
A.E. Abdel-Khalek
Department of Animal Production, Faculty of Agriculture, Mansoura University, Mansoura, Egypt
I.T. El-Ratel
Animal Production Research Institute, Agricultural Research Center, Egypt
W.M. Wafa
Animal Production Research Institute, Agricultural Research Center, Egypt
H.A. El-Nagar
Animal Production Research Institute, Agricultural Research Center, Egypt
G.E. Younan
Animal Production Research Institute, Agricultural Research Center, Egypt
Sara F. Fouda
Department of Poultry Production, Faculty of Agriculture, Mansoura University, Mansoura, Egypt
Asian Journal of Animal and Veterinary Advances
Year: 2016 | Volume: 11 | Issue: 1 | Page No.: 53-59







ABSTRACT
To study ovulatory response and genital and in vitro embryo characteristics of control rabbit does (G1) compared with those treated with Coenzyme Q10 (10 mg CoQ10/kg LBW, G2) and L-Carnitine (40 mg LC kg–1 LBW, G3). Total of 15 mature NZW rabbit does (5.3 mo of age, 3.35 kg LBW) and 6 NZW bucks (7.0-9.0 mo of age and 3.85 kg LBW) were used in this study for 21 days prior to natural mating. All does were slaughtered 64-66 h post-mating to determine genital measurements and follicular and Corpora Lutea (CLs) number, Ovulation Rate (OR) and embryo Recovery Rate (RR) rates. By flushing the oviducts, embryos was counted and evaluated. Also, post-thawing Survival Rate (SR) and Viability Rate (VR) rates were determined. Results showed that dose in G2 showed greater number of follicles (18.2), CLs (13.6) and embryos (16.3) per dose and the highest OR (74.7%), RR (100%), excellent embryos (86.8%) and the lowest good and fair embryos (10.4% for each) compared with G1. Genital characteristics were not affected. Post-thawing SR was not affected, while VR was higher (p<0.05) in G2 and G3 than in G1 (88.9 and 87.5 vs. 76.9%). In vitro blastocyst production rate was higher (p<0.05) in G2 and G3 than in G1 (64.58 and 60.00 vs. 52.63%). In conclusion, pre-conception daily oral dose of rabbit does for 21 days with 10 mg CoQ10/kg LBW has impact on yield, quality, survival and viability of vitrified embryos and in vitro blastocyst production.
PDF Abstract XML References Citation
How to cite this article
A.E. Abdel-Khalek, I.T. El-Ratel, W.M. Wafa, H.A. El-Nagar, G.E. Younan and Sara F. Fouda, 2016. Effect of Pre-Conception Coenzyme Q10 and L-Carnitine Treatments on Ovulatory Response, Genital Characteristics and in vitro Embryo Characteristics in Rabbits. Asian Journal of Animal and Veterinary Advances, 11: 53-59.
DOI: 10.3923/ajava.2016.53.59
URL: https://scialert.net/abstract/?doi=ajava.2016.53.59
DOI: 10.3923/ajava.2016.53.59
URL: https://scialert.net/abstract/?doi=ajava.2016.53.59
INTRODUCTION
In vivo embryos rely on mitochondrial oxidative phosphorylation for energy, a process which is subsequently accompanied by ROS generation (Du Plessis et al., 2008). In order to optimize gamete/embryo quality and improve Assisted Reproductive Technology (ART) outcome, deliberate preventive measures are necessary to reduce any incidental build-up of Reactive Oxygen Species (ROS) leading to oxidative stress development during ART. The fast developing embryo produces energy via ATP generation through mitochondrial oxidative phosphorylation and glycolysis (Agarwal et al., 2014). As it develops, the embryo is capable of producing ROS through several pathways, namely oxidative phosphorylation, NADPH and xanthine oxidase systems (Guerin et al., 2001). The ROS generation is particularly excessive during embryonic genome activation, embryonic compaction and hatching as these processes demand greater amounts of energy (Gott et al., 1990).
One method to achieve this would be by enhancing the antioxidant capacity of the gamete and embryo against the harmful assault of oxidation. Mitochondrial dysfunction may lead to incomplete detoxification of the free radicals, which may lead to oxidative damage to macromolecules such as lipids, proteins and DNA (Abdelrazik et al., 2009).
L-Carnitine (LC) is a small water-soluble molecule which plays an important role in fat metabolism (Abdelrazik et al., 2009). It is essential for the normal mitochondrial oxidation of fatty acids and excretion of acyl-coenzyme A (acyl-CoA) esters and affects the adenosine triphosphate (ATP) levels (Vanella et al., 2000). The LC protects cell membrane and DNA against damage (Ye et al., 2010) by reducing the accumulation of Reactive Oxygen Species (ROS), enhancing the activity of numerous antioxidant enzymes, e.g., superoxide dismutase, catalase and glutathione peroxidase (Rizzo et al., 2010) and has a pivotal role in mitochondrial oxidation of long-chain fatty acids which increase energy supply to the cell (Zhou et al., 2007). The unique dual effects of LC in terms of reducing cellular lipid content and providing antioxidative protection make it a novel candidate reagent for the non-invasive improvement of cryotolerance and developmental competence in embryos of farm animals (Takahashi et al., 2013).
Coenzyme Q10 (CoQ10) is a fat soluble vitamin-like substance present in every cell of the body and serves as a coenzyme for several of the key enzymatic steps of in the production of energy within the cell (Kapoor and Kapoor, 2013). In addition to playing a key role in the mitochondrial electron transport chain, it is a critical coenzyme in the synthesis of adenosine triphosphate (Abdulhasan et al., 2015). Also, CoQ10 is antioxidant that has great importance against free radicals, with its tissue concentration being 5-10-fold higher than the other main lipid soluble antioxidant (Bentinger et al., 2007) as well as it protects the stability of the cell membrane, DNA from free radicals induced oxidative damage and helps recycling of vitamin E and maintain healthy energy levels (El-Tohamy et al., 2012).
Dietary supplementation with CoQ10 may increase mitochondrial activity within the oocyte and developing embryo (Bentov et al., 2010). Since tissue levels of CoQ10 decrease with age, supplementation of this agent may improve mitochondrial function in the ovary and may improve embryo quality, especially in women of advanced reproductive age (Chappel, 2013). Several authors demonstrated that in vitro LC treatment can improve embryo development, however, the available information about whether circulating carnitine or in vivo supplementation with LC (Eder, 2009) or CoQ10 can improve embryo quality are scar.
Therefore, the objective of this study was to evaluate the effect of pre-conception Coenzyme Q10 (10 mg kg–1 LBW) and L-Carnitine (40 mg kg–1 LBW) administration on ovulatory response, genital characteristics and yield, quality, vitrification and developmental competence of rabbit embryos.
MATERIALS AND METHODS
This study was carried out at the International Livestock Management Training Center (ILMTC), belonging to the Animal Production Research Institute, Agricultural Research Center, Ministry of Agriculture.
Animals: Total of 15 New Zealand White (NZW) mature rabbit does (5.3 mo of age and 3.35 kg live body weight) as embryo donors as well as 6 NZW bucks aging 7.0-9.0 mo and weighing 3.85 kg for natural mating were used in this study during the experimental period.
Experimental groups: Rabbit does were divided randomly into 3 groups (n = 5) per group. The 1st group (G1) was considered as a control group without treatment (G1), while does in the 2nd and 3rd groups were given 10 mg Coenzyme Q10 (CoQ10)/kg LBW and 40 mg L-Carnitine (LC)/kg LBW as daily oral doses for 21 days as a treatment period prior to mating.
Experimental procedures: In this study, all does from each group were slaughtered 64-66 h post mating and immediately after slaughtering ovaries and all reproductive tract of each dose were removed, washed by distilled water and dried by cleaning paper. Ovaries and the reproductive tract were excised, submerged in a flacon plastic tissue culture dishes (60×15 mm) containing saline solution at 38.5°C. Embryos were recovered by flushing from reproductive tract. Number of visible follicles (more than 1 mm in diameter) and Corpora Lutea (CLs) on the ovarian surface was recorded for each dose, then Ovulation Rate (OR) and Embryo Recovery Rate (ERR) were calculated as the following:
 |
In addition, measurements (weight, length and width) of the reproductive tract segments (oviducts and uterine horn) were determined.
Embryo recovery: After 64-66 h of mating, embryos were recovered from oviduct by flushing using Phosphate Buffer Solution (PBS) supplemented with 0.2% (w/v) of Bovine Serum Albumin (BSA) and antibiotics (100 IU penicillin and 100 μg mL–1 streptomycin (Sigma Chemical Co., St. Louis, Mo, USA) in Petri dishes. Embryos were washed three times with PBS, counted and evaluated morphologically for quality under inverted microscope to excellent, good, fair and poor embryos. All recovered embryos at morula stage were cryopreserved by vitrification method (straws).
Embryo vitrification: A Basal Medium (BM) containing PBS supplemented with 20% Fetal Calf Serum (FCS) and antibiotics (100 IU mL–1 penicillin and 100 μg mL–1 streptomycin) was used for vitrification. Vitrification was carried out in two steps: in the 1st step, embryos were pipetted into Vitrification Solution 1 (VS1) containing BM+12.5% (v:v) Ethylene Glycol (EG)+12.5% (v:v) dimethyl-sulfoxide (DMSO)+0.5 M sucrose) in disposable sterile Petri dish for 2 min. In the 2nd step, the embryos were pipette into VS2 (BM+20% (v: v) EG+20% (v: v) DMSO+0.5 M sucrose) for 30 sec. Then eight embryos were suspended in VS2, loaded in 0.25 mL plastic straws (IMV, L’Aigle, France), sealed and plunged directly into liquid nitrogen for at least 15 days.
Thawing and evaluation of post-thawing embryos: Vitrified embryos were thawed by plunging the content of straw into 1 mL of thawing solution (PBS+4% BSA+0.33 M sucrose) for 1 min, followed by serial dilutions in PBS+0.175 M sucrose solution for 2 min to remove the intracellular cryoprotectants, then embryos were washed three times in PBS solution for 5 min per time to remove cryoprotectants at 25°C.
Survival rate was calculated based on number of post-thawing recovered embryos, while viability rate in term of normal (viable) and abnormal (unviable) number of vitrified embryos was morphologically evaluated based on abnormalities in mucin coat, intact zona pellucida, blastomeres and refractive cytoplasm.
Embryo culture in vitro: The survived viable embryos were in vitro cultured in 100 μL drops of tissue culture medium-199 (TCM 199, Sigma) supplemented with 4 mg mL–1 BSA and 50 μg mL–1 of Gentamicin sulphate under mineral oil in CO2 incubator at 38°C, 95% humidity and 5% CO2 in air to develop into the blastocyst stage. Embryos were assessed to record blastocyst and hatching blastocyst formation rates for 5 days.
Statistical analysis: Data were analyzed by analysis of variance using computer program of SAS (2000). The significant differences among group means were preformed using Duncan Range Test (Duncan, 1955).
RESULTS
Ovulatory response: Results presented in Table 1 concerning the ovulatory response of treated rabbits showed that number of visible follicles (18.2 and 15.8), corpora lutea (13.6 and 10.2) and recovered embryos (16.3 and 9.6) per dose as well as ovulation rate (74.7 and 64.6%) were significantly (p<0.05) greater in G2 and G3 than in G1 (16.0, 8.2 and 7.4 dose–1 and 51.3%).
| Table 1: | Ovarian characteristics and ovulatory response of rabbit does as affected by CoQ10 and LC treatments |
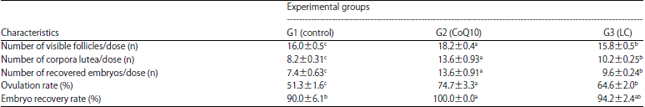 | |
a, b, cMeans within the same row with different superscripts are significantly different at p<0.05, CoQ10: Coenzyme Q10, LC: L-Carnitine | |
| Table 2: | Genital characteristic of rabbit does as affected by CoQ10 and LC treatments |
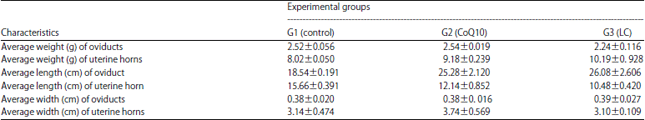 | |
| CoQ10: Coenzyme Q10, LC: L-Carnitine | |
| Table 3: | Quality of recovered fresh embryos of rabbit does as affected by CoQ10 and LC treatments |
 | |
a, bMeans within the same column with different superscripts are significantly different at p<0.05, CoQ10: Coenzyme Q10, LC: L-Carnitine | |
| Table 4: | Post-thawing survival and viability of vitrified rabbit embryos as affected by CoQ10 and LC treatments |
 | |
a, bMeans within the same column with different superscripts are significantly different at p<0.05, CoQ10: Coenzyme Q10, LC: L-Carnitine | |
However, embryo recovery rate was significantly (p<0.05) higher only in G2 than in G1 (100 vs. 90%).
Such results indicated enhancement in the ovulatory response of treated rabbits, in particular, in those treated with CoQ10 as compared to the controls.
Genital characteristics: Data in Table 2 showed insignificant differences in genital characteristics in term of averages of weight, length and width of oviducts and uterine horns was not significant. These results indicated insignificant effect of CoQ10 and LC treatments on all genital characteristics studied.
Quality of recovered embryos: Data presented in Table 3 showed was significantly (p<0.05) the highest percentage of excellent embryos and the lowest percentage of fair and poor embryos were recovered from rabbits treated with CoQ10, while those treated with LC significantly (p<0.05) showed the highest percentage of good embryos as compared to controls. The best quality of embryos was produced in G2 in term of the highest (p<0.05) percentage of excellent embryos (86.8%) and the lowest (p<0.05) good and fair embryos (10.4% for each).
Post-thawing survival and viability rates: Data in Table 4 showed insignificant differences in survival rate of vitrified embryos as affected by treatments as compared to control, ranging between 83.9 in control and 87.0 in LC group. However, post-vitrification proportion of viable embryos was significantly (p<0.05) greater (88.9 and 87.5%) and of unviable embryos was significantly (p<0.05) lower (11.1 and 12.5%) in CoQ10 and LC groups than in control group (75.9 and 23.1%), respectively.
These results indicated beneficial effect of both CoQ10 and LC treatments on viability of post-thawing viability as compared to control.
Embryo developmental competence: Results shown in Table 5 revealed significant (p<0.05) differences in percentages of embryos reaching expanded and hatching stages post-vitrification as affected by CoQ10 and LC. Expansion and hatching rates were significantly (p<0.05) higher for vitrified embryos at blastocyst stages in CoQ10 and LC groups than in control one. Developmental competence in term of blastocyst production rate after in vitro culture of recovered/vitrified embryos was higher (p<0.05) in G2 and G3 than in G1 (64.58 and 60.00 vs. 52.63%).
| Table 5: | Developmental competence of vitrified rabbit embryos as affected by CoQ10 and LC treatments |
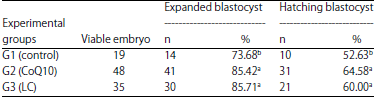 | |
a, bMeans within the same column with different superscripts are significantly different at p<0.05, CoQ10: Coenzyme Q10, LC: L-Carnitine | |
DISCUSSION
The present study aimed to study the effect of oral dose from Coenzyme Q10 (10 mg kg–1 LBW) or L-Carnitine (40 mg kg–1 LBW) for 21 days pre-mating on ovulatory response, genital characteristics and yield, quality, freezing ability by vitrification and post-thawing developmental competence of rabbit embryos. In the current study, administrating CoQ10 or LC showed pronounced effect on ovulatory response of rabbit dose by increasing number of follicles, CLs and recovered embryos, leading to marked improvement in ovulation and embryo recovery rate as compared to the control dose. It is of interest to note that impact of CoQ10 treatment was significantly (p<0.05) more than that of LC on the ovulatory response. It is of worthy noting that increasing number of ovarian structures (follicles and CLs) on the ovarian surface of does in treatment groups (G2 and G3) was associated with insignificant differences in the weight and size of the ovaries in treatment and control groups. Similar results were obtained in NZW rabbits treated with GnRH (Mehrez et al., 2014) and in superovulated California and NZW rabbits (Fahim, 2008).
In the present study, number of CLs/dose treated with CoQ10 (13.6±0.93 dose–1) was nearly similar to that recorded in NZW does treated with GnRH (13.0±247 dose–1) as reported by Mehrez et al. (2014), although greater number of embryos was recorded in the present study than that recorded by Mehrez et al. (2014), being 13.6 and 6.67 dose–1, respectively. In comparable with does treated with CoQ10 or LC, number of CLs in superovulated rabbit does ranged between 7.4-10.3 dose–1 (El-Keraby et al., 1991) or averaged 22 dose–1 (Sara, 2013) in different rabbit breeds treated with GnRH as compared to 6.6-8.0 or 15.58 dose–1 in the controls, respectively. In agreement with the obtained results, Delange (1994) found that mice given CoQ10 before ovarian stimulation made more follicles and better eggs compared to controls.
The noticed increase in number of LCs in treatment groups may be attributed to the effect of CoQ10 or LC on increasing LH surge as compared to control, reflecting higher ovulation rate. The rabbit is a reflexively ovulating species in which sensory and neuro-endocrine stimuli act together to induce a LH preovulatory surge (Dufy-Barbe et al., 1973) and determine the ovulatory response. It is of interest to note that all embryos were recovered from does treated with CoQ10 showing the impact of this treatment on embryo movement (oviduct motility) and in vivo fertilization of ovulated oocytes. In this way, dimensions (length and width) of oviducts or uterine horns were not affected by treatment with CoQ10 or LC.
In pre-implantation embryos, the quantification of amino acid and energy metabolism is emerging as a potential non-invasive tool with which to assess embryo developmental competence and even pregnancy outcome (Sturmey et al., 2008; Picton et al., 2010; Gardner et al., 2011). In vivo embryos rely on mitochondrial oxidative phosphorylation for energy, a process which is subsequently accompanied by Reactive Oxygen Species (ROS) generation (Du Plessis et al., 2008). Deliberate preventive measures are necessary to reduce any incidental build-up of ROS, leading to oxidative stress development during Assisted Reproductive Technology (ART) in order to optimize embryo quality and improve ART outcome. One method to achieve this would be by enhancing the antioxidant capacity of the embryos against the harmful assault of oxidation, because the fast developing embryo produces energy via ATP generation through mitochondrial oxidative phosphorylation and glycolysis (Agarwal et al., 2014). As it develops, the embryo is capable of producing ROS through several pathways, namely oxidative phosphorylation, NADPH and xanthine oxidase systems (Guerin et al., 2001). The ROS generation is particularly excessive during embryonic genome activation, embryonic compaction and hatching as these processes demand greater amounts of energy (Gott et al., 1990). Mechanical removal of lipid droplets (Ogawa et al., 2010) or their reduction by chemical agents (Men et al., 2006) improved tolerance of embryos to cryopreservation. According to the present results, does treated with CoQ10 yielded the highest percentage of transferable embryos in term of excellent and good embryos (92.7%) as compared to 85.4% in LC group and 75.7% in the control group. Such results indicated positive effect of rabbit treatment with CoQ10 on embryo quality. Also, potential competence of embryos to reach blastocyst stage (expanded or hatched) produced from does in G2 and G3 supported the impact of treatment with either CoQ10 or LC as compared to control on in vivo production and embryo transfer in rabbits. In addition, LC is essential for the normal mitochondrial oxidation of fatty acids and excretion of acyl-coenzyme A (acyl-CoA) esters and affects adenosine triphosphate (ATP) levels (Vanella et al., 2000) and has a pivotal role in mitochondrial oxidation of long-chain fatty acids which increase energy supply to the cell (Zhou et al., 2007). The LC plays an important role in the lipids/fatty acid metabolism as an endogenous energy source for the embryos. The LC is an enhancer of lipid metabolism in animal cells; it has a role in the transport of fatty acids from the cytosol to the mitochondria to fuel beta-oxidation (Kruip et al., 1983; McEvoy et al., 2000). Two major functions have been identified for the carnitine system, firstly facilitating the transport of long-chain fatty acids into mitochondria for their utilization in energy generating processes and secondly facilitating the removal of short-chain and medium-chain fatty acids from mitochondria that accumulate as a result of normal and abnormal metabolism (Peluso et al., 2000). The observed increase in developmental competence of oocytes matured with CoQ10 during the fall is at least in part an outcome of the changes in gene transcription levels reported for CoQ10-treated oocytes. In this way, Yi et al. (2007) found an increase in progression to blastocyst stage with no untoward effects when mitochondrial concentrates were injected into mouse 2 pronuclear stage embryos. The later study commented on the need to avoid heteroplasmy of mitochondria to maintain appropriate nuclear-mitochondrial communication required for optimal embryo development. The CoQ10 plays a key role in the mitochondrial electron transport chain, it is a critical coenzyme in the synthesis of ATP (Abdulhasan et al., 2015). Dietary supplementation with CoQ10 may increase mitochondrial activity within the developing embryo (Bentov et al., 2010). The CoQ10 is a coenzyme that aids in the transport of electrons along the mitochondrial respiratory chain. The CoQ10 administration is effective in the treatment of a variety of pathological conditions, including hyper-triglyceridemia and Friedrich’s ataxia. Dietary supplementation with CoQ10 may improve embryo quality, especially in women of advanced reproductive age (Chappel, 2013). Also, it was reported that CoQ10 increased the ATP content in expanded blastocysts (Stojkovic et al., 1999) and stimulated ATP formation in myocardial cells of mouse fetuses (Kishi et al., 1993). Subsequently, Marriage et al. (2004) reported a superior rate of early bovine embryo cleavage, blastocyst formation, percentage of expanding blastocysts and a larger inner cell mass as affected by CoQ10. These changes were associated with an increased ATP content in the group of embryos cultured with CoQ10 (Marriage et al., 2004). In addition, CoQ10 is antioxidant that has great importance against free radicals, with its tissue concentration, being 5-10-fold higher than the other main lipid soluble antioxidant (Bentinger et al., 2007). Also, it protects the stability of the cell membrane, DNA from free radicals induced oxidative damage and helps recycling of vitamin E and maintain healthy energy levels (El-Tohamy et al., 2012). Dietary supplementation with CoQ10 may increase mitochondrial activity and function within the developing embryo (Bentov et al., 2010). In conclusion, pre-conception daily oral dose of rabbit does for 21 days with 10 mg CoQ10/kg LBW has impact on ovulation rate, yield and quality of fresh embryos, survival and viability of vitrified embryos and developmental competence of in vitro embryo culture.
REFERENCES
- Abdelrazik, H., R. Sharma, R. Mahfouz and A. Agarwal, 2009. L-carnitine decreases DNA damage and improves the in vitro blastocyst development rate in mouse embryos. Fertil. Steril., 91: 589-596.
CrossRefDirect Link - Abdulhasan, M.A.K., M.B.M.R. Fakhrildin and M.H. Shubber, 2015. Effect of CoQ10 addition to maturation medium after vitrification of bovine oocytes. Int. J. Sci. Nat., 6: 40-44.
Direct Link - Agarwal, A., D. Durairajanayagam and S.S. du Plessis, 2014. Utility of antioxidants during assisted reproductive techniques: An evidence based review. Reprod. Biol. Endocrinol., Vol. 12.
Direct Link - Bentinger, M., K. Brismar and G. Dallner, 2007. The antioxidant role of coenzyme Q. Mitochondrion, 7: S41-S50.
CrossRefDirect Link - Bentov, Y., N. Esfandiari, E. Burstein and R.F. Casper, 2010. The use of mitochondrial nutrients to improve the outcome of infertility treatment in older patients. Fertil. Steril., 93: 272-275.
CrossRefDirect Link - Du Plessis, S.S., K. Makker, N.R. Desai and A. Agarwal, 2008. Impact of oxidative stress on IVF. Expet Rev. Obstet. Gynecol., 3: 539-554.
CrossRefDirect Link - Dufy-Barbe, L., P. Franchimont and J.M.A. Faure, 1973. Time-courses of LH and FSH release after mating in the female rabbit. Endocrinology, 92: 1318-1321.
CrossRefDirect Link - Eder, K., 2009. Influence of l-carnitine on metabolism and performance of sows. Br. J. Nutr., 102: 645-654.
CrossRefDirect Link - El‐Tohamy, M.M., M.S. Kotp, W.S. El‐Nattat and A.H. Mohamed, 2012. Semen characteristics and oxidative/antioxidative status in semen and serum of male rabbits supplemented with antioxidants during heat stress. Iran. J. Applied Anim. Sci., 2: 175-183.
Direct Link - Gardner, D.K., P.L. Wale, R. Collins and M. Lane, 2011. Glucose consumption of single post-compaction human embryos is predictive of embryo sex and live birth outcome. Hum. Reprod., 26: 1981-1986.
CrossRefDirect Link - Gott, A.L., K. Hardy, R.M.L. Winston and H.J. Leese, 1990. Non-invasive measurement of pyruvate and glucose uptake and lactate production by sigle human preimplantation embryos. Hum. Reprod., 5: 104-108.
Direct Link - Guerin, P., S. El Mouatassim and Y. Menezo, 2001. Oxidative stress and protection against reactive oxygen species in the pre-implantation embryo and its surroundings. Hum. Reprod Update, 7: 175-189.
CrossRefPubMedDirect Link - Kapoor, P. and A.K. Kapoor, 2013. Coenzyme Q10-a novel molecule. J. Indian Acad. Clin. Med., 14: 37-45.
Direct Link - Kishi, T., T. Okamoto, T. Takahashi, K. Goshima and T. Yamagami, 1993. Cardiostimulatory action of coenzyme Q homologues on cultured myocardial cells and their biochemical mechanisms. Clin. Investigator, 71: S71-S75.
CrossRefDirect Link - Kruip, T.A.M., D.G. Cran, T.H. van Beneden and S.J. Dieleman, 1983. Structural changes in bovine oocytes during final maturation in vivo. Gamete Res., 8: 29-47.
CrossRefDirect Link - Marriage, B.J., M.T. Clandinin, I.M. Macdonald and D.M. Glerum, 2004. Cofactor treatment improves ATP synthetic capacity in patients with oxidative phosphorylation disorders. Mol. Genet. Metab., 81: 263-272.
CrossRefPubMedDirect Link - McEvoy, T.G., G.D. Coull, P.J. Broadbent, J.S. Hutchinson and B.K. Speake, 2000. Fatty acid composition of lipids in immature cattle, pig and sheep oocytes with intact zona pellucida. J. Reprod. Fertil., 118: 163-170.
CrossRefDirect Link - Men, H., Y. Agca, L.K. Riley and J.K. Critser, 2006. Improved survival of vitrified porcine embryos after partial delipation through chemically stimulated lipolysis and inhibition of apoptosis. Theriogenology, 66: 2008-2016.
CrossRefDirect Link - Ogawa, B., S. Ueno, N. Nakayama, H. Matsunari and K. Nakano et al., 2010. Developmental ability of porcine in vitro matured oocytes at the meiosis II stage after vitrification. J. Reprod. Dev., 56: 356-361.
CrossRefDirect Link - Peluso, G., R. Nicolai, E. Reda, P. Benatti, A. Barbarisi and M. Calvani, 2000. Cancer and anticancer therapy-induced modifications on metabolism mediated by carnitine system. J. Cell. Physiol., 182: 339-350.
CrossRefDirect Link - Picton, H.M., K. Elder, F.D. Houghton, J.A. Hawkhead and A.J. Rutherford et al., 2010. Association between amino acid turnover and chromosome aneuploidy during human preimplantation embryo development in vitro. Mol. Hum. Reprod., 16: 557-569.
CrossRefDirect Link - Rizzo, A.M., P. Berselli, S. Zava, G. Montorfano, M. Negroni, P. Corsetto and B. Berra, 2010. Endogenous antioxidants and radical scavengers. Adv. Exp. Med. Biol., 698: 52-67.
CrossRefDirect Link - Chappel, S., 2013. The role of mitochondria from mature oocyte to viable blastocyst. Obstetr. Gynecol. Int.
CrossRefDirect Link - Stojkovic, M., K. Westesen, V. Zakhartchenko, P. Stojkovic, K. Boxhammer and E. Wolf, 1999. Coenzyme Q10 in submicron-sized dispersion improves development, hatching, cell proliferation and adenosine triphosphate content of in vitro-produced bovine embryos. Biol. Reprod., 61: 541-547.
Direct Link - Sturmey, R.G., D.R. Brison and H.J. Leese, 2008. Assessing embryo viability by measurement of amino acid turnover. Reprod. Biomed. Online, 17: 486-496.
CrossRefDirect Link - Takahashi, T., Y. Inaba, T. Somfai, M. Kaneda, M. Geshi, T. Nagai and N. Manabe, 2013. Supplementation of culture medium with L-carnitine improves development and cryotolerance of bovine embryos produced in vitro. Reprod. Fertil. Dev., 25: 589-599.
CrossRefDirect Link - Vanella, A., A. Russo, R. Acquaviva, A. Campisi, C. Di Giacomo, V. Sorrenti and M.L. Barcellona, 2000. L-propionyl-carnitine as superoxide scavenger, antioxidant and DNA cleavage protector. Cell Biol. Toxicol., 16: 99-104.
CrossRefDirect Link - Ye, J., J. Li, Y. Yu, Q. Wei, W. Deng and L. Yu, 2010. L-carnitine attenuates oxidant injury in HK-2 cells via ROS-mitochondria pathway. Regul. Pept., 161: 58-66.
CrossRefDirect Link - Yi, Y.C., M.J. Chen, J.Y.P. Ho, H.F. Guu and E.S.C. Ho, 2007. Mitochondria transfer can enhance the murine embryo development. J. Assisted Reprod. Genet., 24: 445-449.
CrossRefDirect Link - Zhou, X., F. Liu and S. Zhai, 2007. Effect of L-carnitine and/or L-acetyl-carnitine in nutrition treatment for male infertility: A systematic review. Asia Pac. J. Clin. Nutr., 16: 383-390.
Direct Link