
Ahmed Abdelbaset-Ismail
Department of Surgery, Anesthesiology and Radiology, Faculty of Veterinary Medicine, Zagazig University, El-Sharkia, Egypt
LiveDNA: 20.11794
Mudasir Bashir Gugjoo
Division of Surgery, Indian Veterinary Research Institute, Izatnagar, India
Alaa Ghazy
Department of Surgery, Anesthesiology and Radiology, Faculty of Veterinary Medicine, Kafrelsheikh University, Kafrelsheikh, Egypt
LiveDNA: 20.1024
Mohamed Gomaa
Department of Surgery, Anesthesiology and Radiology, Faculty of Veterinary Medicine, Zagazig University, El-Sharkia, Egypt
Ahmed Abdelaal
Department of Animal Medicine, Faculty of Veterinary Medicine, Zagazig University, El-Sharkia, Egypt
Amarpal
Division of Surgery, Indian Veterinary Research Institute, Izatnagar, India
LiveDNA: 91.5019
Ahmed Behery
Department of Surgery, Anesthesiology and Radiology, Faculty of Veterinary Medicine, Zagazig University, Ee-Sharkia, Egypt
Abdel-Basit Abdel-Aal
Department of Surgery, Anesthesiology and Radiology, Faculty of Veterinary Medicine, Zagazig University, El-Sharkia, Egypt
Mohamed-Tayiser Samy
Department of Surgery, Anesthesiology and Radiology, Faculty of Veterinary Medicine, Zagazig University, El-Sharkia, Egypt
Kuldeep Dhama
Division of Pathology, ICAR-Indian Veterinary Research Institute, Izatnagar, India
LiveDNA: 91.4710
Asian Journal of Animal and Veterinary Advances
Year: 2016 | Volume: 11 | Issue: 3 | Page No.: 204-209







ABSTRACT
There is no doubt that the donkey, as a drafting animal, plays a critical role in the developing communities for both agriculture production and riding purposes. Therefore, to maintain healthy skeleton in these animals, the understanding of radiographic anatomy of ossification centers and growth plates (locations, types and closure times), need to be advanced. It is imperative that an appropriate maturation point is reached to avoid early overexertion of the donkey and the resulting irreparable damage. It is therefore the aim of this review is to highlight the importance of expanding the limited dataset on epiphyseal growth plate maturation and begin to define its ossification times in the donkey. Moreover, it has been recently proposed that the receptors of gonadal sex hormones including estrogen androgen as well as pituitary sex hormones including luteinizing hormone and prolactin are localized in human and animal growth plates. This indicates that these hormones actively participate in the chondroregulation mechanisms at the ossification growth plates. A connection with that, we noticed that the more primitive stem cells as well as hematopoietic stem cells and mesenechymal stem cells express these receptors as well. Therefore, it would be very interesting to address the link between sex hormones and ossification time of growth plates, as well as the sharing activities of these stem cells during the ossification process.
PDF Abstract XML References Citation
How to cite this article
Ahmed Abdelbaset-Ismail, Mudasir Bashir Gugjoo, Alaa Ghazy, Mohamed Gomaa, Ahmed Abdelaal, Amarpal, Ahmed Behery, Abdel-Basit Abdel-Aal, Mohamed-Tayiser Samy and Kuldeep Dhama, 2016. Radiographic Specification of Skeletal Maturation in Donkeys: Defining the Ossification Time of Donkey Growth Plates for Preventing Irreparable Damage. Asian Journal of Animal and Veterinary Advances, 11: 204-209.
DOI: 10.3923/ajava.2016.204.209
URL: https://scialert.net/abstract/?doi=ajava.2016.204.209
DOI: 10.3923/ajava.2016.204.209
URL: https://scialert.net/abstract/?doi=ajava.2016.204.209
INTRODUCTION
It is well known that the ossification centers are potential indicators for the skeletal maturity in both human and animals (Fukuda and Matsuoka, 1979; O’Connor et al., 2013; Ogden et al., 1981). Each ossification center consists originally of hyaline cartilage and is interpreted by a radiographer as a radiolucent line bordered by slightly higher radiopacities (Von Pfeil and DeCamp, 2009). Next, with increase in age, this cartilage undergoes ossification and is accompanied by decreases in their radiolucency; this process is named endochondral ossification (Kim et al., 2001; Von Pfeil and DeCamp, 2009).
Therefore, from the practical point of view, the study of ossification centers is of great importance, not only in the determination of age and hence acquired information about the starting time for pushing animals into work or training, but also for understanding both normal and abnormal developmental orthopedic diseases such as osteochondritis dissecans (OCD) and physitis (Strand et al., 2007; Lepeule et al., 2011).
In support of this, many studies proposed the timing rate of some epiphyseal and apophyseal ossifications centers in horses (Myers and Emmerson, 1966; Banks, 1972; Brown and MacCallum, 1975; Fretz et al., 1984; Smith et al., 1991; Koskinen and Katila, 1997; Vulcano et al., 1997; Uhlhorn et al., 2000; Strand et al., 2007), goat (Genccelep et al., 2012), dogs (Fukuda and Matsuoka, 1980), cats (Vazquez-Auton et al., 1993), black beef cattle (Oishi et al., 1996), monkey (Hofmann et al., 2007), deer (Flinn et al., 2013) and mice (Fukuda and Matsuoka, 1980).
However, there are very few studies concerning the epiphyseal closure times in donkeys (Ahmed, 2005; Behery et al., 2007; Samy et al., 2007). The aim of this review is therefore to gather the information in the context of ossification time rate in donkeys and to compare this data with information previously documented in horses.
Why skeletal maturity is important: Because ossification centers at younger ages are composed of cartilage, it makes these points weaker. Pushing the animals into extra hard work at these ages, results in many distortion and conformational problems in these growth plates (Shapiro, 1987; Soeta et al., 2000). In this context, understanding the skeletal maturity in horses, mules or donkeys, could aid in capturing the proper age of maturity.
For instance, bone maturity is slow in Arabian horses and donkeys (Myers and Emmerson, 1966; Ahmed, 2005; Behery et al., 2007; Samy et al., 2007). Based on that, these latter animals should not be used for riding before three to four years old and any hard work should be avoided until they are at least five to six years old, in spite of these animals appearing physically mature at ages less than 3 years old (Brown and MacCallum, 1975; Ogden et al., 1981; Samy et al., 2007; Tavana and Peighambarzadeh, 2014).
In addition, premature ossification of growth plates or uneven maturation results from certain practices including unbalanced hoof trimming, leading to conformational defects such as open knees and knock-knees. Moreover, the proper radiographic assessment of the epiphyseal closure times of young animal skeletons allows veterinarians to determine whether there are delayed or premature closure defects (Mason and Bourke, 1973; Salter and Harris, 2001).
Types of the growth plates in equines: Bone formation in immature animals is fulfilled by either intramembranous ossification or endochondral ossification. In the case of intramembranous ossification, the bone is formed by differentiation of the connective tissue cells into bone cells and this mechanism occurs mainly in flat bones (e.g., cranium and scapula) where it leads to increases in bone width. While in case of endochondral ossification, the bone is formed by transformation of chondrogenic cells into bone cells, where it causes increases in bone length in addition to the shaping of articular surfaces (Summerlee, 2002). The latter process is responsible for structuring the three long bone regions: the epiphysis, metaphysis and diaphysis (Boskey, 2002) (Fig. 1).
There are two main types of ossification centers: primary and secondary ossification centers. During prenatal life, the diaphysis is developed from the primary ossification center consisting of a mixture of pluripotent mesenchymal stem cells, calcifying chondrocytes and surrounding thin sheet of cancellous bone (Boskey, 2002). Concerning postnatal life, a recent study evidenced that there are highly specific pluripotent mesenchymal stem cells, so called skeletal stem cells, located in growth plates and during the growth period these cells form osteoblasts and chondrocytes at the area of growth plates (Chan et al., 2015). Regarding the secondary ossification center, pressure epiphyses (at the ends of long bones) and traction epiphyses (apophyses) are considered the secondary ossification centers (Fig. 1). Apophyses are located at the point of insertion or origin of muscles (e.g., tuber-calcis, dorsal tibia tuberosity, lateral and medial tibia malleoli).
The epiphysis is growing from the vascular reserve cartilage zone (which is responsible for the growth of the epiphysis toward articular surfaces) and the epiphyseal growth plate (physis), which is composed from hyaline cartilage between the epiphysis and the metaphysis, responsible mainly for growing the bone in length.
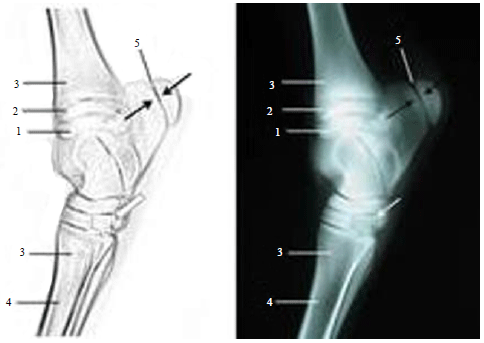 | |
| Fig. 1: | Mediolateral radiograph of normal hock joint of a one month-old donkey foal. Note a clear fusion surface of both tuber-calcis and calcaneus proper (black arrows) and fused first and second tarsal bones (white arrow). The epiphyseal and apophyseal growth plates are visible as a clear radiolucent line between the epiphysis-metaphysis (2) and tubercle-main bone (5), respectively (Ahmed, 2005). (1) Pressure epiphysis (epiphysis), (2) Epiphyseal growth plate (physis), (3) Metaphysis, (4) Diaphysis (shaft) and (5) Traction epiphysis (apophysis) |
The third part “metaphysis” is present between diaphysis and epiphysis. The metaphysis is developed from the contacting zones of the growth plate (Shapiro, 1987; Salter and Harris, 2001; Olsson and Ekman, 2002).
Cartilaginous, fibrous and bony parts are the main compositions of the growth plate (Brighton, 1987; Burdan et al., 2009). The cartilaginous part consists of three major zones: The reserve, proliferative and hypertrophic zones. The latter zone is subdivided into maturation, degeneration, primary spongiosa and secondary spongiosa subzones (Abad et al., 2002). This order of zones is directed from epiphysis toward metaphysis. The fibrous part surrounds the cartilaginous part and “looks like a ring” for supporting the physis and protecting it from shear stresses (Deppermann et al., 1989). The bony parts are located on either side (dorsal and ventral) of the growth plate (Shapiro, 1987; Soeta et al., 2000; Von Pfeil et al., 2009).
Growth plates ossification times in donkeys: Donkeys, as drafting animals, participate actively in developing communities for agriculture production and riding purposes. The number of donkeys in the world has reached approximately 44 million (Tavana and Peighambarzadeh, 2014). For this reason and from our view, greater understanding of the skeleton of the donkey is necessary, especially in context of growth plate closure times, since there is no data currently available. Our previous results provided beneficial information in that context (Ahmed, 2005). Table 1 and 2 shows the approximate closure times of stifle and hock joints of donkeys from our study and compares this data with those previously published in horses.
Our previous study was carried out on clinically apparent healthy Equus africanus asinus donkeys (n = 66) with ages from 1 day to 4 years old. In all animals, the stifle and hock joints were clinically and radiographically investigated. The following radiographic projections were employed; lateromedial, caudocranial for the stifle joint and lateromedial, mediolateral, dorsoplantar for hock joint. The exposure criteria used for radiography were 66-74 kVp, 1-10 mAs for the stifle joint imaging and 50-58 kVp, 1-6 mAs for tarsal joint imaging. In all radiographs, Focal Film Distance (FFD) 100 cm was used. We found that most of the growth plates of stifle and hock joints become ossified (mature) at approximately 4 years of age (Ahmed, 2005; Behery et al., 2007; Samy et al., 2007).
| Table 1: | Approximate closure times of the ossification centers of stifle joint in horses and donkeys, as determined by radiography |
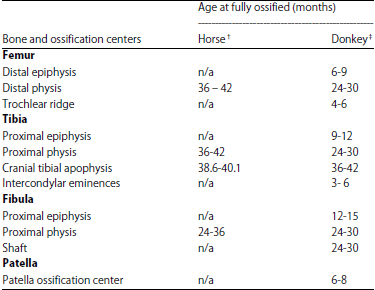 | |
| n/a: Not available, †: Banks et al. (1969), Smith et al. (1991) and Strand et al. (2007), ‡: Ahmed (2005) | |
| Table 2: | Approximate closure times of the ossification centers of tarsal joint in horses and donkeys |
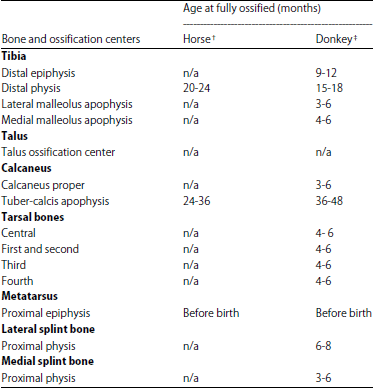 | |
| n/a: Not available, †: Banks et al. (1969), Smith et al. (1991) and Strand et al. (2007), ‡: Ahmed (2005) | |
Moreover, like horses, donkey foals under six months of age are rarely affected by upward fixation of the patella because medial and lateral femoral trochlear ridges are approximately of equal height (Ahmed, 2005; Samy et al., 2007). Other recent report proposed that Icelandic horses reach to their full appendicular skeleton maturity at approximately 3 years of age (Strand et al., 2007).
Unlike donkeys, based on other results, Arabian horses reach to complete fusion of their tuber-olecranon, as a last one do fuse, at 26.6-29.7 months of age (Myers and Emmerson, 1966). Further, more interestingly, the most of epiphyseal lines in donkeys fuse initially at the middle third and followed by their plantar and finally the dorsal thirds (Ahmed, 2005).
Future directions: The highly organized process of forming cartilage and after wards its remodeling into ossified tissues is named “endochondral ossification”. There are several well-known biological factors which orchestrate this mechanism in-situ, such as bone morphogens (BMP2, BMP4, BMP7), vitamin D3 and chondrocyte growth factor, ect. (Dziewiatkowski, 1954; Kasper et al., 1982; Tsumaki and Yoshikawa, 2005). In addition to these factors, it has also been proposed that the gonadal and pituitary sex hormones (SexHs) share actively in this mechanism. These studies showed, at molecular and proteomic level, the localization of the SexHs receptors in human and animal fetal growth plates (Ben-Hur et al., 1997; Kusec et al., 1998; Nilsson et al., 2003). In so doing, this confirms the regulatory effect of the SexHs during the various stages of endochondral ossification including proliferation, differentiation and remodeling phases. Moreover, other studies revealed that follicle stimulating hormone, prolactin and luteinizing hormone have shown as key players in development and ossification of growth plate as well as bone remodeling (Allan et al., 2010; Seriwatanachai et al., 2012). More recently, our research study exclusively provide evidence that primitive population of AC133+Lin-CD45- very small embryonic-like stem cells as well as AC133+Lin-CD45+ hematopoietic stem cells and mesenchymal stem cells express the functional SexH receptors (Abdelbaset-Ismail et al., 2016). Based on these exciting data, the intriguing orchestration mechanism managed by SexHs of the endochondral ossification timing in donkey and horses, from our point of view, is important to address for better understanding and enhancing their skeletal maturation.
CONCLUSION
The rate of skeletal maturity of donkeys needs to be characterized further to avoid any problems during the maturation period, namely developmental orthopedic diseases, including physitis and OCD and also to provide information to farmers and donkey users about the proper starting age for working and training the donkey like their partner horse.
ACKNOWLEDGMENT
This work was supported by Faculty of Veterinary Medicine, Zagazig University, Egypt.
REFERENCES
- Abad, V., J.L. Meyers, M. Weise, R.I. Gafni and K.M. Barnes et al., 2002. The role of the resting zone in growth plate chondrogenesis. Endocrinology, 143: 1851-1857.
PubMedDirect Link - Abdelbaset-Ismail, A., M. Suszynska, S. Borkowska, M. Adamiak, J. Ratajczak, M. Kucia and M.Z. Ratajczak, 2016. Human haematopoietic stem/progenitor cells express several functional sex hormone receptors. J. Cell. Mol. Med., 20: 134-146.
CrossRefDirect Link - Allan, C.M., R. Kalak, C.R. Dunstan, K.J. McTavish, H. Zhou, D.J. Handelsman and M.J. Seibel, 2010. Follicle-stimulating hormone increases bone mass in female mice. Proc. Natl. Acad. Sci. USA., 107: 22629-22634.
CrossRefDirect Link - Banks, W.C., 1972. Radiography of the Tuber calcis and its use in Thoroughbred training. Acta Radiologica: Diagn., 12: 71-72.
CrossRefDirect Link - Behery, A.E., M.T. Samy, I.A.A. Shaaban, A.M. Abd El Aal, A.A. Ahmed and J. Al-Qahirah, 2007. Determination of epiphyseal line closure time of the hock joint in donkeys. Vet. Med. J. Giza, 55: 45-56.
Direct Link - Ben-Hur, H., H.H. Thole, A. Mashiah, V. Insler and V. Berman et al., 1997. Estrogen, progesterone and testosterone receptors in human fetal cartilaginous tissue: Immunohistochemical studies. Calcified Tiss. Int., 60: 520-526.
CrossRefDirect Link - Brown, M.P. and F.J. MacCallum, 1975. A system of grading ossification in limbs of foals to assist in radiologic interpretation. Am. J. Vet. Res., 36: 655-661.
Direct Link - Burdan, F., J. Szumilo, A. Korobowicz, R. Farooquee and S. Patel et al., 2009. Morphology and physiology of the epiphyseal growth plate. Folia Histochemica Cytobiologica, 47: 5-16.
CrossRefPubMedDirect Link - Chan, C.K.F., E.Y. Seo, J.Y. Chen, D. Lo and A. McArdle et al., 2015. Identification and specification of the mouse skeletal stem cell. Cell, 160: 285-298.
CrossRefDirect Link - Deppermann, F., M. Dallek, N. Meenen, D. Lorke and K.H. Jungbluth, 1989. [The biomechanical significance of the periosteum for the epiphyseal groove]. Unfallchirurgie, 15: 165-173, (In German).
PubMedDirect Link - Dziewiatkowski, D.D., 1954. Vitamin A and endochondral ossification in the rat as indicated by the use of sulfur-35 and phosphorus-32. J. Exp. Med., 100: 11-24.
CrossRefDirect Link - Brighton, C.T., 1987. Morphology and biochemistry of the growth plate. Rheumatic Dis. Clin. North Am., 13: 75-100.
PubMedDirect Link - Flinn, E.B., B.K. Strickland, S. Demarais and D. Christiansen, 2013. Age and gender affect epiphyseal closure in white-tailed deer. Southeastern Naturalist, 12: 297-306.
CrossRefDirect Link - Fretz, P.B., N.F. Cymbaluk and J.W. Pharr, 1984. Quantitative analysis of long-bone growth in the horse. Am. J. Vet. Res., 45: 1602-1609.
PubMedDirect Link - Fukuda, S. and O. Matsuoka, 1979. Maturation process of secondary ossification centers in the rat and assessment of bone age. Jikken Dobutsu, 28: 1-9.
PubMedDirect Link - Fukuda, S. and O. Matsuoka, 1980. Comparative studies on maturation process of secondary ossification centers of long bones in the mouse, rat, dog and monkey. Jikken Dobutsu, 29: 317-326.
PubMedDirect Link - Genccelep, M., A. Karasu and O. Alpdogan, 2012. The determination of radius-ulna closure time of growth plates in mohair goat kids by radiography. Small Ruminant Res., 103: 182-186.
CrossRefDirect Link - Hofmann, M.I., C. Schradin and T. Geissmann, 2007. Radiographic evaluation of neonatal skeletal development in Callimico goeldii reveals closer similarity to Callithrix jacchus than to Saguinus oedipus. Am. J. Primatol., 69: 420-433.
Direct Link - Kasper, S., I.G. Worsley, J.M. Rowe, R.P. Shiu and H.G. Friesen, 1982. Chondrocyte growth factor from the human pituitary gland. J. Biol. Chem., 257: 5226-5230.
Direct Link - Kim, H.K.W., P.H. Su and Y.S. Qiu, 2001. Histopathologic changes in growth-plate cartilage following ischemic necrosis of the capital femoral epiphysis: An experimental investigation in immature pigs. J. Bone Joint Surg. Am., 83: 688-697.
Direct Link - Koskinen, E. and T. Katila, 1997. Effect of 19-norandrostenololylaurate on serum testosterone concentration, libido and closure of distal radial growth plate in colts. Acta Veterinaria Scandinavica, 38: 59-67.
PubMedDirect Link - Kusec, V., A.S. Virdi, R. Prince and J.T. Triffitt, 1998. Localization of estrogen receptor-α in human and rabbit skeletal tissues. J. Clin. Endocrinol. Metab., 83: 2421-2428.
CrossRefPubMedDirect Link - Lepeule, J., H. Seegers, V. Rondeau, C. Robert, J.M. Denoix and N. Bareille, 2011. Risk factors for the presence and extent of Developmental Orthopaedic Disease in the limbs of young horses: Insights from a count model. Prevent. Vet. Med., 101: 96-106.
CrossRefDirect Link - Mason, T.A. and J.M. Bourke, 1973. Closure of the distal radial epiphysis and its relationship to unsoundness in two year old thoroughbreds. Aust. Vet. J., 49: 221-228.
CrossRefDirect Link - Tavana, M. and S.Z. Peighambarzadeh, 2014. Radiographic findings of some metacarpophalangeal and metatarsophalangeal joints disorders in donkeys. WALIA J., 30: 198-200.
Direct Link - Myers, Jr., V.S. and M.A. Emmerson, 1966. The age and manner of epiphyseal closure in the forelegs of two Arabian foals. Vet. Radiol., 7: 39-47.
CrossRefDirect Link - Nilsson, O., D. Chrysis, O. Pajulo, A. Boman and M. Holst et al., 2003. Localization of estrogen receptors-α and -β and androgen receptor in the human growth plate at different pubertal stages. J. Endocrinol., 177: 319-326.
CrossRefDirect Link - O'Connor, J.E., J. Coyle, L.D. Spence and J. Last, 2013. Epiphyseal maturity indicators at the knee and their relationship to chronological age: Results of an Irish population study. Clin. Anatomy, 26: 755-767.
CrossRefDirect Link - Ogden, J.A., G.J. Conlogue and A.G.J. Rhodin, 1981. Roentgenographic indicators of skeletal maturity in marine mammals (Cetacea). Skeletal Radiol., 7: 119-123.
CrossRefDirect Link - Oishi, A., S. Hamada, H. Sakamoto, S. Kamiya and K. Yanagida et al., 1996. Radiographical evaluation of bone maturation in Japanese black beef cattle. J. Vet. Med. Sci., 58: 529-535.
CrossRefDirect Link - Salter, R.B. and W.R. Harris, 2001. Injuries involving the epiphyseal plate. J. Bone Joint Surg., 83: 1753-1753.
Direct Link - Samy, M.T., A.E. Behery, I.A. Shabaan, A.M.A. El Aal and A.A. Ahmed, 2007. Determination of epiphyseal lines closure time of stifle joint in donkeys. Vet. Med. J. Giza, 55: 57-65.
Direct Link - Seriwatanachai, D., N. Krishnamra and N. Charoenphandhu, 2012. Chondroregulatory action of prolactin on proliferation and differentiation of mouse chondrogenic ATDC5 cells in 3-dimensional micromass cultures. Biochem. Biophys. Res. Commun., 420: 108-113.
CrossRefDirect Link - Smith, B.L., J.A. Auer, T.S. Taylor, D.S. Hulse and M.T. Longnecker, 1991. Use of orthopedic markers for quantitative determination of proximal radial and ulnar growth in foals. Am. J. Vet. Res., 52: 1456-1460.
PubMedDirect Link - Soeta, S., R. Mori, T. Kodaka, Y. Naito and K. Taniguchi, 2000. Histological disorders related to the focal disappearance of the epiphyseal growth plate in rats induced by high dose of vitamin A. J. Vet. Med. Sci., 62: 293-299.
CrossRefDirect Link - Strand, E., L.C. Braathen, M.C. Hellsten, L. Huse-Olsen and S. Bjornsdottir, 2007. Radiographic closure time of appendicular growth plates in the Icelandic horse. Acta Veterinaria Scandinavica, Vol. 49.
CrossRefDirect Link - Tsumaki, N. and H. Yoshikawa, 2005. The role of bone morphogenetic proteins in endochondral bone formation. Cytokine Growth Factor Revi., 16: 279-285.
CrossRefDirect Link - Uhlhorn, H., P. Eksell and J. Carlsten, 2000. Scintigraphic characterization of distal radial physeal closure in young Standardbred racehorses. Vet. Radiol. Ultrasound., 41: 181-186.
CrossRefDirect Link - Vazquez-Auton, J.M., M.I.V. Lopez, F. Moreno, F. Gil and R. Latorre, 1993. Chronology of the postnatal ossification of the shoulder and elbow joints in the Siamese cat (Felis catus L.). Anat. Histol. Embryol., 22: 26-38.
CrossRefDirect Link - Von Pfeil, D.J. and C.E. DeCamp, 2009. The epiphyseal plate: Physiology, anatomy and trauma. Compend. Contin. Educ. Vet., 31: E1-E11.
PubMedDirect Link - Von Pfeil, D.J., C.E. DeCamp and S.K. Abood, 2009. The epiphyseal plate: Nutritional and hormonal influences; hereditary and other disorders. Compend. Contin. Educ. Vet., 31: E1-E13.
PubMedDirect Link - Vulcano, L.C., M.J. Mamprim, L.M.R. Muniz, A.F. Moreira and S.P.L. Luna, 1997. Radiographic study of distal radial physeal closure in thoroughbred horses. Vet. Radiol. Ultrasound., 38: 352-354.
CrossRefDirect Link