
Lakshman Santra
Division of Veterinary Biotechnology, Indian Veterinary Research Institute, Izatnagar, 243122 Bareilly, Uttar Pradesh, India
Saurabh Gupta
Division of Veterinary Biotechnology, Indian Veterinary Research Institute, Izatnagar, 243122 Bareilly, Uttar Pradesh, India
LiveDNA: 91.10672
Ajay Kumar Singh
Division of Veterinary Biotechnology, Indian Veterinary Research Institute, Izatnagar, 243122 Bareilly, Uttar Pradesh, India
Manish Mahawar
Division of Animal Biochemistry, Indian Veterinary Research Institute, Izatnagar, 243122 Bareilly, Uttar Pradesh, India
Ravikumar Gandham
Division of Veterinary Biotechnology, Indian Veterinary Research Institute, Izatnagar, 243122 Bareilly, Uttar Pradesh, India
Sujoy K. Dhara
Division of Veterinary Biotechnology, Indian Veterinary Research Institute, Izatnagar, 243122 Bareilly, Uttar Pradesh, India
LiveDNA: 91.1986
Asian Journal of Animal and Veterinary Advances
Year: 2017 | Volume: 12 | Issue: 3 | Page No.: 123-131







ABSTRACT
The RNA interference is an established phenomenon of eukaryotic gene silencing. Recent advancement of genetic engineering allows employ this powerful tool for silencing multiple genes in several domestic animals and thus adds an additional dimension to animal welfare and livestock product improvement. This study reviews the current state of and prospects for the development of transgenic livestock using the mighty tool RNAi. The mini review accesses and summarizes information available in listed references. It starts with brief introduction to basis of RNA interference followed by outlining the current application of RNAi mediated transgenesis in livestock sector and identifies potential ways of developing and improving genetically modified livestock produces. This mini review explains importance of RNAi as a tool for improving animal produce. Here we present highlights of different recent studies that describe gene silencing by RNAi as an intervention tool for improvement of quality of various animal produces, acceleration of postnatal body growth, protection of livestock animals from certain fatal diseases etc. Diverse approaches are described to identify potential ways of developing genetically modified livestock. Further, this study emphasizes on recent advances in the field of gene targeting mentioning briefly on diverse molecular scissors and their potential involvement in executing conditional knock out or shRNA mediated knockdown strategies. Overall this mini review provides an update of the current status of harnessing RNAi for intended livestock produce and also points to the trend of future transgenic animal husbandry towards making the concept of ‘an animal for all purpose’ to come into reality.
PDF Abstract XML References Citation
How to cite this article
Lakshman Santra, Saurabh Gupta, Ajay Kumar Singh, Manish Mahawar, Ravikumar Gandham and Sujoy K. Dhara, 2017. RNAi Mediated Transgenesis for Improving Animal Produce. Asian Journal of Animal and Veterinary Advances, 12: 123-131.
DOI: 10.3923/ajava.2017.123.131
URL: https://scialert.net/abstract/?doi=ajava.2017.123.131
DOI: 10.3923/ajava.2017.123.131
URL: https://scialert.net/abstract/?doi=ajava.2017.123.131
INTRODUCTION
The RNA interference (RNAi) is a natural defence mechanism displayed by most of eukaryotes to protect themselves from exogenous RNA molecules. A process to mimic RNAi has been developed and this has gained popularity as a method of choice for silencing gene expression in a variety of systems. This method uses double stranded RNAs (dsRNAs) to target complementary RNAs for destruction. Hence, custom designed dsRNAs, complementary to a specific gene, help targeted mRNA degradation and thus results in gene silencing or gene knockdown. Loss of function phenotypes of many genes can be engendered reliably and quickly by RNAi via introduction of chemically synthesized small interfering RNA (siRNA) oligonucleotides or vectors that express siRNAs using cellular machineries. Recent advancement of genetic engineering allows employ this powerful tool for silencing many genes in a wide range of animals. This helps confer resistance to livestock against several fatal diseases and also make animal produce more suitable (healthy and safe) for human use. Thus RNAi based approach adds an additional dimension to animal welfare and product improvement. Here in this short review, about the basics of RNAi and potential applications of it was discussed for improving animal produce.
BASICS OF RNAi
Phenomenon of RNAi was first elaborated by Fire et al.1 as a mechanism that induces post-transcriptional, sequence-specific RNA degradation in response to introduction of long dsRNAs in Caenorhabditis elegans. Later it was realised that this silencing mechanism is universal and is found in diverse set of organisms. In plant system RNAi is known as co-suppression and in fungi the same phenomenon is called quelling2.
Several genetic and biochemical studies on flies, worms, fungi, plants and mammalian system have facilitated understand the mechanism of RNAi based gene silencing. The core RNA-silencing response involves processing of the trigger dsRNA into smaller 21-25 bp fragments with dinucleotide 3’ overhangs by an ATP dependent enzyme dicer 3. Also, 21-25 bp double stranded fragments produced by dicer activity are referred to as small interfering RNA (siRNA). These siRNAs serve as ‘guide’ to bring nuclease machinery to target mRNA. Each siRNA associates with a protein complex called RNA inducing silencing complex (RISC)4. Both dicer and RISC are evolutionarily most conserved components of RNAi silencing machinery. The RISC complex allows base pairing between antisense strand and target mRNA. Such base pairing of antisense strand leads to endonucleolytic cleavage of the target mRNA. As a result an mRNA is cleaved into two fragments: One missing its poly A tail and other fragment missing the 5’ cap. Both these conditions make mRNA fragments vulnerable to further degradation by RNA surveillance machinery5 (Fig. 1).
Following one round cleavage the siRNA/RISC complex becomes free to target another mRNA molecule. Thus, the initial trigger dsRNA generates several siRNA, each of these siRNAs recruits and activates a RISC complex which targets multiple mRNAs. This indicates about the sub-stoichiometric potency of the trigger dsRNA. Recent evidences suggest that in some species, additional amplification occurs via a mechanism termed ‘transitive RNAi’6. In this mechanism, any siRNA or even short antisense RNA functions to prime specific RNA-dependent RNA polymerases (RdRPs) to synthesize copy RNA (cRNA) of the target RNA. Such de novo produced dsRNAs serve as substrates to dicer and are cleaved into additional siRNAs6.
FUNCTIONS
The first and foremost function of RNA interference is protection of cells from various viral RNAs and transposable elements7. Other function includes the miRNAs (microRNA) associated regulation of development, especially at the timing of morphogenesis and the maintenance of undifferentiated state of stem cells8. The miRNAs mediated RNAi is also linked to the formation of tumors and dysregulation of the cell cycle in many organisms, including human being. Here, miRNAs function as both oncogenes and tumor suppressors.
Apart from post-transcriptional gene silencing mechanism, RNAi is also reported to directly modify DNA at genome level. Many experimental evidences establish that siRNA can inactivate transcription through direct DNA methylation and other type of covalent modification in the genomes of certain species. In case of S. pombe, RNAi machinery plays a critical role in formation and maintenance of higher order chromatin structure and function9. It is further reported that siRNAs have direct role in eliminating specific DNA sequences during programmed genome arrangement in the protest isolate Tetrahymena10.
POTENTIALS
The RNAi is a powerful tool for inhibiting gene function in a s equence-specific manner. Its immense potentials is now being used for generating loss-of-function or reduction-of-function phenotypes for studying the functional genomics11.
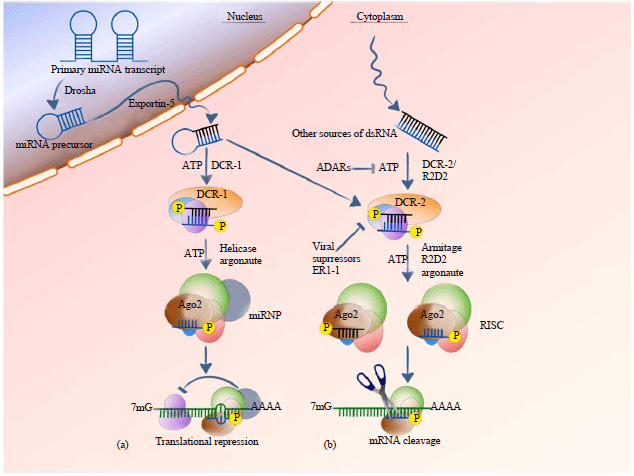 | |
| Fig. 1(a-b): | Schematic representation of small RNA guided post-transcriptional gene silencing, (a) Primary miRNA transcripts are processed to miRNA precursors in the nucleus by the RNase-III-like enzyme drosha. The miRNA precursor is subsequently exported to the cytoplasm through nuclear receptor exportin-5. Dicer additionally processes this miRNA precursor to siRNA-duplex-like intermediates that subsequently is unwound while assembling into miRNP/RISC. Mature miRNAs then bind to Ago proteins that cause translational repression or cleavage of target mRNAs and (b) There may be other sources of long dsRNA in the cytoplasm e.g., viral RNAs, artificially introduced dsRNA, dsRNAs generated by RdRPs and genomic sense and antisense transcripts. Similar to miRNA precursors, long dsRNA is processed by the RNase III enzyme dicer into 21-23 nucleotide dsRNA intermediates. With the help of RNA helicase, armitage and R2D2, the single-stranded siRNA-containing RISC is formed |
This has immense implication in biotechnology and in biomedical research.
Gene targeting by short hairpin RNA (shRNA) construct: The RNAi can be adopted for mammalian cells by different methods. The most common is transient transfection of siRNA oligonucleotides. The other alternative approach is use of in vivo expression constructs for generation of small dsRNA triggers. This construct consists mainly of two complementary 19-23 bp RNA sequences linked by a short loop of 4-11 nucleotides, similar to the hairpin found in miRNA12. Following entry into the nucleus the construct is expressed under suitable promoter such as RNA polymerase III. The resulting transcript is subsequently exported to the cytoplasm, processed by dicer, then get incorporated into RISC and finally shunted into the RNAi pathway.
From the practical point of view, use of shRNA (short hairpin RNA) has certain advantage over siRNA. Once made, use of shRNA is inexpensive compared to considerable cost involved in chemically synthesized siRNAs. Since shRNAs resemble endogenously expressed hairpin RNAs, these provide stable expression of silencing trigger. Therefore, one does not require repeated siRNAs transfection13. Moreover, shRNA constructs can be delivered using viral vectors into primary cell types and even in whole organism. On the other hand, transfection of siRNA may be difficult in many primary cells.
Constitutive knockdown of target gene: To avoid repeated shRNA cassette delivery, it is desirable to have a system/method for long-term stable siRNA expression at organ levels. So, efforts are being directed towards generation of transgene integrated genome with gene targeting technologies that would support constant siRNAs supply14. Once stably integrated in germ line, the construct would pass to next generations by inheritance15. This approach would cut down the cost and reduce the recombinant viral vector stimulated immunological complications also.
Stage specific/conditional knockdown of target gene: It is sometimes observed that inhibition of a gene that plays a critical role during embryonic development often leads to embryonic lethality. For such a case, constant gene knockdown is not a good choice. Therefore to overcome embryonic lethality and to silence gene functions in specific tissues or in a specific window of time during development, vectors for conditional knockdown must be used. This allows gene silencing events to happen at specific cell types or at specific developmental stages16,17.
Conditionality can be achieved by using either Cre/loxP system18 or Flp/frt system19. Cre/loxP system is based on the site-specific Cre recombinase (Cyclization recombination) of the bacteriophage P1, whereas, Flp/frt system is based on Flp recombinase (flipase) of yeast. Cre and Flp recombinases recognize 34 bp sequences, called loxP (locus of crossing [x-ing]-over of bacteriophage P1) and frt (flippase recognition target), respectively. If two recombinase recognition sequences are placed in the same DNA strand and in the same orientation, the site-specific recombination event will result in deletion of the flanked sequence. Several Cre/lox based systems for conversion of constitutively active shRNA expression vectors into conditional shRNA expression vectors are now available20. In these conditional vectors shRNA expression can be regulated by an inserted loxP-flanked ‘stop element’ that blocks shRNA production. After cre-mediated recombination the stop element is removed and the shRNA is expressed. Furthermore, introducing the Cre element under the regulation of inducible promoter additionally employs temporal control over the gene expression. In addition, use of tissue specific promoter(s) to drive the expression of shRNA can restrict tissue specific gene expression21. Thus, inducible conditional strategy has given liberal opportunity to customize RNAi mediated inhibition of a target gene at a given tissue at any time22.
RNAi FOR IMPROVING QUALITY OF ANIMAL PRODUCE
Rapid growth rate: The High Growth (HG) is a phenotype in mice, characterized by post weaning overgrowth without increasing adiposity. This phenotype is expressed due to a loss of function mutation of a candidate gene. The gene is suppressor of cytokine signalling 2 (SOCS2) gene23. The SOCS family of proteins act in a classical negative feedback loop to regulate JAK-STAT signal transduction by a variety of cytokines. Functionally, SOCS proteins interact with cytokine receptors and/or Jak kinases, thereby inhibiting activation of kinases and STAT proteins. The SOCS2 protein is closely associated to growth in mammals through the growth hormone (GH)/insulin-like growth factor I (IGF1) axis. This important physiological GH/IGF1 pathway has central role in longevity determination and aging from yeast to humans. The SOCS2 protein interacts with IGF1 and GH receptors as a negative regulator of growth hormone signalling. Thus, SOCS2 mutation deregulates the GH/IGF1 system. Hence, RNAi mediated suppression or inhibition of the SOCS2 gene would increase post natal body growth and helps attain puberty rapidly.
Another candidate gene that may be targeted for RNAi mediated silencing24 is SOCS1. The SOCS1 has a biological role in the developing mammary gland. This protein acts as a negative regulator of prolactin signalling during expansion and differentiation of the mammary gland cells. It is established that SOCS1 deficiency results in accelerated mammary gland development and rescues lactation in prolactin receptor-deficient condition24. Therefore, it is likely that siRNA based tissue specific gene silencing of SOCS1 during pregnancy would expedite the growth and development of mammary gland. This might also improve the overall milk production.
Increasing meat quantity: Myostatin (MSTN) is a muscle-specific member of the transforming growth factor beta (TGF-β) superfamily. Earlier this gene has been known as growth and differentiation factor-8 (GDF-8). The MSTN is known to have major role in regulation of skeletal muscle growth. It is expressed predominantly in skeletal muscle and acts as a negative regulator of skeletal muscle growth by suppressing proliferation and differentiation of myoblasts. Loss of function mutations in the MSTN gene are associated with increased skeletal muscle mass popularly termed as ‘double muscling’25. This kind of loss of function mutations are naturally found to occur in certain cattle breeds like Belgian Blue, Piedmontese, South Devon and Asturiana de los Valles26. In order to improve growth performance of animal or livestock, strategies have been developed to manipulate MSTN expression. Among these, dsRNA has been applied to induce sequence specific post-transcription gene silencing by employing either siRNA or shRNA, MSTN gene knockdown using RNAi has potential benefits for livestock production, as noted in the case of generation of transgenic sheep27. This would increase live weight, accelerate weight gain, increase carcass weight and improve feeding efficiency. Moreover, MSTN knock down strategies can also be employed in chicken with the hope that this might help produce transgenic chickens with increased muscle mass28.
Improving milk quality: Appropriate changes in ratios of milk proteins or introduction of novel proteins into bovine milk can lead to improvement in existing products. The RNAi approach has been adapted to increase concentration of valuable components in milk (e.g., casein) as well as removing undesirable components (e.g., lactose) in cow milk.
Unlike human milk, cow milk contains the major heat-labile whey protein called, β-lactoglobulin. The exact biological function of it in bovine milk physiology is not known. However, β-lactoglobulin is known to have allergenic properties29. Many people suffer from milk allergy when they take cow milk. It is generally assumed that milk depleted of this β-lactoglobulin protein would be a better source for human consumption. This can be achieved either by disrupting the β-lactoglobulin gene using Homologous Recombination (HR) or by specific knockdown of its expression using RNAi.
However, production of gene knockouts via HR is practically difficult in farm animals due to low recombination efficiencies in primary cells. Consequently, RNAi technology might provide a more straightforward approach to unravel the role of β-lactoglobulin and to reduce the allergenicity of cow’s milk. In principle, the technology for a gene knockdown by RNAi in ruminants is now available and using the same technology cattle with depleted β-lactoglobulin proteins from milk is under development. Reports demonstrated that β-lactoglobulin down regulation concurrently increases the κ-casein and β-casein in milk significantly in transgenic cow with targeted microRNA against β-lactoglobulin29. The κ-casein is a key milk protein and has significant influence on the cheese making process30. Shorter renneting time and the curd firmness are two attributes of quality cheese. Presence of high κ-casein enhanced heat stability of milk and may expedite the renneting process. Additionally, higher β-casein level in milk helps increase the curd firmness. Thus, the β-lactoglobulin depleted milk not only has potentially hypoallergenic properties, its high total casein content should provide for increased calcium levels and high cheese yields too.
Another important gene that deserves RNAi mediated intervention is α-lactalbumin31-33. It is established that targeted knockdown of α-lactalbumin results in the disruption of lactose synthesis. Lactose is the major sugar present in milk. It is synthesized by the lactose synthase complex composed of a galactosyltransferase and α-lactalbumin. The majority of the adult population suffers from intestinal disorders as a consequence of lactose indigestion. Such lactose intolerance results from the physiological down-regulation of the intestinal lactose-hydrolysing enzyme at weaning. Therefore, supply of low lactose milk remains a possible solution and it demands an intervention for in vivo low-lactose milk production. Knockdown of α-lactalbumin expression through RNAi could offer better control to achieve an acceptable reduction of the lactose and water content of milk without impacting on its vital attributes34. Thus it might offer an opportunity to reduce the lactose content and at the same time significantly lower transportation costs of liquid milk.
Animal as bioreactor for production of humanized organs: Another potential field of RNA interference is regenerative medicine. In this field of biomedical application, malfunctioned organs are replaced with new freshly harvested functional one. There are increasing number of people dying every year due to lack of replacement organs, particularly in case of heart, liver and kidney. Transgenic pigs have the potential to alleviate this short-fall since pig remains an animal of choice as bioreactor for harvesting xenogenic organs. Currently it is found that use of pig derived human tissues is hampered by an undesired porcine protein which is evolutionarily lost in primates. This unwanted protein is α(1,3) galactosyltransferase35. This is an enzyme responsible for porcine specific protein glycosylation and it is considered as xenoepitope to primate system. This principal cell-surface xenoepitope is a primary target of a natural antibody and hence is a key determinant in Hyper Acute Rejection (HAR). To avoid these glycosylation related problems, efforts are being made to create transgenic pigs that would enable tissue specific silencing of α(1,3)galactosyltransferase. Animals lacking α(1,3)galactosyltransferase is not expected to synthesize the epitope and should have a reduced HAR response36. This would lead to production of organs suitable for transplantation in human and other primates37 (Fig. 2).
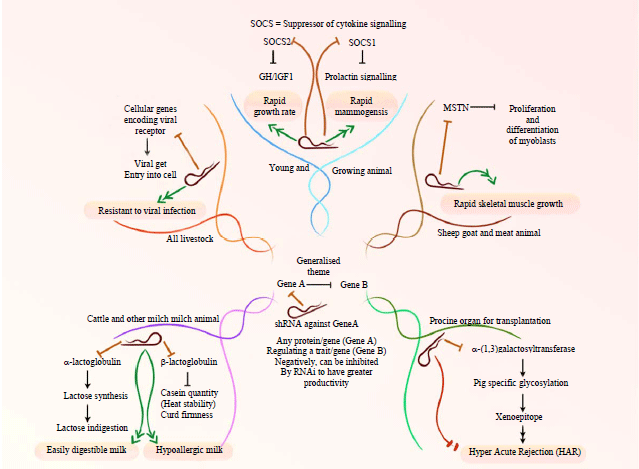 | |
| Fig. 2: | Schematic representation of potential scope of shRNA transgenesis to improve quality of animal produces |
PREVENTING PRION DISEASE
In the recent past, prion has been considered a great threat to international animal and animal product business for its potential zoonotic risk. Prion is an infectious protein that causes fatal neurodegenerative prion diseases or transmissible spongiform encephalopathies. Most popular examples of prion diseases include scrapie in sheep, the mad cow disease in cattle and Creutzfeldt-Jakob disease in human. In these diseases, a misfolded isoform of the cellular prion protein (PrP) accumulates and acts as a novel infectious agent. Since the etiology of prion diseases is not exogenous, the only alternative way to protect animal health is targeting the endogenous vicious genes for inactivation. It has been possible to knock out the ovine prion locus38, however, the cloned lambs carrying the knockout locus died shortly after birth indicating for its inevitability for normal functioning. In this regard RNAi mediated knockdown of PrP expression would be more promising to produce livestock animals resistant to prion diseases39. On the other hand, cloned cattle with a knockout for the prion locus have been successfully produced and indeed show clear evidence of resistance to bovine spongiform encephalopathies infection40. So, gene targeting strategies may vary depending on target species. Resistant livestock, thus free from such diseases would eliminate the risk for potential transmission of the disease to humans and provide additional safeguards for biomedical and food applications.
IMPROVING GENERAL HEALTH
To improve general health and productive efficiency livestock should be prevented from infectious diseases (viral, bacterial etc.). Though conventional breeding programs aim to improve disease resistance but the approach has not been very successful. However, the prospect of transgenic technology is particularly appealing because it provides new and targeted strategies for the improvement of animal health. Gene knockdown strategy might be useful towards suppressing infectious pathogens, particularly viruses by targeting the RNA of the invading agents. The RNA viruses are possibly best suited to this approach, as theoretically both the genomic and the transcribed strands can be targeted41,42. Accordingly it would be possible to interfere simultaneously with replication and expression. Contextually, over two thirds of the OIE list-A pathogens are RNA viruses. These list-A pathogens include foot and mouth disease, classical swine fever and fowl plague, all of which have caused significant outbreaks of disease. An alternative strategy would be to target host genes. In pigs, aminopeptidase N is the primary receptor for the transmissible gastroenteritis43 coronavirus79. Knocking down expression of the gene that encodes this receptor could reduce viral infectivity and enhance resistance, although this strategy might suffer from unpredictable pleiotropic effects.
It is anticipated that the coming future would develop more advanced yet regulated RNA interference based transgenic technology that would result in improved animal welfare, reduced dependence on animal medications, improved production and reproductive performance and ultimately could provide safer food products of superior quality44,45.
In the recent past, majority of transgenesis has been carried out using gene targeting by homologous recombination that occurs with very low frequency. Rather than relying on rare spontaneous events, the frequency of homologous recombination can be considerably increased by introducing a double-strand DNA break at the target site. Highly specific endonuclease enzymes have now been generated to achieve this. The latest of them with ultimate precision are custom designed homing endonuclease46, ZFN47, TALENS48 etc. These technologies allow customisation of target sites for transgene cassette incorporation. To this list a recent addition that has gained huge popularity is CRISPR/Cas system49. These "molecular scissors" now made it possible to introduce a double-strand break at a single predetermined site in the genome. This stimulates the frequency of gene targeting by several orders of magnitude and helps minimizing the chance of off target cleavage during gene targeting.
CONCLUSION AND FUTURE RECOMMENDATION
In the present review we have attempted to describe basics and applications of RNAi for improvement of quality of animal produces. We have emphasized and described trends in the field with published data. From this it is evident that production related traits in livestock are mostly associated with gain of function genes rather than loss of function. So majority of efforts are devoted to ensure the gain of function through transgene integration into the livestock genome. Though scope of gene silencing is relatively less, yet its potential is tremendous.
Delivery of siRNA or shRNA construct remains a bottleneck for a wide application of this technology to generate transgenic animals. Therefore, attentions are shifted from transient siRNA delivery to stable siRNA (shRNA) cassette integration for constitutive gene silencing. This would obviate the need of frequent transfection. Though retro/lenti-viral based transduction system has been found to be efficient but gene construct is integrated randomly in the target cells. This often results in inadvertent off-target effects and such delivery system always remains beyond the control of the researcher. Additionally, specifically for RNAi based technology, it is always better to have multiple regulatory switches controlling the interfering RNA expression. Multilevel control system adds more stringency to the regulatory aspect and thus helps switch on and switch off a particular gene or set of genes in order to have desired trait or product.
Sometimes a gene may have important function for a particular organ or tissue type whereas on other tissue it may not have any identified role. So inactivation or silencing of it may cause adverse effect on health. So tissue specific gene silencing strategies become attractive. These would enable to silence the target gene at a particular tissue only, leaving the rest of the body unaffected. Again, some genes play critical role at embryonic development, so stable inactivation or silencing may cause embryonic lethality. To avoid such consequences these genes should be under strict temporal regulation so that only at particular time or stage the gene would be silenced. For tissue specific or temporal gene regulation, conditional gene silencing strategies are generally adopted. This would enable to customize the induction or silencing of a particular gene at particular tissue at specified time as per one’s desire.
Although the therapeutic proteins, derived from transgenic animals are well accepted by many people, using transgenic technology for improvements in animal productivity is not still accepted in many countries because of public concerns about Genetically Modified (GM) organisms. It is hoped that more the public would be aware of scientific reality about transgenic technology more would be the general acceptance of GM organisms. Considering the rapid advancement in gene targeting and transgenic technology, it is expected that the concept of ‘an animal for all purpose’ would be reality in the near future.
ACKNOWLEDGMENTS
The authors acknowledge the financial support provided by Department of Biotechnology (DBT), Government of India; National Agricultural Science Fund (NASF), Indian Council of Agricultural Research (ICAR), New Delhi and Indian Council of Medical Research (ICMR), New Delhi.
REFERENCES
- Fire, A., S. Xu, M.K. Montgomery, S.A. Kostas, S.E. Driver and C.C. Mello, 1998. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature, 391: 806-811.
CrossRefPubMedDirect Link - Ogden, K.M. and B.V.V. Prasad, 2015. Quelling an innate response to dsRNA. Oncotarget, 6: 28535-28536.
CrossRefDirect Link - Sioud, M., 2015. RNA interference: Mechanisms, technical challenges and therapeutic opportunities. Methods Mol. Biol., 1218: 1-15.
CrossRefDirect Link - Fischer, S.E.J., 2015. RNA interference and microRNA-mediated silencing. Curr. Protoc. Mol. Biol., Vol. 112.
CrossRef - Lima, W.F., C.L. de Hoyos, X.H. Liang and S.T. Crooke, 2016. RNA cleavage products generated by antisense oligonucleotides and siRNAs are processed by the RNA surveillance machinery. Nucleic Acids Res., 44: 3351-3363.
CrossRefDirect Link - Zhuang, J.J., S.A. Banse and C.P. Hunter, 2013. The nuclear argonaute NRDE-3 contributes to transitive RNAi in Caenorhabditis elegans. Genetics, 194: 117-131.
CrossRefDirect Link - Ding, S.W. and O. Voinnet, 2007. Antiviral immunity directed by small RNAs. Cell, 130: 413-426.
CrossRefDirect Link - Malone, C.D. and G.J. Hannon, 2009. Small RNAs as guardians of the genome. Cell, 136: 656-668.
CrossRefDirect Link - Nowacki, M., V. Vijayan, Y. Zhou, K. Schotanus, T.G. Doak and L.F. Landweber, 2008. RNA-mediated epigenetic programming of a genome-rearrangement pathway. Nature, 451: 153-158.
CrossRefDirect Link - Mochizuki, K., 2012. Developmentally programmed, RNA-directed genome rearrangement in Tetrahymena. Dev. Growth Differ., 54: 108-119.
CrossRefDirect Link - Joshi, B.H. and K.P. Pachchigar, 2014. siRNA: Novel therapeutics from functional genomics. Biotechnol. Genet. Eng. Rev., 30: 1-30.
CrossRefDirect Link - Brummelkamp, T.R., R. Bernards and R. Agami, 2002. A system for stable expression of short interfering RNAs in mammalian cells. Science, 296: 550-553.
CrossRefPubMedDirect Link - Paddison, P.J., A.A. Caudy, E. Bernstein, G.J. Hannon and D.S. Conklin, 2002. Short hairpin RNAs (shRNAs) induce sequence-specific silencing in mammalian cells. Genes Dev., 16: 948-958.
CrossRefDirect Link - Paul, C.P., P.D. Good, I. Winer and D.R. Engelke, 2002. Effective expression of small interfering RNA in human cells. Nat. Biotechnol., 20: 505-508.
CrossRefPubMedDirect Link - Carmell, M.A., L. Zhang, D.S. Conklin, G.J. Hannon and T.A. Rosenquist, 2003. Germline transmission of RNAi in mice. Nat. Struct. Mol. Biol., 10: 91-92.
CrossRefDirect Link - Guo, Z.M., K. Xu, Y. Yue, B. Huang and X.Y. Deng et al., 2005. Temporal control of Cre recombinase-mediated in vitro DNA recombination by Tet-on gene expression system. Acta Biochimica Biophysica Sinica, 37: 133-138.
CrossRefDirect Link - Allison, S.J. and J. Milner, 2014. RNA Interference by single- and double-stranded siRNA with a DNA extension containing a 3′ nuclease-resistant mini-hairpin structure. Mol. Therapy Nucl. Acids, Vol. 3.
CrossRefDirect Link - Yarmolinsky, M. and R. Hoess, 2015. The legacy of nat sternberg: The genesis of Cre-lox technology. Annu. Rev. Virol., 2: 25-40.
CrossRefDirect Link - Lacroix, C., D. Giovannini, A. Combe, D.Y. Bargieri and S. Spath et al., 2011. FLP/FRT-mediated conditional mutagenesis in pre-erythrocytic stages of Plasmodium berghei. Nat. Protocols, 6: 1412-1428.
CrossRefDirect Link - Matsukura, S., P.A. Jones and D. Takai, 2003. Establishment of conditional vectors for hairpin siRNA knockdowns. Nucl. Acids Res., Vol. 31.
CrossRefDirect Link - Nielsen, T.T., I. van Marion, L. Hasholt and C. Lundberg, 2009. Neuron-specific RNA interference using lentiviral vectors. J. Gene Med., 11: 559-569.
CrossRefDirect Link - Allen, D., P.F. Kenna, A. Palfi, H.P. McMahon and S. Millington-Ward et al., 2007. Development of strategies for conditional RNA interference. J. Gene Med., 9: 287-298.
CrossRefDirect Link - Horvat, S. and J.F. Medrano, 2001. Lack of Socs2 expression causes the high-growth phenotype in mice. Genomics, 72: 209-212.
CrossRefDirect Link - Lindeman, G.J., S. Wittlin, H. Lada, M.J. Naylor and M. Santamaria et al., 2001. SOCS1 deficiency results in accelerated mammary gland development and rescues lactation in prolactin receptor-deficient mice. Genes Dev., 15: 1631-1636.
CrossRefDirect Link - Rodriguez, J., B. Vernus, I. Chelh, I. Cassar-Malek and J.C. Gabillard et al., 2014. Myostatin and the skeletal muscle atrophy and hypertrophy signaling pathways. Cell. Mol. Life Sci., 71: 4361-4371.
CrossRefDirect Link - Luo, J., Z. Song, S. Yu, D. Cui and B. Wang et al., 2014. Efficient generation of myostatin (MSTN) biallelic mutations in cattle using zinc finger nucleases. PloS ONE, Vol. 9.
CrossRefDirect Link - Hu, S., W. Ni, W. Sai, H. Zi and J. Qiao et al., 2013. Knockdown of myostatin expression by RNAi enhances muscle growth in transgenic sheep. PLoS ONE, Vol. 8.
CrossRefDirect Link - Dushyanth, K., T.K. Bhattacharya, R. Shukla, R.N. Chatterjee, T. Sitaramamma, C. Paswan and P.G. Vishnu, 2016. Gene expression and polymorphism of Myostatin gene and its association with growth traits in chicken. Anim. Biotechnol., 27: 269-277.
CrossRefDirect Link - Jabed, A., S. Wagner, J. McCracken, D.N. Wells and G. Laible, 2012. Targeted microRNA expression in dairy cattle directs production of β-lactoglobulin-free, high-casein milk. Proc. Natl. Acad. Sci. USA., 109: 16811-16816.
CrossRefDirect Link - Karatzas, C.N. and J.D. Turner, 1997. Toward altering milk composition by genetic manipulation: Current status and challenges. J. Dairy Sci., 80: 2225-2232.
CrossRefDirect Link - Stinnakre, M.G., J.L. Vilotte, S. Soulier and J.C. Mercier, 1994. Creation and phenotypic analysis of alpha-lactalbumin-deficient mice. Proc. Natl. Acad. Sci. USA., 91: 6544-6548.
Direct Link - Vilotte, J.L., 2002. Lowering the milk lactose content in vivo: Potential interests, strategies and physiological consequences. Reprod. Nutr. Dev., 42: 127-132.
CrossRefDirect Link - Soulier, S., L. Lepourry, M.G. Stinnakre, J.C. Mercier and J.L. Vilotte, 1997. Expression of a bovine α-lactalbumin transgene in α-lactalbumin-deficient mice can rescue lactation. In vivo relationship between bovine α-lactalbumin expression content and milk composition. J. Dairy Res., 64: 145-148.
Direct Link - Laible, G., 2009. Enhancing livestock through genetic engineering-recent advances and future prospects. Comp. Immunol. Microbiol. Infect. Dis., 32: 123-137.
CrossRefDirect Link - Lai, L., D. Kolber-Simonds, K.W. Park, H.T. Cheong and J.L. Greenstein et al., 2002. Production of α-1,3-galactosyltransferase knockout pigs by nuclear transfer cloning. Science, 295: 1089-1092.
CrossRefDirect Link - Dai, Y., T.D. Vaught, J. Boone, S.H. Chen and C.J. Phelps et al., 2002. Targeted disruption of the α1,3-galactosyltransferase gene in cloned pigs. Nat. Biotecnol., 20: 251-255.
CrossRefPubMedDirect Link - Liu, Y., J.Y. Yang, Y. Lu, P. Yu and C.R. Dove et al., 2013. α-1,3-galactosyltransferase knockout pig induced pluripotent stem cells: A cell source for the production of xenotransplant pigs. Cell. Reprogramm., 15: 107-116.
Direct Link - Denning, C., S. Burl, A. Ainslie, J. Bracken and A. Dinnyes et al., 2001. Deletion of the α(1,3)galactosyl transferase (GGTA1) gene and the prion protein (PrP) gene in sheep. Nat. Biotechnol., 19: 559-562.
CrossRefDirect Link - Golding, M.C., C.R. Long, M.A. Carmell, G.J. Hannon and M.E. Westhusin, 2006. Suppression of prion protein in livestock by RNA interference. Proc. Natl. Acad. Sci. USA., 103: 5285-5290.
CrossRefDirect Link - Wongsrikeao, P., S. Sutou, M. Kunishi, Y.J. Dong, X. Bai and T. Otoi, 2011. Combination of the somatic cell nuclear transfer method and RNAi technology for the production of a prion gene-knockdown calf using plasmid vectors harboring the U6 or tRNA promoter. Prion, 5: 39-46.
CrossRefDirect Link - Subramanya, S., S.S. Kim, N. Manjunath and P. Shankar, 2010. RNA interference-based therapeutics for human immunodeficiency virus HIV-1 treatment: Synthetic siRNA or vector-based shRNA? Expert Opin. Biol. Ther., 10: 201-213.
CrossRefDirect Link - Sabin, L.R., R. Zhou, J.J. Gruber, N. Lukinova and S. Bambina et al., 2009. Ars2 regulates both miRNA- and siRNA-dependent silencing and suppresses RNA virus infection in Drosophila. Cell, 138: 340-351.
CrossRefDirect Link - Schwegmann-Wessels, C., S. Bauer, C. Winter, L. Enjuanes, H. Laude and G. Herrler, 2011. The sialic acid binding activity of the S protein facilitates infection by porcine transmissible gastroenteritis coronavirus. Virol. J., Vol. 8.
CrossRefDirect Link - Clark, J. and B. Whitelaw, 2003. A future for transgenic livestock. Nat. Resour. Genet., 4: 825-833.
CrossRefDirect Link - Piedrahita, J.A. and N. Olby, 2011. Perspectives on transgenic livestock in agriculture and biomedicine: An update. Reprod. Fertil. Dev., 23: 56-63.
CrossRefPubMedDirect Link - Belfort, M. and R.P. Bonocora, 2014. Homing endonucleases: From genetic anomalies to programmable genomic clippers. Methods Mol. Biol., 1123: 1-26.
CrossRefDirect Link - Jabalameli, H.R., H. Zahednasab, A. Karimi-Moghaddam and M.R. Jabalameli, 2015. Zinc finger nuclease technology: Advances and obstacles in modelling and treating genetic disorders. Gene, 558: 1-5.
CrossRefDirect Link - Wright, D.A., T. Li, B. Yang and M.H. Spalding, 2014. Talen-mediated genome editing: Prospects and perspectives. Biochem. J., 462: 15-24.
CrossRefDirect Link - Singh, V., D. Braddick and P.K. Dhar, 2017. Exploring the potential of genome editing CRISPR-Cas9 technology. Gene, 599: 1-18.
CrossRefDirect Link