
M. Alagawany
Department of Poultry, Faculty of Agriculture, Zagazig University, 44511 Zagazig, Egypt
LiveDNA: 20.7214
Mayada R. Farag
Department of Forensic Medicine and Toxicology, Faculty of Veterinary Medicine, Zagazig University, 44511 Zagazig, Egypt
M.S. El-Kholy
Department of Poultry, Faculty of Agriculture, Zagazig University, 44511 Zagazig, Egypt
S.A.A. El-Sayed
Department of Nutrition and Clinical Nutrition, Faculty of Veterinary Medicine, Zagazig University, P.O. Box 44511, Zagazig, Egypt
K. Dhama
Division of Pathology, Indian Veterinary Research Institute, Izatnagar, Bareilly, 243122 Uttar Pradesh, India
LiveDNA: 91.4710
Asian Journal of Animal and Veterinary Advances
Year: 2017 | Volume: 12 | Issue: 1 | Page No.: 1-9







ABSTRACT
Objective: Three separate experiments were performed to evaluate the biological in vitro effects of two natural antioxidants, resveratrol (RES), cinnamaldehyde (CIN) and their combinations (RES+CIN) on rabbit erythrocytes. Materials and Methods: Suspensions of erythrocytes were divided into 15 groups (each group contains 5 replicates). In three experiments, control group was treated with phosphate buffered saline and dimethyl sulfoxide (DMSO) under the same concentration, while the phytochemical groups in the three experiments were 25, 50, 75 and 100 μM of RES or CIN or their combinations (equal mixture of RES and CIN) added to the control group, respectively and incubated for 3 h at 37°C and exposed to gentle shaking every 15 min. Results: The RES or CIN separately or in combination at different doses enhanced the activities of superoxide dismutase and catalase and improved the concentration of reduced glutathione compared to the control group. Malondialdehyde, protein carbonyl and adenosine triphosphate contents showed linear and quadratic decrease with increasing the concentration of RES or CIN or their combinations. Conclusion: The tested bioactive compounds (RES+CIN) could be helpful in improving the body cells functions without causing hazardous effects and efficiently protected the cells against oxidative injury when used at concentrations up to 100 μM and found to have synergistic effects.
PDF Abstract XML References Citation
How to cite this article
M. Alagawany, Mayada R. Farag, M.S. El-Kholy, S.A.A. El-Sayed and K. Dhama, 2017. Effect of Resveratrol, Cinnamaldehyde and their Combinations on the Antioxidant Defense System and ATP Release of Rabbit Erythrocytes: In vitro Study. Asian Journal of Animal and Veterinary Advances, 12: 1-9.
DOI: 10.3923/ajava.2017.1.9
URL: https://scialert.net/abstract/?doi=ajava.2017.1.9
DOI: 10.3923/ajava.2017.1.9
URL: https://scialert.net/abstract/?doi=ajava.2017.1.9
INTRODUCTION
Diet rich in herbs or herbal extracts provides animals with antioxidant polyphenolic phytochemicals which are supposed to exert positive effect on production and animal health1,2. One of these effective compounds is resveratrol (3,5,4'-trihydroxy-trans-stilbene, RES) which is found predominantly as an aromatic phytoalexin of stilbenes type in grapes, peanuts, berries and yucca, also produced naturally by several plants in response to injury or when the plant is under attack by pathogens like fungi or bacteria3. The other well-known molecule: Cinnamaldehyde (3-phenyl-2-propenal, CIN) is present mostly in some herbs such as cinnamon4.
The main application of RES and CIN is in poultry and animal nutrition in particular as feed additives to reduce free radicals in a wide variety of animal species. Several studies carried out on supplementing diets with feed additives rich in natural antioxidants as RES and CIN demonstrated their capability to improve the performance, immunity and health of livestock besides protecting animals against various degenerative diseases and different types of cancer2,5. Numerous in vivo and in vitro reports declared the beneficial impacts of RES, such as anti-aging, cardio-protective, antioxidant, anticancer, anti-inflammatory, immunomodulatory as well as chemopreventive and metabolic modifier activities that are considered in the treatment and management of a vast array of human diseases6. Hao et al.7 stated that supplementation of RES (5, 22.5 and 45 mg kg–1 diet) could decrease malonaldehyde (MDA) concentration in the liver of animal with high fat and high cholesterol diets. Also, the levels of glutathione peroxidase (GSH-Px), superoxide dismutase (SOD) were increased with resveratrol groups. Blood MDA concentration was decreased with increasing resveratrol supplementation in layer quail diets from 200-400 mg kg–1 diet1. Similarly, cinnamaldehyde showed antimicrobial effect, vasodilatory and anti-platelet aggregating activities and antitumor actions8.
To our knowledge, there are limited studies on the effect of RES and CIN or their combination on the antioxidant defense system of the erythrocytes. Thus, the aim of the current study is to assess the potential effects of RES, CIN separately or in combination as natural antioxidant agents at different concentrations (0, 25, 50, 75 and 100 μM) on SOD, catalase (CAT) and reduced glutathione (GSH) of isolated rabbit erythrocytes in addition to its effects on cell status explored by the hemoglobin and adenosine triphosphate (ATP) content on hemolysate as markers for hemolysis, besides investigating the influence of RES, CIN and their combination (RES+CIN) on the protein and lipid oxidation extent in erythrocytes. Since, SOD-CAT system is the rst line of defense against peroxidative injury and is indicative of the rate of free radical formation and due to the important function of reduced glutathione in cellular transformation9.
MATERIALS AND METHODS
Chemicals: The RES (3,5,4'-trihydroxy-trans-stilbene, purity of 99%) and pure CIN (3-phenyl-2-propenal, purity ≥98%) were obtained from Oxford Laboratory Mumbai, India. All other chemicals were obtained from Sigma (St., Louis, MO, USA). Kits of antioxidants were obtained from Biodiagnostic, BD and Egypt.
Animals and care: Male New Zealand white rabbits (3 months of age and initial weight of 2.000±50 g) were used. Animals were individually housed in stainless steel cages at room temperature (25±2°C) with a relative humidity of 50-60% and on a 12 h light-darkness cycle. The rabbits had free access to commercial pellet diet and water ad libitum. The animals’ welfare and care were conformed to the guidelines of the Animal Use Research Ethics Committee of Zagazig University, Egypt.
Preparation of erythrocytes: Blood samples (5 mL each) were collected aseptically from the ear vein of all rabbits in heparin containing test-tubes to avoid coagulation. The collected blood samples were centrifuged for 10 min at 3000 rpm at 4°C and then plasma and buffy coat were removed carefully. The erythrocytes were washed once with 0.9% NaCl solution then harvested by centrifugation then washed 2 times with ice-cold phosphate buffered saline (PBS: 145 mM NaCl, 1.9 mM NaH2PO4, 8.1 mM Na2HPO4) and finally resuspended in PBS to obtain cell suspensions at 10% hematocrit to be used for incubations10.
Treatment of erythrocytes: The RES or CIN was primarily solubilized alone small amounts of dimethyl sulfoxide (DMSO) (not exceed 0.1%) with further dilution in PBS till the required concentrations (0, 25, 50, 75 and 100 μM). Three separate experiments were carried out to investigate the biological in vitro effect of RES, CIN and RES+CIN as natural antioxidants on isolated rabbit erythrocytes. The experiments were performed simultaneously and were each designed as a completely randomized design. Suspended erythrocytes were divided into 15 groups (5 replicates/group). In three experiments, the control group treated with PBS and DMSO within the indicated concentration, while the bioactive compound groups in experiment 1, 2 and 3 containing 25, 50, 75 and 100 μM of RES or CIN or RES+CIN (equal mixture of RES and CIN) added to the control group respectively and incubated at 37°C and exposed to gentle shaking every 15 min for 3 h. After incubation, the mixtures were stored at 4°C and used within 6 h of preparation. For antioxidant indices, lipid and protein oxidation markers, the mixtures were stored after incubation at -20°C and thawed 1 day later for RBCs lysis by osmotic pressure, then they were centrifuged and supernatants obtained.
Investigating the antioxidant defense system in erythrocytes
Antioxidant indices: The catalase activity (CAT), superoxide dismutase activity (SOD), reduced glutathione (GSH) and MDA contents were measured using commercial Biodiagnostic kits and a spectrophotometer (Shimadzu, Japan).
Protein oxidation: Protein oxidation was assessed by measuring the content of protein carbonyl (PrC) according to Uchida and Stadtman11. The results were expressed as μmol of 2,4-DNPH-incorporated mg–1 protein based on a molar extinction coefficient of 2.1×104 M–1 cm–1 for aliphatic hydrazones.
Hemoglobin and protein determination: The hemolysis process of erythrocytes was monitored by hemoglobin (Hb) release. After incubation time periods, 50 mL of cell suspension was added to 1 mL of PBS and centrifuged (1000 g, 7 min). The Hb content of supernatants was measured by absorbance at 540 nm using the Varian Cary 50 UV-vis spectrophotometer. Total Protein (TP) concentration in the hemolysate was determined by standard kits as described by the manufacturer companies.
Measurement of ATP content in hemolysate: The procedure for measurement of ATP content released extracellular in the hemolysate was measured according to the method of Adams12. Then ATP expressed as μmol g–1 Hb.
Statistical analysis: Data were statistically analyzed by ANOVA using GLM procedure of SAS13 as a completely randomized design. The statistical model included treatment for all experiments. Orthogonal polynomial contrasts were used to test the linear and quadratic impacts of the gradually increased concentrations of RES, CIN or RES+CIN supplementation to rabbit erythrocytes in experiments 1, 2 and 3, respectively.
RESULTS
Experiment 1: The influence of RES on antioxidant indices of the normal rabbit erythrocytes incubated with indicated concentrations for 3 h is presented in Table 1. Different concentrations of RES (25, 50, 75 or 100 μM) enhanced SOD and CAT activities and increased GSH concentration compared to the control group. Activity of CAT was increased linearly (p<0.001) by increasing the concentration of RES from 0-100 μM. Similarly, the activity of SOD was improved linearly and quadratically (p<0.001) by elevating the concentration of RES from 25-100 μM, where 100 μM increased SOD activity to be (1074 IU g–1 Hb) compared to control value (879 IU g–1 Hb). The same effects of concentration on SOD activity were observed for GSH concentration.
Table 1 shows that, lipid peroxidation (LPO) measured by MDA concentration and Protein Oxidation (PO) represented by PrC content were significantly affected by the used concentrations of RES (25, 50, 75 and 100 μM). The PrC and MDA contents were decreased linearly and quadratically (p<0.001) with increasing the concentration of RES. The lowest PrC and MDA concentrations obtained after incubation with 100 μM of RES for 3 h (1.20 and 3.17 μmol g–1 Hb) compared to untreated group (2.10 and 5.70 μmol g–1 Hb), respectively and compared with the other treated groups.
The effects of RES on ATP, TP and Hb contents in isolated rabbit erythrocyte are shown in Table 2. The RES addition to rabbit erythrocyte evoked a significant linear and quadratic decrease in ATP content (p<0.001 or = 0.020), respectively compared with the non-supplemented group or 25 μM of RES. The lowest value (2.98 μmol g–1 Hb) of ATP was achieved by 100 µM of RES supplementation.
| Table 1: | Effects of resveratrol incubated in rabbit erythrocyte on antioxidant indices and lipid and protein peroxidation in experiment 1 |
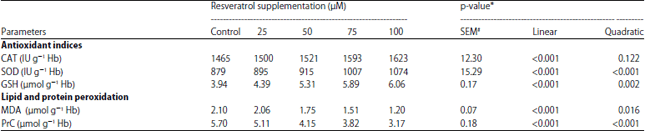 | |
| #SEM: Standard error means, *Linear and quadratic effect of RES supplementation | |
| Table 2: | Effects of resveratrol incubated in rabbit erythrocyte on hemolysis parameters in experiment 1 |
 | |
| #SEM: Standard error means, *Linear and quadratic effect of RES supplementation | |
| Table 3: | Effects of cinnamaldehyde incubated in rabbit erythrocyte on antioxidant indices and lipid and protein peroxidation in experiment 2 |
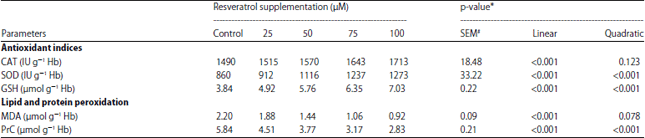 | |
| #SEM: Standard error means, *Linear and quadratic effect of CIN supplementation | |
| Table 4: | Effects of cinnamaldehyde incubated in rabbit erythrocyte on hemolysis parameters in experiment 2 |
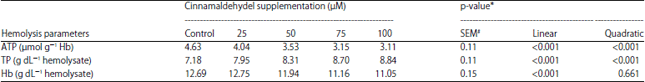 | |
| #SEM: Standard error means, *Linear and quadratic effect of CIN supplementation | |
The TP content in hemolysate was linearly (p<0.001) and quadratically (p = 0.028) elevated by increasing the level of RES to record the highest value (8.34 g dL–1 hemolysate) at 100 μM of RES compared to control (7.23 g dL–1 hemolysate) or other treatments. The Hb content was linearly and quadratically (p<0.001) affected by RES supplementation. The highest Hb content (13.51 g Hb dL–1 hemolysate) was obtained with 25 μM of RES compared to the control and other treated groups.
Experiment 2: The influences of CIN at different doses on antioxidant defense system in the erythrocytes hemolysate are represented in Table 3. The CIN supplementation improved the activities of SOD and CAT and concentration of GSH compared to the control group. The CAT activity was linearly (p<0.001) increased when CIN concentration increased from 0-100 μM. Activity of SOD and GSH content were linearly and quadratically (p<0.001) increased with increasing the levels of CIN from 0-100 μM, where 100 μM increased the activity of SOD and the GSH level to be (1273 IU g–1 Hb and 7.03 μmol g–1 Hb) compared to control value (879 IU g–1 Hb and 3.94 μmol g–1 Hb), respectively.
The MDA and PrC content were linearly (p<0.001) and quadratically (p = 0.078 or <0.001, respectively) decreased with increasing the concentration of CIN (Table 3). Since, the lowest MDA and PrC concentrations obtained with 100 μM of CIN (0.92 and 2.83 μmol g–1 Hb) compared to untreated group (2.10 and 5.70 μmol g–1 Hb) respectively and compared to the other treated groups.
In Table 4, CIN addition to rabbit erythrocytes evoked a significant quadratic and linear decrease in ATP content (p<0.001), compared with the non-supplemented group. The lowest value (3.11 μmol g–1 Hb) of ATP was recorded by 100 μM of CIN supplementation. The concentration of TP in hemolysate was linearly and quadratically (p<0.001) increased with increasing the concentration of CIN to record the highest value (8.84 g dL–1 hemolysate) at 100 μM after 3 h of incubation comparing with control group (7.23 g dL–1 hemolysate) or other treatments. The Hb level was statistically (linearly, p = 0.022) reduced with CIN addition by 50, 75 and 100 μM in comparison with control or 25 μM of CIN.
Experiment 3: The effects of RES+CIN on antioxidant parameters in rabbit erythrocyte are summarized in Table 5. RES+CIN supplementation improved the activities of CAT and SOD as well as GSH concentration compared to control group.
| Table 5: | Combined effect of resveratrol (RES) and cinnamaldehyde (CIN) incubated in rabbit erythrocyte on antioxidant indices and lipid and protein peroxidation in experiment 3 |
 | |
| #SEM: Standard error means, *Linear and quadratic effect of RES+CIN supplementation | |
| Table 6: | Combined effect of resveratrol (RES) and cinnamaldehyde (CIN) incubated in rabbit erythrocyte on hemolysis parameters in experiment 3 |
 | |
| #SEM: Standard error means, *Linear and quadratic effect of RES+CIN supplementation | |
The CAT and SOD activities and GSH level were quadratically and linearly (p<0.001) increased with increasing the concentration of RES+CIN from 0-100 μM.
Linear and quadratic (p<0.001) decrease were detected in MDA and PrC contents with increasing the concentration of RES+CIN (Table 5). Since, the lowest values of MDA and PrC obtained with 100 μM of RES+CIN (0.54 and 2.54 μmol g–1 Hb) compared to untreated group (2.10 and 5.70 μmol g–1 Hb) respectively and compared to other treated groups.
Table 6 shows the RES+CIN addition to rabbit erythrocyte caused a significant linear and quadratic decrease in ATP content (p<0.001), compared to un-supplemented group. The lowest value (2.65 μmol g–1 Hb) of ATP was recorded by 100 μM of RES+CIN supplementation. The concentration of TP in hemolysate was linearly and quadratically (p<0.001) elevated with increasing the concentration of RES+CIN to achieve the highest value (9.12 g dL–1 hemolysate) at 100 μM comparing with control group (7.23 g dL–1 hemolysate) or other treatments. On the contrary, Hb level was statistically (linearly, p<0.001) lowered with RES+CIN addition compared to the control group.
DISCUSSION
Natural antioxidants and feed additives are reported in some in vivo and in vitro studies to alleviate oxidative stress and scavenging Reactive Oxygen Species (ROS) however, these kinds of additives showed some pro-oxidant activities and can induce apoptosis in cancer cells. In this regard, the antioxidant and pro-oxidant properties of these phytochemicals in animal feed should be considered with care and their combined effect should be characterized with some special interest. Therefore, in the current experiments we tried to investigate the using of RES, CIN and RES+CIN in isolated rabbit erythrocytes (as model for biological lipid membranes) at different concentrations that could be helpful in improving the actions of body cells without causing hazardous impacts.
In the current experiments, the most obvious findings are that the different doses of RES or CIN or their combinations (25, 50, 75 and 100 μM) displayed a positive effect on enhancing SOD and CAT activities as well as GSH and TP concentrations after incubation with additives for 3 h. On the contrary, increasing the concentrations of phytogenic products significantly reduced the MDA and PrC concentrations in addition to ATP and Hb contents of erythrocytes suggesting the safety of these concentrations on the lipid membrane of erythrocytes.
The results concerning the effects of RES on erythrocyte antioxidant defense system could be explained by the opinion of Lopez-Velez et al.14 who stated that resveratrol could provide cell protection due to antioxidant capacity, it could act directly as an effective eliminator of superoxide, hydroxyl and metal-induced free radicals and through balancing of hydroxyl phenolic groups. The other possible mechanism involved in resveratrol’s antioxidant activity is increasing the antioxidant enzymes activities like CAT and SOD, glutathione S-transferase and NADPH quinine oxido-reductase as described by Young et al.15. In experiment 1, our results are also comparable with Ungvari et al.16 who observed a free radical capturing effect of 1, 10 and 100 μM resveratrol in a cell-free assay. The antioxidant impacts of resveratrol has been also reported by Rubiolo and Vega17 in primary rat hepatocytes where resveratrol decreases cell death for up to 24 h by enhancing the activates of erythroid-derived nuclear factor, a key transcription factor regulating the response of antioxidant. Similarly, Sahin et al.1 pointed out that diet supplemented with resveratrol decreased the level of MDA and increased the enzyme activities such as CAT and SOD in the serum.
Resveratrol has been also suggested to exert its antioxidant protection effects through decreasing the generation of ROS as described in some previous studies such as that of Li et al.18 in which pretreatment with 25-100 μM resveratrol for 72 h reduced accumulation of ROS in the rat aortic smooth muscle cells in vitro. Similarly, 50 μM resveratrol protected primary cortical astrocyte cultures from oxidative damage caused by H2O2 by inhibiting intracellular ROS production19. Pre-incubation of the human primary small airway epithelial and human alveolar epithelial (A549) cells with 10 μM resveratrol for 24 h decreased Cigarette Smoke Extract (CSE) induced generation of ROS20. On the same context, pre-incubation of Human Coronary Artery Endothelial Cells (HCAECs) with 10 and 50 μM resveratrol for different periods (15 and 60 min, 6 h). Provide an in vitro protection against oxidative-stress-induced by H2O2 and decrease cell, indicating the free radical scavenging capability of resveratrol21. Das22 found that resveratrol as a natural antioxidant scavenges ROS and protects the organism from oxidative stress where ROS can react with macromolecules like nucleic acids, proteins and lipids resulting in alterations in the structure and activity of biologically relevant molecules causing cell damage. There are some in vivo studies concerning the antioxidant activity of resveratrol for example, the supplementation of resveratrol in broiler diets helped in enhancing the oxidative enzymes activities or improving the antioxidant status of the birds and the total antioxidant capacity in plasma23. Such antioxidant effects would be expected to improve the health of animal. Additionally, supplementation of chicken’s diet with resveratrol resulted in a linear increase in serum GSH, growth hormone and insulin-like growth factor-1 levels, in addition to the activities of GSH-Px, SOD and CAT significantly decreased the serum MDA concentration compared with chickens that were fed diets without resveratrol during heat stress24. Our results obtained from experiment 1 are also in agreement with Mikstacka et al.25 where resveratrol prevented the reduction in GSH content and reduced lipid peroxidation in human erythrocytes in a concentration-dependent manner. Brito et al.26 also stated that in vitro pre-incubation of bovine aortic endothelial cells for 14 h with resveratrol at 50 μM protects the cells from peroxynitrite-mediated cell death via an elevation in the intracellular GSH. Resveratrol can also act by reduction or prevention of the apolipoprotein B peroxidation accompanied with low density lipoprotein27.
In a like manner, CIN enhanced the activities of SOD and CAT as well as GSH concentration and significantly reduced the MDA and PrC concentrations in a dose dependent manner in the experiment 2. These findings are in agreement with Subash-Babu et al.28 who demonstrated that cinnamaldehyde enhances the activity of antioxidant defense system against reactive oxygen species produced under hyperglycemic conditions in animal providing protection to pancreatic β-cells. These impacts may be returned to the ability of cinnamaldehyde to act as a potential source of antioxidant scavenging free radicals as it exhibited radical scavenging activities in different in vitro models like DPPH, superoxide, nitric oxide, H2O2 scavenging activity and reducing power29. Cinnamaldehyde also exhibit strong antioxidant capacity to scavenge free radicals of oxygen and lipids as reported by Mathew and Abraham30.
The obtained improvement in SOD and CAT observed in these studies after addition of RES and CIN separately or in combination suggest their benefits in keeping the normal physiological conditions of the erythrocytes that are required for such enzymes to exert their free radical scavenging role in eliminating ROS, therefore, protect the cells against oxidative damage providing cell protection against oxidative stress as stated by Evans and Halliwell31. It is well known that, the antioxidant enzyme SOD is distributed widely in all cells and high amount of it presents in erythrocytes32. The SOD protects cell against oxidative damage by converting the highly reactive superoxide anion to O2 and to a less reactive species33, H2O2. While, CAT present in erythrocytes in the form of soluble protein that protect hemoglobin from peroxidation as CAT is required for inhibiting the formation of deleterious H2O2 in cells34. So increasing their activities is suggestive for the antioxidant properties of the different doses of phytochemical additives.
Phytochemicals (RES and CIN) also induced elevation of the GSH level which provide the primary antioxidant defense for the stored erythrocytes, protect membrane lipids and proteins and keep membrane skeleton stability and survival of erythrocytes against peroxidative stress. It is also well established that, in RBCs, GSH protects important proteins against oxidation and that it is a vital sulfhydryl buffer, maintaining SH groups in Hb and enzymes in the reduced state35. Oxidant scavenging at the intracellular level within the cytosol appears to rely on reduced glutathione and glutathione peroxidase for elimination of low micromolar levels of lipid hydroperoxides and hydrogen peroxides36. The GSH oxidation can be induced directly due to the attack of free radicals or indirectly through consumption of GSH during repair processes such as the reduction of oxidized membrane-protein thiol groups37. Moreover, reduced glutathione could bind with MDA and other deleterious endogenous substances beside its crucial role in cellular transformation9. This could explain the results of this study concerning the impacts of RES and CIN or their combinations on MDA concentration, where MDA concentration was lowered with increasing the concentration of RES or CIN or RES+CIN. Hence, lower level of this compound may improve the steady state of antioxidant system in animal. The MDA, the marker of LPO of erythrocytes is a highly reactive bifunctional molecule, that cross-link erythrocyte proteins and phospholipids to alter the functions of cell membrane leading to decrease erythrocytic survival and has been proposed as a general mechanism for cell injury and death i.e., induce hemolysis38. This comes on line with the results obtained in the present studies regarding the impact of RES and CIN on Hb and ATP content of the hemolysate. The ATP is used by erythrocytes to maintain membrane shape, control deformation and maintain osmotic stability as well as sub-membrane skeletal-network proteins36. The basal stimulated ATP-release in the present studies found to be correlated tightly with extracellular hemoglobin, a marker of hemolysis. This is in agreement with Sikora et al.39 who demonstrated that the source of extracellular ATP was cell lysis. These closely related effects could be observed by the opinion of Banerjee et al.40 who stated that as a result of LPO, the erythrocyte membrane becomes weak and loses its integrity rapidly leading to Hb release. The present findings supported those of May41 that cell membrane is often the initial site of injury and peroxidation of membrane lipids causes hemolysis and alter the protein and lipid content to different extents.
Erythrocyte SOD contains copper and zinc (CuZn-SOD). Zinc keeps the stability of the enzyme, while copper maintains its activity35. The interaction between MDA and CuZn-SOD leads to the modi cation of histidine amino acid residues and the production of protein-protein cross-linked derivatives as a result each type of ROS gives a different protein oxidation pattern42. This could explain the generation of protein carbonyl derivatives which may be the main cause of the reduced protein content of the rabbit erythrocytes after incubation with RES or CIN or their combinations on a dose-dependent manner.
CONCLUSION
The impacts noted in experiments indicated that RES, CIN or RES+CIN as natural phytogenic additives could be helpful in improving the function of the body cells without causing hazardous effects and efficiently protect the cells against oxidative injury when used at concentrations up to 100 μM and found to have synergistic effects (Experiment 3). These findings may be attributed to the possibility of these types of phytochemicals to improve the antioxidant indices (CAT, SOD and GSH) as well as TP and Hb contents. In addition, phytogenic products supplementation to rabbit erythrocyte resulted in a significant decrease in LPO, PO and ATP contents, compared with the un-supplemented group suggesting their role in eliminating ROS that is considered to play a critical role in lipid peroxidation, ageing and the peroxidative hemolysis.
REFERENCES
- Sahin, K., F. Akdemir, C. Orhan, M. Tuzcu, A. Hayirli and N. Sahin, 2010. Effects of dietary resveratrol supplementation on egg production and antioxidant status. Poult. Sci., 89: 1190-1198.
CrossRefDirect Link - Alagawany, M.M., M.R. Farag, K. Dhama, M.E. Abd El-Hack, R. Tiwari and G.M. Alam, 2015. Mechanisms and beneficial applications of resveratrol as feed additive in animal and poultry nutrition: A review. Int. J. Pharmacol., 11: 213-221.
CrossRefDirect Link - Fremont, L., 2000. Biological effects of resveratrol. Life Sci., 66: 663-673.
CrossRefPubMedDirect Link - Faix, S., Z. Faixova, I. Placha and J. Koppel, 2009. Effect of Cinnamomum zeylanicum essential oil on antioxidative status in broiler chickens. Acta. Vet. Brno, 78: 411-417.
Direct Link - Ka, H., H.J. Park, H.J. Jung, J.W. Choi, K.S. Cho, J. Ha and K.T. Lee, 2003. Cinnamaldehyde induces apoptosis by ROS-mediated mitochondrial permeability transition in human promyelocytic leukemia HL-60 cells. Cancer Lett., 196: 143-152.
CrossRefPubMedDirect Link - Frojdo, S., D. Cozzone, H. Vidal and L. Pirola, 2007. Resveratrol is a class IA phosphoinositide 3-kinase inhibitor. Biochem. J., 406: 511-528.
CrossRefPubMedDirect Link - Hao, R.Y., Y.X. Li, F. Yu, Y. Zhao and X.N. Zheng et al., 2011. Effects of resveratrol on lipid peroxidation in mice with high fat and high cholesterol diet. J. China Med. Univ., 40: 17-25.
Direct Link - Wu, S.J., L.T. Ng and C.C. Lin, 2005. Cinnamaldehyde-induced apoptosis in human PLC/PRF/5 cells through activation of the proapoptotic Bcl-2 family proteins and MAPK pathway. Life Sci., 77: 938-951.
CrossRefDirect Link - Bukowska, B., 2003. Effects of 2,4-D and its metabolite 2,4-dichlorophenol on antioxidant enzymes and level of glutathione in human erythrocytes. Comp. Biochem. Physiol. C Toxicol. Pharmacol., 135: 435-441.
CrossRefPubMedDirect Link - Yang, H.L., S.C. Chen, N.W. Chang, J.M. Chang and M.L. Lee et al., 2006. Protection from oxidative damage using Bidens pilosa extracts in normal human erythrocytes. Food Chem. Toxicol., 44: 1513-1521.
CrossRefPubMedDirect Link - Uchida, K. and E.R. Stadtman, 1993. Covalent attachment of 4-hydroxynonenal to glyceraldehyde-3-phosphate dehydrogenase. A possible involvement of intra- and intermolecular cross-linking reaction. J. Biol. Chem., 268: 6388-6393.
Direct Link - Lopez-Velez, M., F. Martinez-Martinez and C. del Valle-Ribes, 2003. The study of phenolic compounds as natural antioxidants in wine. Crit. Rev. Food Sci. Nutr., 43: 233-244.
CrossRefPubMedDirect Link - Young, J.F., L.O. Dragsted, B. Daneshvar, S.T. Lauridsen, M. Hansen and B. Sandstrom, 2000. The effect of grape-skin extract on oxidative status. Br. J. Nutr., 84: 505-513.
Direct Link - Ungvari, Z., Z. Orosz, A. Rivera, N. Labinskyy and Z. Xiangmin et al., 2007. Resveratrol increases vascular oxidative stress resistance. Am. J. Physiol. Heart Circulatory Physiol., 292: H2417-H2424.
CrossRefDirect Link - Rubiolo, J.A. and F.V. Vega, 2008. Resveratrol protects primary rat hepatocytes against necrosis induced by reactive oxygen species. Biomed. Pharm., 62: 606-612.
CrossRefDirect Link - Li, Y., Z. Cao and H. Zhu, 2006. Upregulation of endogenous antioxidants and phase 2 enzymes by the red wine polyphenol, resveratrol in cultured aortic smooth muscle cells leads to cytoprotection against oxidative and electrophilic stress. Pharmacol. Res., 53: 6-15.
CrossRefPubMedDirect Link - Almeida, L.M.V., C.C. Pineiro, M.C. Leite, G. Brolese, R.B. Leal, C. Gottfried and C.A. Goncalves, 2008. Protective effects of resveratrol on hydrogen peroxide induced toxicity in primary cortical astrocyte cultures. Neurochem. Res., 33: 8-15.
CrossRefPubMedDirect Link - Kode, A., S. Rajendrasozhan, S. Caito, S.R. Yang, I.L. Megson and I. Rahman 2008. Resveratrol induces glutathione synthesis by activation of Nrf2 and protects against cigarette smoke-mediated oxidative stress in human lung epithelial cells. Am. J. Physiol. Lung Cell Mol. Physiol., 294: L478-L488.
CrossRefDirect Link - Sayin, O., N. Arslan, Z.S. Altun and G. Akdogan, 2011. In vitro effect of resveratrol against oxidative injury of human coronary artery endothelial cells Turk. J. Med. Sci., 41: 211-218.
Direct Link - Das, A., 2011. Heat stress-induced hepatotoxicity and its prevention by resveratrol in rats. Toxicol. Mech. Methods, 21: 393-399.
CrossRefPubMedDirect Link - Sridhar, M., R.U. Suganthi and V. Thammiaha, 2015. Effect of dietary resveratrol in ameliorating aflatoxin B1-induced changes in broiler birds. J. Anim. Physiol. Anim. Nutr., 99: 1094-1104.
CrossRefDirect Link - Liu, L.L., J.H. He, H.B. Xie, Y.S. Yang, J.C. Li and Y. Zou, 2014. Resveratrol induces antioxidant and heat shock protein mRNA expression in response to heat stress in black-boned chickens. Poult. Sci., 93: 54-62.
CrossRefDirect Link - Mikstacka, R., A.M. Rimando and E. Ignatowicz, 2010. Antioxidant effect of trans-resveratrol, pterostilbene, quercetin and their combinations in human erythrocytes in vitro. Plant Foods Hum. Nutr., 65: 57-63.
CrossRefPubMedDirect Link - Brito, P.M., A. Mariano, L.M. Almeida and T.C. Dinis, 2006. Resveratrol affords protection against peroxynitrite-mediated endothelial cell death: A role for intracellular glutathione. Chemico-Biol. Interact., 164: 157-166.
CrossRefDirect Link - Kubota, S., T. Kurihara, H. Mochimaru, S. Satofuka and K. Noda et al., 2009. Prevention of ocular inflammation in endotoxin-induced uveitis with resveratrol by inhibiting oxidative damage and nuclear factor-κB activation. Invest. Ophthalmol. Visual Sci., 50: 3512-3519.
CrossRefDirect Link - Subash-Babu, P., A.A. Ali and S. Ignacimuthu, 2014. Beneficial antioxidative and antiperoxidative effect of cinnamaldehyde protect streptozotocin-induced pancreatic β-cells damage in wistar rats. Biomol. Ther., 22: 47-54.
CrossRefDirect Link - Haripriya, D., K. Nadhiya and K. Vijayalakshmi, 2013. Antioxidant potential of Cinnamaldehyde: An in vitro study. Int. J. Pharmaceut. Res. Bio-Sci., 2: 270-278.
Direct Link - Mathew, S. and T.E. Abraham, 2006. In vitro antioxidant activity and scavenging effects of Cinnamomum verum leaf extract assayed by different methodologies. Food. Chem. Toxicol., 44: 198-206.
CrossRefDirect Link - Evans, P. and B. Halliwell, 2001. Micronutrients: Oxidant/antioxidant status. Br. J. Nutr., 85: S67-S74.
CrossRefDirect Link - Speranza, M.J., A.C. Bagley and R.E. Lynch, 1993. Cells enriched for catalase are sensitized to the toxicities of bleomycin, adriamycin, and paraquat. J. Biol. Chem., 268: 19039-19043.
CrossRefDirect Link - McCord, J.M., 2000. The evolution of free radicals and oxidative stress. Am. J. Med., 108: 652-659.
CrossRefDirect Link - Aebi, H., 1984. Catalase in vitro. In: Methods in Enzymology, Packer, L., Academic Press, Cambridge, Massachusetts, United States, ISBN: 9780121820053, pp: 121-126.
CrossRefDirect Link - Cimen, M.Y.B., 2008. Free radical metabolism in human erythrocytes. Clinica Chimica Acta, 390: 1-11.
CrossRefDirect Link - Suttorp, N., W. Toepfer and L. Roka, 1986. Antioxidant defense mechanisms of endothelial cells: Glutathione redox cycle versus catalase. Am. J. Physiol.-Cell Physiol., 251: C671-C680.
Direct Link - Wafa, T., N. Amel, C. Issam, C. Imed, M. Abdelhedi and H. Mohamed, 2011. Subacute effects of 2,4-dichlorophenoxyacetic herbicide on antioxidant defense system and lipid peroxidation in rat erythrocytes. Pestic. Biochem. Physiol., 99: 256-264.
CrossRefDirect Link - Sikora, J., S.N. Orlov, K. Furuya and R. Grygorczyk, 2014. Hemolysis is a primary ATP-release mechanism in human erythrocytes. Blood, 124: 2150-2157.
CrossRefPubMedDirect Link - Banerjee, A., A. Kunwar, B. Mishra and K.I. Priyadarsini, 2008. Concentration dependent antioxidant/pro-oxidant activity of curcumin: Studies from AAPH induced hemolysis of RBCs. Chem. Biol. Interact., 174: 134-139.
CrossRefDirect Link - May, J.M., 1998. Ascorbate function and metabolism in the human erythrocyte. Front Biosci., 3: d1-d10.
PubMedDirect Link - Kwon, H.Y., S.Y. Choi, M.H. Won, T.C. Kang and J.H. Kang, 2000. Oxidative modification and inactivation of Cu, Zn-superoxide dismutase by 2,2'-azobis(2-amidinopropane) dihydrochloride. Biochimica Biophysica Acta (BBA)-Protein Struct. Mol. Enzymol., 1543: 69-76.
CrossRefDirect Link