
H.M. Ghanem
Department of Animal Husbandry and Wealth Development, Faculty of Veterinary Medicine, Mansoura University, Egypt
A.I. Ateya
Department of Animal Husbandry and Wealth Development, Faculty of Veterinary Medicine, Mansoura University, Egypt
LiveDNA: 20.16146
Y.Y. El Seady
Department of Animal Physiology, Faculty of Veterinary Medicine, Mansoura University, Egypt
S.M. Nasr
Department of Animal Husbandry and Wealth Development, Faculty of Veterinary Medicine, Damanhur University, Egypt
LiveDNA: 20.12509
N.A. El Kholy
Researcher at Central Diagnostic and Research Laboratory, Faculty of Veterinary Medicine, Kafr Elsheikh University, Egypt
Asian Journal of Animal and Veterinary Advances
Year: 2016 | Volume: 11 | Issue: 9 | Page No.: 548-555







ABSTRACT
Objective: The effect of breed, apoVLDL-II gene polymorphism and metabolic biochemical markers on growth and body composition traits were investigated in 400 different four chicken breeds (Cobb, Hubbard, Aves and Ross). Methodology: Blood samples were collected from chickens of each breed for DNA extraction and biochemical assay. Association analysis of the impact of breed revealed that Ross breed has the highest measurements for growth and body composition traits except for feed intake; where highest feed intake was recorded for Aves one. Results: The PCR-BfmI digestion of 492 bp of a fragment of intron-I of apoVLDL-II gene revealed two fragments (396 and 96 bp) for genotype BB, three fragments (492, 396 and 96 bp) for genotype AB and undigested fragment (492 bp) for genotype AA. Statistical analysis indicated that there was significant association between apoVLDL-II genotypes and growth and body composition traits. The AB genotype was higher than both BB and AA genotypes in all growth and body composition traits except for feed intake. In concern to metabolic biochemical data, broiler breeds could be significantly (p<0.05) affect serum levels of T3 and T4 to reach the peak value in Ross and minimal value in Aves breed. These data could be compatible with growth and body composition data. Meanwhile, the level of plasma proteins or lipid profile do not showed a significant different between breeds. Conclusion: This study highlights the possibility of breed effect, apoVLDL-II/BfmI locus and metabolic biochemical markers as candidates for growth and body composition traits in chickens.
PDF Abstract XML References Citation
How to cite this article
H.M. Ghanem, A.I. Ateya, Y.Y. El Seady, S.M. Nasr and N.A. El Kholy, 2016. Effect of Breed, ApoVLDL-II Gene Polymorphism and Metabolic Biochemical Markers on Growth and Body Composition Traits in Commercial Broiler Breeds. Asian Journal of Animal and Veterinary Advances, 11: 548-555.
DOI: 10.3923/ajava.2016.548.555
URL: https://scialert.net/abstract/?doi=ajava.2016.548.555
DOI: 10.3923/ajava.2016.548.555
URL: https://scialert.net/abstract/?doi=ajava.2016.548.555
INTRODUCTION
Poultry represent nearly one-fourth of all the meat produced globally. It is a source of protein that plays an important role in human nutrition. Modern intensive production strategies can produce market ready broiler chickens in less than 6 weeks. This achievement arises from improved productivity via genetic selection, improved feeding and health management practices1. It has been estimated that 90% of the phenotypic changes in poultry have come from genetic progress2. So the main objective of primary breeders was to select breed that deliver the best commercial performance (growth rate, feed conversion and meat yield) with high economic return for their customers3. Genetic diversity in local or domestic breeds of animals not only allows breeders and researchers to develop new characteristics in response to changes in environment, diseases or market conditions and maintain genetic diversity but also improves productivity. The yields of animals are the result of the combined effects of genotype and environmental conditions. In order to increase the yield level, it is necessary to optimize the environmental conditions and to improve the genetic structure of the animal by selection and crossbreeding4.
Identifying the quantitative trait loci responsible for the economic important traits in chickens and understanding the genetic and metabolic control of growth will provide an opportunity for genetic improvement and facilitate poultry breeding programs. The application of genetic selection methods in the poultry industry has resulted in increased growth rate and carcass quality5. The restriction enzymes cleaved the DNA molecule at specific recognition sites, originating a set of fragments with different length that could be separated according to their molecular size by conventional gel electrophoresis6.
Very low density apolipoprotein-II (apoVLDL-II) is a major constituent of the very low density lipoprotein fraction of hen serum. It is a small phospholipid-binding protein; synthesized in the liver7,8. The apoVLDL-II protein is also present and detectable in the plasma and liver of normal young cockerels9,10. The function of apoVLDL-II is the lipid transportation (triacylglycerol) in the form of very low density lipoprotein in the plasma. This protein binds to phospholipid and forms an outer polar shell surrounding the water-insoluble lipid core11. Therefore, apoVLDL-II and its gene could be used as candidates for growth and body composition traits. Association between apoVLDL-II gene polymorphism and growth and body composition traits in the chicken is scare12,13.
Hormonal control of avian growth and development has been nearly exclusively studied in poultry species14. In vertebrates, including birds and Thyroid Hormones (TH) are essential for the control of many fundamental physiological processes, such as development, differentiation, growth, metabolism and thermoregulation15. Moreover, the physiological bases of the improvement in body weight and growth in commercial broilers made by genetic selection16. It was proved that, the absolute plasma levels of thyroid hormones could be varies among avian species throughout their age14. Multiple normal physiologic processes in the bird (e.g., reproductive cycle) as well as species can influence plasma lipid profile concentrations17. However, comparative studies could not detect variability in the lipid profile of avian breeds18.
The objectives of the present study were to reveal the effect of breed, BfmI polymorphism in the intron-I of apoVLDL-II gene using PCR-RFLP and metabolic biochemical markers (thyroid hormones, total protein, albumin and lipid profile) on growth and body composition traits in four different commercial broiler breeds (Cobb, Hubbard, Aves and Ross).
MATERIALS AND METHODS
Experimental birds and management: In this study, a total of 400 one-day-old different four chicken breeds (Cobb, Hubbard, Aves and Ross) 100 birds each were used. The experimental birds were brooded together in the brooding unit (deep litter system) under the same environmental conditions as one group for 6 days for adaptation using 100 W electric bulb, 30°C brooding temperature and 14 h of light per day. At 7 days of age, each group was randomly allotted to three subgroups of birds each as a replicates to begin the experiment, the feeding trial lasted for 42 days. Good ventilation and fresh air were provided to reduce ammonia concentration in the house. Chickens were fed a concentrate diet formulated according to NRC19 (Table 1) and clean fresh water was available all times. Vitamin E and selenium, AD3E, prophylactic antibiotics and anti-coccidian drugs were also provided. This study protocol was approved by the committee of animal welfare and ethics, Faculty of Veterinary Medicine, Mansoura University.
Data measurements: Body weight was recorded at the beginning of the experiment. Chicks were weighted in groups (each replicate individually) to the nearest gram (g), then weekly till the end of the experiment using digital balance (Sartorius). Weekly weight gain was calculated by subtracting the body weight between two successive weights using the following equation:
BWG = Bw2-Bw1
where, Bw1 is a previous week’s body weight and Bw2 is a former body weight.
| Table 1: | Composition of diet fed to chickens |
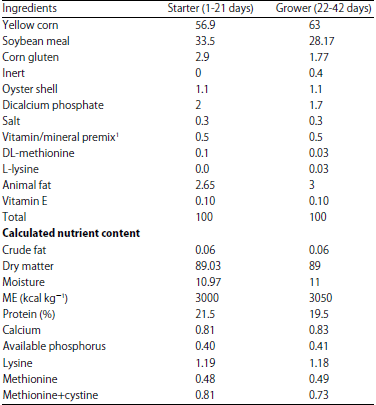 | |
For each kg of the diets, Vitamin A: 9,000,000 IU, Vitamin D3: 2,000,000 IU, Vitamin B1: 1,800 mg, Vitamin B2: 6,600 mg, Vitamin B3: 10,000 mg, Vitamin B6: 3,000 mg, Vitamin B12: 15 mg, Vitamin E: 18,000 mg, Vitamin K3: 2,000 mg, Vitamin B9: 1,000 mg, Vitamin B5: 30,000 mg, Folic acid: 21 mg, Nicotinic acid: 65 mg, Biotin: 14 mg, Choline chloride: 500,000 mg, Mn: 100,000 mg, Zn: 85,000 mg, Fe: 50,000 mg, Cu: 10,000 mg, I: 1,000 mg and Se: 200 mg | |
Feed efficiency was calculated from each replicate as gram body weight gain per gram feed consumed. The birds were fastening for 10 h before slaughter20. Then each replicate were individually weighed to the nearest gram and slaughtered. Following a 4 min bleeding time, each bird was dipped in a water bath for 2 min and feathers were removed by hand. Then the weights of hot carcass, dressing percentage and giblets (gizzard, liver and heart) were recorded.
Experimental samples: Blood samples were collected from chickens of each breed into tubes containing an anticoagulant disodium EDTA for DNA extraction. Also, at end of experiment (42 days) and after slaughtering blood samples were collected in a clean tube, left to coagulate and collect serum that stored frozen until biochemical assay.
PCR-RFLP for apoVLDL-II gene: The PCR was done for amplification of a fragment of intron-I of apoVLDL-II gene with expected amplicon size of 492 bp using the following primers12:
| • | Forward: 5‘-CCT CTA TGA CAT GGT TGC CT-3‘ |
| • | Reverse: 5‘-ATG GGT TTG ACC CTG CTA TG-3‘ |
The polymerase chain reaction mixture was done in a 25 μL consisted of: 3 μL DNA, 8.5 μL H2O (d.d water), 12.5 μL PCR master mix (Jena Bioscience, Germany), 0.5 μL of each primer. The final reaction mixture was achieved in a thermal cycler and the PCR temperature schedule program was carried out by an initial denaturation at 94°C for 4 min followed by 34 cycles of 94°C for 1 min for denaturation, primer hybridization temperature at 58°C for 1 min, primer extension at 72°C for 1 min and the final elongation at 72°C lasts for 10 min.
The amplified DNA fragments of apoVLDL-II gene were digested with fast digest BfmI (Thermo scientific, #FD1164) (the same action of SfcI) at 37°C for 5 min. The reaction volume was d one in 30 μL consisted of: 10 μL PCR product, 17 μL H2O (d.d water), 2 μL 10x fast digest green buffer and 1 μL restriction enzyme. The cleaved fragments were detected by agarose gel electrophoresis then the fragment patterns were visualized under UV using gel documentation system.
Biochemical data
Thyroid hormones: Serum levels of thyroid hormones (T3 and T4) were determined by ELIZA methods using commercial kits supplied (Pishtaz Teb Diagnostic, Germany)21.
Total protein and albumin: Total Protein (TPs) and albumin were measured in the serum by UV-calorimetric spectrophotometric method using commercial kits (Vitro Scient, Egypt)22.
Lipid profile: Lipid profile including Total Cholesterol (TC), triglycerides (TGs), High Density Lipoprotein (HDL) and Low Density Lipoprotein (LDL) were determined using a commercial kits supplied (Stan Bio., USA)23.
Statistical analysis
Association analysis: Preliminary test of percentage data revealed that they were homogeneous with no need for transformation. To test association among apoVLDL-II gene, growth and body composition, General Linear Model procedure (PROC GLM) of Statistical Analysis System package (SAS version 9.2, SAS Institute, Inc., Cary, NC.24 and Cilek25) was used implementing the following model:
Yijk = μ+Gi+Bj+eijk
where, Yijk is the phenotypic value of the trait, μ is overall population mean, Gi is fixed effect of apoVLDL-II genotype, Bj is fixed effect of breed and eijk is random error effect. Due to the non-significant effect of both replicate and (genotype×breed) interaction, they were left out of the full model.
Gene and genotypic frequencies in apoVLDL-II locus: Gene and genotypic frequencies were estimated by simple counting of the alleles based on the electrophoresis results26,27.
RESULTS
Effect of breed: Association analysis revealed that significant effect of chicken breed on growth and body composition traits. Ross breed has the highest measurements of live body weight, weight gain, feed efficiency, dressing percentage and giblets weight followed by Cobb-500, Hubbard and Aves breeds, respectively. Meanwhile, the highest feed intake was recorded for Aves breed (Table 2).
Effect of apoVLDL-II gene polymorphism: One pair of specific primers was used to amplify specific DNA fragments 492 bp of intron-I of apoVLDL-II gene (Fig. 1). Restriction analysis of 492 bp PCR products digested with BfmI revealed that, two fragments (396 and 96 bp) for genotype BB, three fragments (492, 396 and 96 bp) for genotype AB and undigested fragment (492 bp) for genotype AA (Fig. 2).
| Table 2: | Effect of breed on growth and body composition traits (LSM±SEM) |
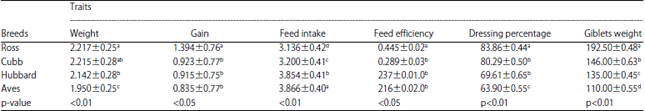 | |
| Means of different levels within the same column having different superscripts are significantly different | |
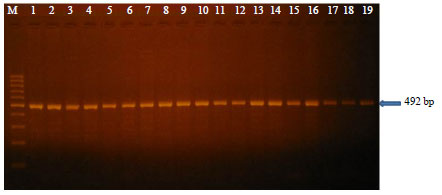 | |
| Fig. 1: | Representative PCR results of apoVLDL-II gene, Lane M: DNA marker and Lanes 1-19: 492 bp amplified fragment of apoVLDL-II gene |
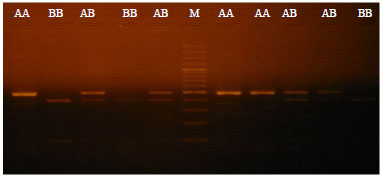 | |
| Fig. 2: | Representative BfmI restriction fragment pattern of apoVLDL-II gene (492 bp), BB: Restriction fragment of 396 and 96 bp, AB: Restriction fragment of 492, 396 and 96 bp, AA: Restriction fragment of 492 bp and M: 100 bp molecular size marker |
| Table 3: | Frequency of genotypes and alleles in the apoVLDL-II locus |
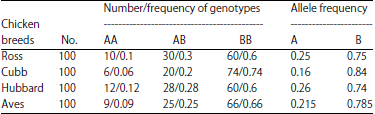 | |
| Table 4: | Associations of apoVLDL-II genotypes with growth and body composition (LSM±SEM) |
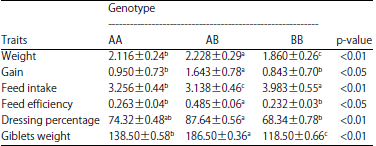 | |
| Means of different levels within the same column having different superscripts are significantly different | |
Using PCR-RFLP method, the incidence of apoVLDL-II genotypes and frequencies of alleles were calculated. The B allele was more frequent than A allele in the four breeds (Table 3). Statistical analysis indicated that there was significant association between apoVLDL-II genotypes and growth and body composition traits. The AB genotype was higher in all growth and body composition traits except for feed intake than both BB and AA genotypes (Table 4).
Effect of metabolic biochemical markers: In this comparative study, serum levels of T3 and T4 were determined in different four breeds of chickens with different growth rate and intensity. It was noticed that broiler breeds could be significantly (p<0.05) affect serum levels of T3 and T4 to reach the peak value in Ross and minimal value in Aves breed. These data could be compatible with growth and body composition data. Meanwhile, the level of plasma proteins or lipid profile do not showed a significant different between breeds (Fig. 3).
DISCUSSION
Improving economic traits in chicken has become of the major hurdle that can be achieved by identification and utilization of QTLs that provide the potential for genetic improvement in selection programs without slaughtering. Recent advances in molecular genetics and development of molecular techniques have led to the discovery of genes or markers associated with genes involved in growth and body composition traits28. When apolymorphic locus is associated with the economic traits of animals, therefore that polymorphic locus can be used as genetic markers for the trait and becomes useful as a selection criterion for genetic improvement of such trait (marker assisted selection).
In this study, Ross breed has the highest measurements of live body weight, weight gain, feed efficiency, dressing percentage and giblets weight than other breeds. Meanwhile, the highest feed intake was recorded for Aves breed. These findings emphasize that the chicken breed could be significantly affect growth and body composition traits. These results are consistent with the findings of several studies29-33. The researchers proven that the breed has significant (p<0.05) effect on live weight at 8 weeks. In the same respect, it was cited that Cobb-500 breed achieved higher body weight and weight gain than the other breeds34-39. However, it was found that no significant effect of breed on live body weight40. In regard to carcass traits, significant effect of breed in all the carcass traits evaluated including dressing percentage and giblets weight was reported32,41,42.
In this study, apoVLDL-II gene polymorphism and its effect on growth and body composition traits in the four chicken breeds revealed that, a specific PCR product of a desirable size (492 bp) involving intron-I of apoVLDL-II gene. The following DNA restriction fragments were obtained for apoVLDL-II-BfmI digestion: Digested (396 and 96 bp) for genotype BB, three fragments (492, 396 and 96 bp) for genotype AB and undigested fragment (249 bp) for genotype AA with the B allele more frequent than A allele in the four breeds. Statistical analysis indicated that there was significant association between apoVLDL-II genotypes and growth and body composition traits. The AB genotype was higher in all growth and body composition traits except for feed intake than both BB and AA genotypes. The association of apoVLDL-II gene polymorphism on chicken growth and body composition traits in Iranian commercial broiler line was investigated3. According to the results, genotyping the apoVLDL-II gene was carried out using PCR-RFLP method. The apoVLDL-II SfcI digestion showed a mutation in 492 bp fragment. The obtained DNA restriction fragments were similar to that denoted in this study but AA genotype was not considered at analysis due to its very low frequency. Th apoVLDL-II gene polymorphism was significantly (p<0.05) associated with body weight at 6 week (BW6), Carcass Weight (CW), Breast Muscle Weight (BMW), Drumstick Weight (DW) and wing weight (WINW). No significant difference was observed in the back weight (BAKW) and Abdominal Fat Weight (AFW).
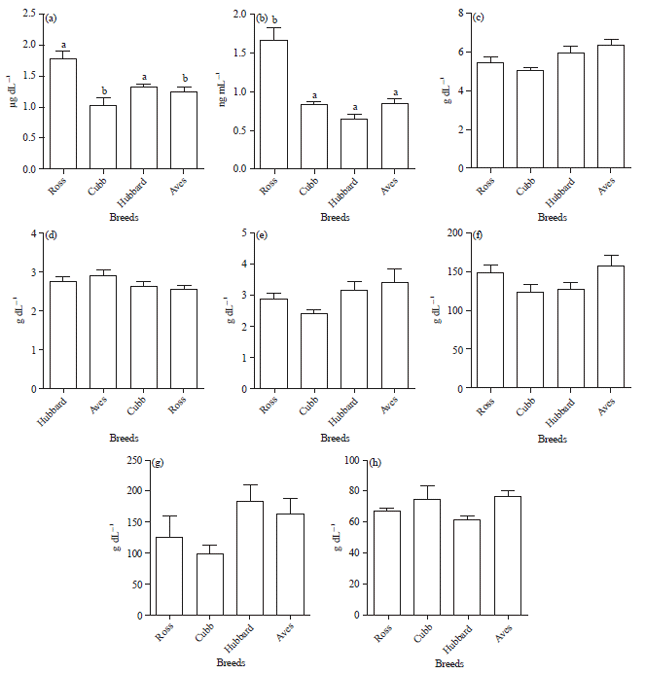 | |
| Fig. 3(a-h): | Serum level of metabolic biochemical markers (a) T4, (b) T3 thyroid hormones, (c) Total protein, (d) Albumin, (e) Globulin, (f) Cholesterol, (g) Triglycerides and (h) High density lipoprotein in the four different chicken breeds |
Detection of apoVLDL-II gene polymorphisms and evaluation the associations between apoVLDL-II SNP and growth and body composition traits in chickens was also identified12. The researchers established by crossing broiler sires with dams from 2 unrelated highly inbred lines (Leghorn and Fayoumi). The restriction enzyme SfcI digested PCR products had fragment sizes of 396 and 96 bp for the broiler line and 492 bp for the Fayoumi line. The apoVLDL-II polymorphism between broiler and Fayoumi was associated with multiple traits of growth and body composition in the 148 male F2 individuals, including BW, breast muscle weight, drumstick weight and tibia length.
Another study on apoVLDL-II gene polymorphism and growth traits, the association of avian apoVLDL-II gene polymorphism with body weight and fat was investigated13. Polymorphism was detected by PCR-RFLP and PCR-SSCP techniques. Polymorphism in apoVLDL-II gene was significantly (p<0.05) associated with body weight and fat weight at VLDL9 and VLDL17 loci in lean chicken. In addition, polymorphism of apoVLDL-II gene at VLDL6, VLDL9 and VLDL10 loci were significantly (p<0.05) associated with body weight and fat weight.
In this study, the broiler breeds could be significantly (p<0.05) affect serum levels of T3 and T4 to reach the peak value in Ross and minimal value in Aves breed. These data could be in accordance with growth and body composition phenotypic measurements. It was approved that concentrations of T3 appeared to be high when growth rate improved15,16. These data could be in agreement of the data obtained in the present study where thyroid hormones levels have positive relation with the broiler breeds of higher growth rate. Likewise, sex linked dwarf chickens have a very low T3 concentration that may reflect the essentiality of thyroid gland for normal growth43.
In this study, the level of plasma proteins or lipid profile do not showed a significant different between breeds. Multiple normal physiologic processes in the bird (e.g., reproductive cycle), as well as species can influence plasma cholesterol concentrations17. This could be in agreement with the results denoted in this study. Moreover, variability in the lipid profile of avian breeds could not be detected18. However, in this case, a breeds difference in this analyze was not found, suggesting that cholesterol concentrations may be relatively insensitive indicators of physiologic or breeds in bird. Similar to findings in humans, the data of present study suggest that increases in body weight and BCS are linked to increased plasma lipid. However, at this time based on the preliminary findings, it was suggested that, plasma concentrations of cholesterol, triglyceride and HDL alone should not be used as a sole indicator of growth or body weight but certainly biochemical data deserves further investigation in this and other avian species commonly breeds for fattening purposes.
CONCLUSION
This study supports the plausible effect of breed, apoVLDL-II gene polymorphisms and the metabolic biochemical markers (thyroid hormones, total protein, albumin and lipid profile) as candidates for growth and body composition traits in chickens. Moreover, the effectiveness of RFLP as a molecular genetic marker may be contains great genetic potential resource to improve such traits results in an effective selection.
ACKNOWLEDGMENTS
The authors acknowledge Dr. Tarek Kamal, Dr. Aya Abd El-Latif and Dr. Walaa Hamada, Biotechnology Laboratory, Faculty of Veterinary Medicine, Kafr Elsheikh University for their valuable advices.
REFERENCES
- Apata, D.F., 2012. The emergence of antibiotics resistance and utilization of probiotics for poultry production. Sci. J. Microbiol.
Direct Link - Havenstein, G.B., P.R. Ferket, S.E. Scheideler and B.T. Larson, 1994. Growth, livability and feed conversion of 1957 vs 1991 broilers when fed typical 1957 and 1991 broiler diets. Poult. Sci., 73: 1785-1794.
CrossRefPubMedDirect Link - Seyedabadi, R.H., C. Amirinia, N. Mirmozafari, R.V. Torshizi and M. Chamani, 2010. Association between single nucleotide polymorphism of apoVLDL-II gene with growth and body composition traits in Iranian commercial broiler line. Afr. J. Biotechnol., 9: 4175-4178.
Direct Link - Cilek, S. and M.E. Tekin, 2005. Environmental factors affecting milk yield and fertility traits of Simmental cows raised at the Kazova State Farm and phenotypic correlations between these traits. Turk. J. Vet. Anim. Sci., 29: 987-993.
Direct Link - Zhou, H., A.D. Mitchell, J.P. McMurtry, C.M. Ashwell and S.J. Lamont, 2005. Insulin-like growth factor-I gene polymorphism associations with growth, body composition, skeleton integrity and metabolic traits in chickens. Poult. Sci., 84: 212-219.
PubMedDirect Link - Pereira, F., J. Carneiro and A. Amorim, 2008. Identification of species with DNA-based technology: Current progress and challenges. Recent Pat. DNA. Gene Seq., 2: 187-200.
PubMedDirect Link - Luskey, K.L., M.S. Brown and J.L. Goldstein, 1974. Stimulation of the synthesis of very low density lipoproteins in rooster liver by estradiol. J. Biol. Chem., 249: 5939-5947.
PubMedDirect Link - Chan, L., R.L. Jackson, B.W. O'Malley and A.R. Means, 1976. Synthesis of very low density lipoproteins in the cockerel. Effects of estrogen. J. Clin. Invest., 58: 368-379.
CrossRefDirect Link - Lin, C.T. and L. Chan, 1980. Effects of estrogen on specific protein synthesis in the cockerel liver: An immunocytochemical study on major apoproteins in very low density and high density lipoproteins and albumin. Endocrinology, 107: 70-75.
CrossRefDirect Link - Blue, M.L. and D.L. Williams, 1981. Induction of avian serum apolipoprotein II and vitellogenin by tamoxifen. Biochem. Biophys. Res. Commun., 98: 785-791.
CrossRefDirect Link - Chan, L., 1983. Hormonal control of apolipoprotein synthesis. Annu. Rev. Physiol., 45: 615-623.
CrossRefDirect Link - Li, H., N. Deeb, H. Zhou, C.M. Ashwell and S.J. Lamont, 2005. Chicken quantitative trait loci for growth and body composition associated with the very low density apolipoprotein-II gene. Poult. Sci., 84: 697-703.
CrossRefPubMedDirect Link - Musa, H.H. and G.H. Chen, 2007. Association of polymorphisms in avian apoVLDL-II gene with body weight and abdominal fat weight. Afr. J. Biotechnol., 6: 2009-2013.
Direct Link - Vyboh, P., M. Zeman, M. Jurani, J. Buyse and E. Decuypere, 1996. Plasma thyroid hormone and growth hormone patterns in precocial Japanese quail and altricial European starlings during postnatal development. Comp. Biochem. Physiol. Part C: Pharmacol. Toxicol. Endocrinol., 114: 23-27.
CrossRefDirect Link - Lauterio, T.J., E. Decuypere and C.G. Scanes, 1986. Growth, protein synthesis and plasma concentrations of growth hormone, thyroxine and triiodothyronine in dwarf, control and growth-selected strains of broiler-type domestic fowl. Comp. Biochem. Physiol. Part A: Physiol., 83: 621-632.
CrossRefDirect Link - Belcher, C., J. Heatley, C. Petzinger, S. Hoppes and C.D. Larner et al., 2014. Evaluation of plasma cholesterol, triglyceride and lipid density profiles in captive monk parakeets (Myiopsitta monachus). J. Exotic Pet Med., 23: 71-78.
CrossRefDirect Link - NRC., 1994. Nutrient Requirements of Poultry. 9th Edn., National Academy Press, Washington, DC., USA., ISBN-13: 9780309048927, Pages: 176.
Direct Link - Young, D.S., L.C. Pestaner and V. Gibberman, 1975. Effects of drugs on clinical laboratory tests. Clin. Chem., 21: 1D-432D.
PubMedDirect Link - Fossati, P. and L. Prencipe, 1982. Serum triglycerides determined colorimetrically with an enzyme that produces hydrogen peroxide. Clin. Chem., 28: 2077-2080.
CrossRefPubMedDirect Link - Cilek, S., 2009. Reproductive traits of holstein cows raised at Polatli State farm in Turkey. J. Anim. Vet. Adv., 8: 1-5.
Direct Link - Falconer, D.S. and T.F.C. Mackay, 1996. Introduction to Quantitative Genetics. 4th Edn., Prentice Hall, Harlow, England, ISBN-13: 9780582243026, Pages: 464.
Direct Link - Kaygisiz, A., C. Bengi and S. Cilek, 2011. Investigation of leptin gene polymorphisms in East Anatolian red Anatolian and black cattle and determination of genetic distance from brown Swiss cattle. J. Anim. Plant Sci., 21: 121-125.
Direct Link - Gao, Y., R. Zhang, X. Hu and N. Li, 2007. Application of genomic technologies to the improvement of meat quality of farm animals. Meat. Sci., 77: 36-45.
CrossRefDirect Link - Orr, H.L., C. Hunt and C.J. Randall, 1984. Yield of carcass, parts, meat, skin and bone of eight strains of broilers. Poult. Sci., 63: 2197-2200.
CrossRefDirect Link - Olawumi, S.O. and S.S. Fagbuaro, 2011. Productive performance of three commercial broiler genotypes reared in the derived Savannah zone of Nigeria. Int. J. Agric. Res., 6: 798-804.
CrossRefDirect Link - Shahin, K.A. and F. Abd Elazeem, 2005. Effects of breed, sex and diet and their interactions on carcass composition and tissue weight distribution of broiler chickens. Archiv Tierzucht, 48: 612-626.
Direct Link - Ojedapo, L.O., O. Akinokun, T.A. Adedeji, T.B. Olayeni, S.A. Ameen and S.R. Amao, 2008. Effect of strain and sex on carcass characteristics of three commercial broilers reared in deep litter system in the derived Savannah area of Nigeria. World J. Agric. Sci., 4: 487-491.
Direct Link - Razuki, W.M., S.A. Mukhlis, F.H. Jasim and R.F. Hamad, 2011. Productive performance of four commercial broiler genotypes reared under high ambient temperatures. Int. J. Poult. Sci., 10: 87-92.
CrossRefDirect Link - Gonzales, E., J. Buyse, M.M. Lodi, T.S. Takita, N. Bugs and E. Decuypere, 1998. Performance, incidence of metabolic disturbances and endocrine variables of food-restricted male broiler chickens. Br. Poult. Sci., 39: 671-678.
CrossRefDirect Link - Sarker, M.S.K., S.U. Ahmed, S.D. Chowdhury, M.A. Hamid and M.M. Rahman, 2001. Performance of different fast growing broiler strains in winter. Pak. J. Biol. Sci., 4: 251-254.
CrossRefDirect Link - Sarker, M.S.K., M.A. Islam, S.U. Ahmed and J. Alam, 2002. Profitability and meat yield traits of different fast growing broiler strains in winter. J. Biol. Sci., 2: 361-363.
CrossRefDirect Link - Abdullah, A.Y., N.A. Al-Beitawi, M.M.S. Rjoup, R.I. Qudsieh and M.A.A. Ishmais, 2010. Growth performance, carcass and meat quality characteristics of different commercial crosses of broiler strains of chicken. J. Poult. Sci., 47: 13-21.
CrossRefDirect Link - Hassanien, H.H.M., 2011. Productive performance of broiler chickens as affected by feed restriction systems. Asian J. Poult. Sci., 5: 21-27.
CrossRefDirect Link - Ghanem, H.M., 2014. Impact of breed and feed restriction on some productive and carcass traits in broiler chickens. Int. J. Sci. Res., 3: 2745-2751.
Direct Link - Becker, W.A., J.V. Spencer, L.W. Mirosh and J.A. Verstrate, 1981. Abdominal and carcass fat in five broiler strains. Poult. Sci., 60: 693-697.
CrossRefPubMedDirect Link - Musa, H.H., G.H. Chen, J.H. Cheng, B.C. Li and D.M. Mekki, 2006. Study on carcass characteristics of chicken breeds raised under the intensive condition. Int. J. Poult. Sci., 5: 530-533.
CrossRefDirect Link - Taha, A.E., F.A. Abd El-Ghany and M.M. Sharaf, 2011. Strain and sex effects on productive and slaughter performance of local Egyptian and Canadian chicken strains. J. World's Poult. Res., 1: 11-17.
Direct Link - Tixier-Boichard, M., L.M. Huybrechts, E. Kuhn, E. Decuypere, J. Charrier and P. Mongin, 1989. Physiological studies on the sex-linked dwarfism of the fowl: A review on the search for the gene's primary effect. Genet. Select. Evol., 21: 217-234.
CrossRefDirect Link