryniszewska, 2014. Prevalence of resistance to aminoglycosides and fluoroquinolones among Pseudomonas aeruginosa strains in a University Hospital in Northeastern Poland. Braz. J. Microbiol., 45: 1455-1458.
CrossRefDirect Link - Wang, Y.T., M.F. Lee and C.F. Peng, 2014. Mutations in the quinolone resistance-determining regions associated with ciprofloxacin resistance in Pseudomonas aeruginosa isolates from Southern Taiwan. Biomarkers Genomic Med., 6: 79-83.
CrossRefDirect Link - Lin, D., S.L. Foley, Y. Qi, J. Han and C. Ji et al., 2012. Characterization of antimicrobial resistance of Pseudomonas aeruginosa isolated from canine infections. J. Applied Microbiol., 113: 16-23.
CrossRefDirect Link - Yonezawa, M., M. Takahata, N. Matsubara, Y. Watanabe and H. Narita, 1995. DNA gyrase gyrA mutations in quinolone-resistant clinical isolates of Pseudomonas aeruginosa. Antimicrob. Agents Chemother., 39: 1970-1972.
CrossRefDirect Link - Loureiro, M.M., B.A. de Moraes, V.L.F. Mendonca, M.R.R. Quadra, G.S. Pinheiro and M.D. Asensi, 2002. Pseudomonas aeruginosa: Study of antibiotic resistance and molecular typing in hospital infection cases in a neonatal intensive care unit from Rio de Janeiro City, Brazil. Memorias Instituto Oswaldo Cruz, 97: 387-394.
CrossRefPubMedDirect Link - Nakano, M., T. Deguchi, T. Kawamura, M. Yasuda, M. Kimura, Y. Okano and Y. Kawada, 1997. Mutations in the gyrA and parC genes in fluoroquinolone-resistant clinical isolates of Pseudomonas aeruginosa. Antimicrob. Agents Chemother., 41: 2289-2291.
Direct Link - Ogunleye, A.O., 2012. Identification of GyrA mutations conferring fluoroquinolone resistance in Pseudomonas aeruginosa isolated from poultry in Ibadan, Oyo State, Nigeria. Afr. J. Microbiol. Res., 6: 1573-1578.
CrossRefDirect Link - Al-Marjani, M.F., 2014. Presence of qnr gene in environmental and clinical Pseudomonas aeruginosa isolates in Baghdad. Int. J. Curr. Microbiol. Applied Sci., 3: 853-857.
Direct Link - Kittinger, C., M. Lipp, R. Baumert, B. Folli and G. Koraimann et al., 2016. Antibiotic resistance patterns of Pseudomonas spp. isolated from the river Danube. Front. Microbiol., Vol. 7.
CrossRefDirect Link - Ochoa, S.A., A. Cruz-Cordovaa, G.E. Rodea, V. Cazares-Dominguez and G. Escalona et al., 2015. Phenotypic characterization of multidrug-resistant Pseudomonas aeruginosa strains isolated from pediatric patients associated to biofilm formation. Microbiol. Res., 172: 68-78.
CrossRefDirect Link - Cayci, Y.T., A.Y. Coban and M. Gunaydin, 2014. Investigation of plasmid-mediated quinolone resistance in Pseudomonas aeruginosa clinical isolates. Indian J. Med. Microbiol., 32: 285-289.
CrossRefDirect Link - Chenia, H.Y., 2016. Prevalence and characterization of plasmid-mediated quinolone resistance genes in Aeromonas spp. isolated from South African freshwater fish. Int. J. Food Microbiol., 231: 26-32.
CrossRefDirect Link - Salma, R.M., F.A. Dabboussi, I.O. Kassaa, R.H. Khudary and M.M. Hamze, 2012. gyrA and parC mutations in quinolone-resistant clinical isolates of Pseudomonas aeruginosa from Nini Hospital in North Lebanon. Int. J. Microbiol. Res., 3: 82-86.
Direct Link - Szmolka, A., N. Cramer and B. Nagy, 2012. Comparative genomic analysis of bovine, environmental and human strains of Pseudomonas aeruginosa. FEMS Microbiol. Lett., 335: 113-122.
CrossRefDirect Link