
Abdelfattah Monged Selim
Department of Animal Medicine, Faculty of Veterinary Medicine, Benha University, Moshtohor, 13736, Kaliobia, Egypt
Wolfgang Gaede
State Institute for Consumer Protection of Saxony-Anhalt, Department of Veterinary Medicine, Haferbreiter Weg Str.132-135, D-39576 Stendal, Germany
Asian Journal of Animal and Veterinary Advances
Year: 2015 | Volume: 10 | Issue: 11 | Page No.: 761-771







ABSTRACT
Mycobacterium avium subsp. paratuberculosis (MAP) is the etiological agent of paratuberculosis (Johne′s disease) in ruminants. The infected and carrier animals shed the microorganism intermittently in faeces. In order to ensure the sensitive identification of MAP-shedders by the examination of faecal samples. Real-time PCR assays were compared which amplify the insertion sequences IS900 and ISMAV2 and furthermore the genomic element F57. The assays were designed as duplex-PCR including the amplification of PUC19-plasmid as internal control. The analytical sensitivity of the assays was determined using DNA of 6 different isolates of MAP in broad linear range (50 ng-5 fg μL–1). The specificity was validated using 23 known species and subspecies of the Mycobacteriacea and 18 other non-Mycobacteriacea pathogens. The sensitivity for detection of MAP-DNA was 5 fg/ reaction targeting IS900. Reproducible detection limit for real-time PCR targeting ISMAV2 and F57 was 50 fg reaction. All Mycobacteriacea different from MAP and non-Mycobacteriacea gave negative results for ISMAV2 and F57 sequence. For IS900 weak positive signals were observed with highly concentrated DNA (5 ng μL–1) of 3 Mycobacterium avium subsp. avium strains from cattle and poultry but not with low concentrated DNA (5 fg μL–1). Thus false-positive results should not be found if analyzing ruminant faeces directly with IS900-PCR. ISMAV2-PCR and F57-PCR has to be preferred to IS900-PCR if real-time PCR is intended for the specification of cultured Mycobacteria.
PDF Abstract XML References Citation
How to cite this article
Abdelfattah Monged Selim and Wolfgang Gaede, 2015. Comparative Evaluation of PCR Assay for Direct Detection of Mycobacterium avium subsp. paratuberculosis in Ruminant. Asian Journal of Animal and Veterinary Advances, 10: 761-771.
DOI: 10.3923/ajava.2015.761.771
URL: https://scialert.net/abstract/?doi=ajava.2015.761.771
DOI: 10.3923/ajava.2015.761.771
URL: https://scialert.net/abstract/?doi=ajava.2015.761.771
INTRODUCTION
Mycobacterium avium subsp. paratuberculosis (MAP) is etiological cause of Johne´s disease, a chronic inflammatory enteric disease affecting cattle, sheep, goat and other ruminants. A great individual variability of subclinical and clinical symptoms the infection is typically characterized by a long incubation period followed by chronic progressive diarrhoea, decrease in milk production, oedema, anaemia loss of weight leading to cachexia and finally to death (Harris and Barletta, 2001; Donat et al., 2015; Mitchell et al., 2015).
Initially intermittent and later persistent faecal excretion of MAP starts several months up to 2.5 years before the onset of clinical signs and leads to the contamination of both the infected animal and the environment. This way the transmission of the infection mainly occurs following ingestion of MAP by offspring during the first month of life. Intrauterine transmission and direct excretion with milk are also described as additional routes of infection but their epidemiological importance is hardly to estimate (Mitchell et al., 2015; Schukken et al., 2015).
Therefore, the essential precondition for eradication of paratuberculosis from herds is the identification and elimination of MAP-shedders. In control programs, ELISAs are widely used screening assays for the detection of antibodies against MAP in blood or milk because of their advantages like easy performance, high capacity and low costs. Due to the late formation of antibodies ELISAs are characterized by low sensitivity especially in animals without or with moderate shedding of MAP in their faeces (Kohler et al., 2008; Zare et al., 2014).
Therefore, the identification of infected animals by direct detection of MAP in ruminant faeces is an indispensible tool for eradication. The diagnostic “gold standard” is faecal culture, which allows MAP-detection with high specificity and sensitivity but consumes up to 3 months until visible bacterial growth (Whitlock et al., 2000; Zare et al., 2014; Steuer et al., 2015). The diagnostic sensitivity of faecal culture can additionally be reduced by chemical decontamination steps which are necessarily implemented before cultivation to remove typical enteric bacteria. These decontamination steps can partly inactivate MAP resulting in reduced sensitivity and in false-negative results especially in the examination of subclinically infected animals that shed mycobacterium in very low doses (Stabel, 1997).
PCR-assays provide a rapid alternative for sensitive detection of MAP in clinical samples including faeces. The insertion element 900 (IS900) is the mostly used target for identification and also differentiation of MAP from other mycobacteria. The IS900 is 1.451 bp in length and is found in 15-20 copies in the MAP genome (Khare et al., 2004; Selim, 2011; Selim et al., 2013). Some recent reports indicate risk of false-positive results because IS900-like sequences can be found in M. avium subsp. avium and in some isolates of the M. intracellulare-complex (Cousins et al., 1999; Motiwala et al., 2004). Therefore, alternative genetic elements including ISMAV2 and F57 sequence have been evaluated to improve the specificity and therefore the reliability of MAP-detection by PCR.
The ISMAV2 occurs in at least three copies in MAP-cells (Stabel and Bannantine, 2005; Stratmann et al., 2006; Stevenson, 2015). Also, the F57 sequence is a single copy element of 620 bp and was found to be specific for MAP. It was not present in M. avium subsp. avium or any other mycobacteria (Tasara and Stephan, 2005; Schonenbrucher et al., 2008; Han et al., 2015). In this study, we compared the sensitivity and specificity of real-time PCR-protocols targeting the genetic elements IS900, ISMAV2 and F57 for direct detection of Mycobacterium avium subsp. Paratuberculosis.
MATERIAL AND METHODS
Bacterial strains: The specificity of the real-time PCR protocols was evaluated using 6 MAP-isolates and 23 non MAP-isolates representing 14 further mycobacterial species. Origin and detailed characterisation of these mycobacteria were described by Mobius et al. (2008). Additionally 18 non-mycobacterial pathogens, which can frequently found in faeces and other samples from cattle were examined (Table 1).
Preparation of DNA from bacterial strains: Genomic DNA of mycobacterium strains kindly provided by the Friedrich Loeffler Institut (FLI), Jena was prepared by the Cetyl-trimethyl-ammonium-bromide (CTAB) method (Van Soolingen et al., 1991).
| Table 1: | Mycobacteria and non-mycobacteria analysed for the determination of the specificity of real-time MAP-PCR |
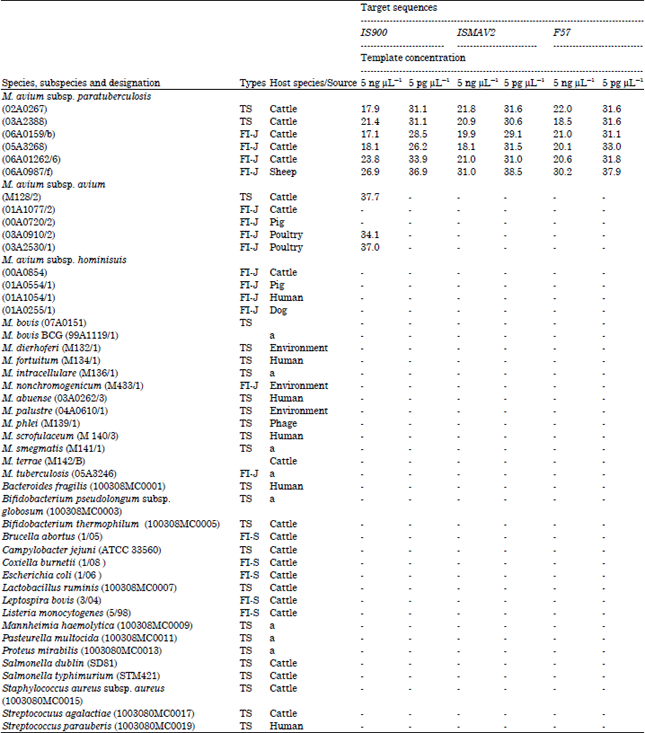 | |
TS: Reference strains of species or subspecies, FI-J: Field isolates from Germany cultivated in FLI Jena, FI-S: Field isolates from LAV Stendal, ATCC: Designation of type strains by the American Type Culture Collection, Rockville, USA, DSM: Designation of type and reference strains of the German Collection of Microorganisms and Cell Cultures, Braunschweig, Germany, Host species unknown | |
The DNA of other non-non-mycobacteria strains was isolated using the High Pure PCR Template Preparation Kit (Roche Diagnostics, Mannheim, Germany) according to the manufacturer's instructions. DNAconcentration was measured by spectrophotometer (DU640 Photometer, Beckman Coulter GmbH, Krefeld, Germany) at 260 and 280 nm.
Target sequences for MAP-detection using real-time PCR and Internal Control (IC): For detection of MAP real-time PCR targeting three different genomic regions were evaluated (Table 2). The identification of the most frequently used IS900 was based on the procedure of Khare et al. (2004). ISMAV2-PCR was self-designed using the Software Primer3 Input version 0.4.0 (Whitehead Institute for Biomedical Research, Singapore) applying the published sequence AF286339.1 (Strommenger et al., 2001). The detection of the F57 sequence was described by Schonenbrucher et al. (2008).
For monitoring of amplification conditions and the detection of PCR-inhibitors especially in faecal samples commercially available PUC19-plasmid DNA (BioRon, Ludwigshafen, Germany) was used as non-competitive internal amplification control (IC).
In order to optimize of the IC-concentration per reaction a serial dilution of PUC19-plasmid was prepared ranging from 100 to 1011 copies per ml. Based on primers and probe according to Made et al. (2008), the concentration of PUC19-plasmid was finally adjusted to 100 copies per reaction resulting in a Ct-value of about 35 when amplification of IC was combined with MAP-detection in a duplex-PCR.
Duplex real-time PCR assay (MAP with IC): Duplex real-time PCR of each target was carried out in 25 μL reaction volume. The PCR-mixtures contained 0.75 μL of both reverse and forward MAP-primer, 0.375 μL MAP-probe, 0.625 μL of both reverse and forward IC-primer, 0.25 μL IC-probe, 100 copies of PUC19-plasmid and 12.5 μL 2xQuantiTect Multiplex No Rox MasterMix (Qiagen, Hilden, Germany). Finally 5.0 μL template were added. All primers were delivered by Biotez (Berlin, Germany) and probes by Eurogentec (Belgium), respectively. They were added to the reaction mixtures in separate working solutions of 10 pmol μL–1.
The PCR reaction was performed on Stratagene Mx3005 (Agilent Technologies, Santa Clara, USA) with the following program: Initial denaturation and activation of Taq-polymerase for 15 min at 95°C followed by 45 cycles of 1 min at 94°C and 1 min at 60°C (Table 2).
Determination of analytical sensitivity by dilution of isolated MAP-DNA: Sensitivity of real-time PCR was determined by serial dilution of purified DNA from cultures of MAP- reference strain 03A3288 and field strains 05A3268, 06A0162 and 06A0159/b.
| Table 2: | Primers and probes for MAP-detection and internal amplification control (PUC19) |
 | |
DNA concentration of stock solutions was measured by spectrophotometer (DU640 Photometer, Beckman Coulter GmbH, Krefeld, Germany) at 260 and 280 nm. Afterwards serial dilutions were freshly prepared from stock solutions with sterile water in broad range from 50 ng to 0.05 fg μL–1. These serial dilutions were examined 5-fold for each target sequence, following with calculation of mean of Ct values and Standard Deviation (SD).
Determination of analytical sensitivity by dilution of ISMAV2-plasmid: The sensitivity of the self-designed ISMAV2-PCR was additionally evaluated by serial dilution of ISMAV2-plasmid ranging from 109 to 100 copies per milliliter. The ISMAV2-plasmid was derived from a PCR-product by Gene express (Berlin, Germany). Serial dilutions of ISMAV2-plasmid were examined 5-fold followed by calculation of mean of Ct-values and Standard Deviation (SD).
Analytical specificity and inclusivity: For determination of the analytical specificity of real-time PCR for IS900, ISMAV2 and F57, 23 mycobacteria species and 18 other non-mycobacteria strains were analyzed at both high and low DNA concentration of 5 ng μL–1 and 5 pg μL–1 as shown in Table 1. For inclusivity DNA-templates of 6 MAP-strain received from the FLI in Jena were applied (Table 1).
Statistical methods: Excel 2007 (Microsoft, Redmond, Washington, US) was used for tables, builds, mean values, standard deviation, correlation coefficients and regression analysis of each protocol for real-time PCR. PCR efficiency was calculated using the following equation:
Efficiency = 10(-1/slope)-1
The Kappa test was calculated using the statistical program Win Episcope 2.0 (University of Edinburgh, United Kingdom).
RESULTS
Analytical sensitivity: The detection limit of IS900 PCR based on serial dilution of isolated MAP-DNA was 5 fg reaction, i.e. approximately one MAP-genome per reaction. Rodriguez-Lazaro et al. (2005) reported 5.28 fg DNA per MAP-genome. For ISMAV2 the detection limit based on consistent positive results was 50 fg/ reaction. At the 5 fg-level one negative result in the 5-fold assay was found. The reproducible detection limit of the F57-PCR sequence was also seen at 50 fg reaction but with further reduced reproducibility at the lower concentrated DNA, i.e. only two positive reactions were observed in the 5-fold assay at the 5fg-level (Table 3). The internal control has been positive in all samples at a level of Ct-values 34-36.
The sensitivity of ISMAV2-PCR was additionally evaluated with a serial dilution of ISMAV2-plasmid from 109-100 copies per milliliter. The reproducible detection limit was 0.5 copies/reaction. At 0.05 copies/reaction the PCR assay was positive only in two PCR assay out of five repeats and with Ct-value >39 (SD, 0.14) (Table 4).
The calculated results for the analytical sensitivity are summarized in Table 5. In principles the correlation coefficients (r2) exceed 0.99 and show strong linear relationship between ct-values and the corresponding MAP-targets. Only for the amplification of diluted genomic MAP-DNA with the F57-assay r2 was slightly decreased at a level of 0.9891. The calculated efficiency of real-time PCR ranged between 92.5 and 109.8%.
| Table 3: | Analytical sensitivity recorded by Ct-values for serial dilution of quantified MAP-DNA |
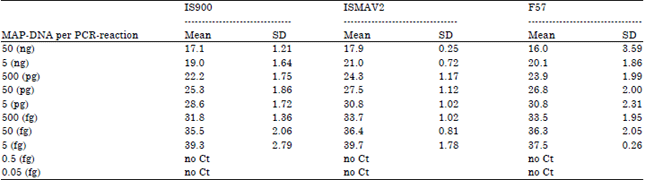 | |
*One run negative from 5-fold assays at 5 fg MAP-DNA per reaction **Only 2runs positive from 5-fold assays at 5 fg MAP-DNA per reaction | |
| Table 4: | Sensitivity of ISMAV2-PCR determined by serial dilution of ISMAV2-plasmid |
 | |
| *Only 2 runs positive from 5-fold assays at 101 MAP-plasmid μL, SD: Standard deviation | |
| Table 5: | Summarized data of determination of analytical PCR sensitivity |
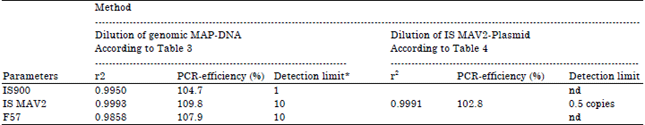 | |
*Last dilution with consitent positive results, estimated as to MAP-genoms / PCR, nd: Not deteted | |
Analytical specificity and inclusivity: All six MAP-strains analysed for the determination of the inclusivity were detected by IS900, ISMAV2 and F57 PCR protocols (Table 1). IS900-PCR showed weakly positive results for 3 of 5 Mycobacterium avium subsp. avium strains which isolated from of bovine and avian at highly concentrated DNA-templates (5 ng μL–1). For low concentrated DNA of Mycobacterium avium subsp. avium (5 pg μL–1) and also for the remaining mycobacteria and non-mycobacteria species the IS900-PCR showed the expected negative results.
ISMAV2-PCR and F57-PCR showed high specificity for detection of MAP. Both protocols showed no cross reaction with mycobacterial strains and non mycobacterial strains different from MAP.
DISCUSSION
For direct detection of MAP in several sample matrices former conventional and nested PCR-protocols are increasingly replaced by real-time PCR. The main advantages are simplified laboratory procedure due to elimination of electrophoresis, reduced risk for contaminations,especially during nested PCR and the shortening of analysis time. The application of real-time PCR essentially requires similar performance compared to established protocols for sensitive nested PCR.
Several protocols for real-time PCR have been described until now. IS900 represents the most widely applied target sequence. Alternative assays have been described for ISMAV2, the genetic element F57 und locus 251 (Tasara and Stephan, 2005; Selim et al., 2013; Fawzy et al., 2015). Analytical sensitivity of real-time MAP-PCR differs according to both target sequence and template type. In this study, results obtained for serial dilutions of genomic MAP-DNA correlate with the frequency of the target sequence in MAP-cells. The copy-number of IS900 varies between 15-20 per cell (Englund et al., 2002; Fawzy et al., 2015), thereby facilitate high sensitivity of IS900-based assays. ISMAV2 can only be found in three copies per MAP-cell resulting in a lower sensitivity than IS900-PCR (Stratmann et al., 2002; Stabel and Bannantine, 2005; Sevilla et al., 2014). The lowest analytical sensitivity seen for F57-PCR corresponds to the existence of the F57 genetic element in only one copy per MAP-cell (Tasara and Stephan, 2005; Vansnick et al., 2007; Keller et al., 2014). Different detection limits according to the cellular copy number of the target sequence were also observed by Herthnek and Bolske (2006) for IS900 and F57 whereas Schonenbrucher et al. (2008) found no difference between ISMAV2 and F57. Based on a unique plasmid (Irenge et al., 2009) described identical analytical sensitivity for IS900, F57 and ISMAP02 in triplex real-time PCR.
For comparison with further data for sensitivity of PCR the detection limit based on DNA-amount can be transformed into copy-number. With respect to a MAP-genome of 5,867,714 bp (Bannantine et al., 2002) each MAP cell theoretically contains approximately 6.8 fg of DNA. A slightly lower DNA-content of 5.1 fg MAP-DNA per MAP-cell was calculated by Ravva and Stanker (2005). Therefore 5 fg DNA can be approximated to one MAP-genome or one cell equivalent.
Using genomic MAP-DNA as template the reported detection limits was 5 fg for all targets whereas the lowest reproducible level of detection was 5 fg (~1 MAP) for IS900-PCR and 50 fg (~10 MAP) for ISMAV2 and F57. Furthermore, the detection limit was 0.5 plasmid copy/PCR reaction in ISMAV2-PCR using serial dilution of ISMAV2 plasmid. Taking methodical differences into consideration these evaluated real-time PCR assays revealed high sensitivity.
The obtained data showed that the analytical sensitivity of real-time PCR is more sensitive than previous reported limit 0.1 pg/PCR reaction of triplex real-time PCR target ISMAV2 and F57 by (Schonenbrucher et al., 2008) and 10 plasmid copies/PCR reaction in another triplex real-time PCR target IS900, ISMAP02 and F57 by Irenge et al. (2009).
In general, real-time PCR is considerable more sensitive than conventional single-step PCR for which detection limits were found to range between 1 pg and 100 fg (Stabel and Bannantine, 2005; Mobius et al., 2008). The previous reported detection limits for nested PCR are 1.2 fg (Englund et al., 1999), 1CFU ~5 fg, (Vansnick et al., 2004) and 10 copies ~50 fg, (Ikonomopoulos et al., 2004). Mobius et al. (2008) found detection limits up to 1 fg for nested PCR by direct comparison of several PCR-assays analyzing unique dilution series of MAP-DNA.
For IS900-PCR false-positive amplification for Mycobacterium avium subsp. avium was observed for highly concentrated DNA (5 fg μL–1) of one strain isolated from cattle and two poultry strains. Since, DNA from Mycobacterium avium subsp. avium is unlikely to occur in faecal samples at this high level the risk of false-positive results in clinical samples seems to be low. Tasara et al. (2005) got also false-positive results of nested IS900-PCR for M. chelonae, M. scofulaceum, M. terrae, M. xenopi and Mycobacterium strain 2333. As far as we could examine the specificity of real-time PCR targeting IS900 we did not observe cross reactions with other mycobacteria.
The absence of false-positive PCR-results for mycobacteria different from MAP and non-mycobacterial bacteria confirmed the selective specificity of the genomic element F57 and of ISMAV2 only for Mycobacterium avium subsp. paratuberculosis as described for F57 by Tasara and Stephan (2005) and Keller et al. (2014) and for ISMAV2 by Stabel and Bannantine (2005) and Han et al. (2015).
Nevertheless we conclude that ISMAV2-PCR and F57-PCR should be preferred to IS900-assays if real-time PCR is used for the specification of cultured mycobacteria.
These findings support the strategy recommended by Herthnek and Bolske (2006) to use a highly sensitive assay for MAP-screening followed by confirmation of positive screening results by at least one independent assay detecting a target sequence with highest specificity. From the present results the diagnostic procedure for direct PCR for detection of MAP in faecal samples should start with IS900-PCR as the most sensitive assay followed by ISMAV2-PCR and/or F57-PCR for confirmation. Although several MAP-target sequences are recommended for application in multiplex PCR (Tasara et al., 2005; Han et al., 2015) or in multiplex real-time PCR (Irenge et al., 2009). We preferred the simultaneous detection of only one MAP-target with an internal amplification control PCR to prevent reduced sensitivity due to competitive inhibition as observed by Rajeev et al. (2005) for duplex PCR targeting IS900 und MAP-specific locus 251.
Apart from often used house keeping genes as internal control the adjustable addition of PUC19-plasmid was used to monitor the absence of inhibitors. This way we intend to avoid any decrease of sensitivity by interaction of primers and probes or by competitive inhibition of MAP-PCR by a surplus of a house-keeping gene especially in samples with low MAP-content.
CONCLUSION
The sensitivity of alternative real-time PCR correlates with the number of target sequences in MAP-genome. These findings recommended IS900-PCR for primary screening of faecal samples. ISMAV2-PCR and/or F57-PCR should follow for confirmation of IS900 positive results. ISMAV2-PCR and F57-PCR has to be preferred to IS900-PCR if real-time PCR is intended for the specification of cultured Mycobacteria.
REFERENCES
- Bannantine, J.P., E. Baechler, Q. Zhang, L. Li and V. Kapur, 2002. Genome scale comparison of Mycobacterium avium subsp. paratuberculosis with Mycobacterium avium subsp. avium reveals potential diagnostic sequences. J. Clin. Microbiol., 40: 1303-1310.
CrossRefDirect Link - Cousins, D.V., R. Whittington, I. Marsh, A. Masters, R.J. Evans and P. Kluver, 1999. Mycobacteria distinct from Mycobacterium avium subsp. paratuberculosis isolated from the faeces of ruminants possesses IS900-like sequences detectable by IS900 polymerase chain reaction: Implications for diagnosis. Mol. Cell. Probes., 13: 431-442.
CrossRefPubMedDirect Link - Donat, K., J. Kube, J. Dressel, E. Einax, M. Pfeffer and K. Failing, 2015. Detection of Mycobacterium avium subspecies paratuberculosis in environmental samples by faecal culture and real-time PCR in relation to apparent within-herd prevalence as determined by individual faecal culture. Epidemiol. Infect., 143: 975-985.
CrossRefDirect Link - Englund, S., A. Ballagi-Pordany, G. Bolske and K.E. Johansson, 1999. Single PCR and nested PCR with a mimic molecule for detection of Mycobacterium avium subsp. paratuberculosis. Diagn. Microbiol. Infect. Dis., 33: 163-171.
CrossRefPubMedDirect Link - Englund, S., G. Bolske and K.E. Johansson, 2002. An IS900-like sequence found in a Mycobacterium sp. other than Mycobacterium avium subsp. paratuberculosis. FEMS. Microbiol. Lett., 209: 267-271.
CrossRefPubMedDirect Link - Fawzy, A., T. Eisenberg, A. El-Sayed and M. Zschock, 2015. Improvement of sensitivity for Mycobacterium avium subsp. paratuberculosis (map) detection in bovine fecal samples by specific duplex F57/IC real-time and conventional IS900 PCRs after solid culture enrichment. Trop. Anim. Health Prod., 47: 721-726.
CrossRefDirect Link - Han, J.I., Y.H. Jung, C. Choe, J. Yoo and S.J. Kang et al., 2015. Multiplex quantitative real-time polymerase chain reaction assay for rapid detection of Mycobacterium avium subsp. paratuberculosis in fecal samples. Vet. Soc. Korea, 32: 219-223.
Direct Link - Harris, N.B. and R.G. Barletta, 2001. Mycobacterium avium subsp. paratuberculosisin veterinary medicine. Clin. Microbiol. Rev., 14: 489-512.
CrossRefDirect Link - Herthnek, D. and G. Bolske, 2006. New PCR systems to confirm real-time PCR detection of Mycobacterium avium subsp. paratuberculosis. BMC Microbiol., Vol. 6.
CrossRefDirect Link - Ikonomopoulos, J., M. Gazouli, I. Pavlik, M. Bartos and P. Zacharatos et al., 2004. Comparative evaluation of PCR assays for the robust molecular detection of Mycobacterium avium subsp. paratuberculosis. J. Microbiol. Meth., 56: 315-321.
CrossRefDirect Link - Irenge, L.M., K. Walravens, M. Govaerts, J. Godfroid, V. Rosseels, K. Huygen and J.L. Gala, 2009. Development and validation of a triplex real-time pcr for rapid detection and specific identification of M. avium sub sp. paratuberculosis in faecal samples. Vet. Microbiol., 136: 166-172.
CrossRefDirect Link - Keller, S.M., R. Stephan, R. Kuenzler, M. Meylan and M.M. Wittenbrink, 2014. Comparison of fecal culture and F57 real-time polymerase chain reaction for the detection of Mycobacterium avium subspecies paratuberculosis in Swiss cattle herds with a history of paratuberculosis. Acta Veterinaria Scandinavica, Vol. 56.
Direct Link - Khare, S., T.A. Ficht, R.L. Santos, J. Romano and A.R. Ficht et al., 2004. Rapid and sensitive detection of Mycobacterium avium subsp. paratuberculosis in bovine milk and feces by a combination of immunomagnetic bead separation-conventional PCR and real-time PCR. J. Clin. Microbiol., 42: 1075-1081.
CrossRefPubMedDirect Link - Kohler, H., B. Burket, I. Pavlik, R. Diller, L. Geue, F.J. Conraths and G. Martin, 2008. Evaluation of five ELISA test kits for the measurement of antibodies against Mycobacterium avium subspecies paratuberculosis in bovine serum. Berliner Munchener Tierarztliche Wochenschrift, 121: 203-210.
PubMedDirect Link - Made, D., R. Reiting, E. Strauch, K. Ketteritzsch and A. Wicke, 2008. A real-time PCR for detection of pathogenic Yersinia enterocolitica in food combined with an universal internal amplification control system. J. Verbraucherschutz Lebensmittelsicherheit, 3: 141-151.
CrossRefDirect Link - Mitchell, R.M., Y. Schukken, A. Koets, M. Weber and D. Bakker et al., 2015. Differences in intermittent and continuous fecal shedding patterns between natural and experimental Mycobacterium avium subspecies paratuberculosis infections in cattle. Vet. Res., Vol. 46.
CrossRefDirect Link - Mobius, P., H. Hotzel, A. Raβach and H. Kohler, 2008. Comparison of 13 single-round and nested PCR assays targeting IS900, ISMav2, f57 and locus 255 for detection of Mycobacterium avium subsp. paratuberculosis. Vet. Microbiol., 126: 324-333.
CrossRefDirect Link - Motiwala, A.S., A. Amonsin, M. Strother, E.J.B. Manning, V. Kapur and S. Sreevatsan, 2004. Molecular epidemiology of Mycobacterium avium subsp. paratuberculosis isolates recovered from wild animal species. J. Clin. Microbiol., 42: 1703-1712.
CrossRefDirect Link - Rajeev, S., Y. Zhang, S. Sreevatsan, A.S. Motiwala and B. Byrum, 2005. Evaluation of multiple genomic targets for identification and confirmation of Mycobacterium avium subsp. paratuberculosis isolates using real-time PCR. Vet. Microbiol., 105: 215-221.
CrossRefDirect Link - Ravva, S.V. and L.H. Stanker, 2005. Real-time quantitative PCR detection of Mycobacterium avium subsp. paratuberculosis and differentiation from other mycobacteria using SYBR Green and TaqMan assays. J. Microbiol. Methods., 63: 305-317.
CrossRefPubMedDirect Link - Schonenbrucher, H., A. Abdulmawjood, K. Failing and M. Bulte, 2008. New triplex real-time PCR assay for detection of Mycobacterium avium subsp. paratuberculosis in bovine feces. Applied Environ. Microbiol., 47: 2751-2758.
PubMedDirect Link - Schukken, Y.H., R.H. Whitlock, D. Wolfgang, Y. Grohn and A. Beaver et al., 2015. Longitudinal data collection of Mycobacterium avium subspecies paratuberculosis infections in dairy herds: The value of precise field data. Vet. Res., Vol. 46.
CrossRefDirect Link - Selim, A., M. El-Haig and E.S. Galila, 2013. Direct detection of Mycobacterium avium subsp. paratuberculosis in bovine milk by multiplex real-time PCR. Biotechnol. Anim. Husbandry, 29: 513-525.
CrossRefDirect Link - Sevilla, I.A., J.M. Garrido, E. Molina, M.V. Geijo, N. Elguezabal, P. Vazquez and R.A. Juste, 2014. Development and evaluation of a novel multicopy-element-targeting triplex PCR for detection of Mycobacterium avium subsp. paratuberculosis in feces. Applied Environ. Microbiol., 80: 3757-3768.
CrossRefDirect Link - Stabel, J.R., 1997. An improved method for cultivation of Mycobacterium paratuberculosis from bovine fecal samples and comparison to three other methods. J. Vet. Diagn. Invest., 9: 375-380.
CrossRefDirect Link - Stabel, J.R. and J.P. Bannantine, 2005. Development of a nested PCR method targeting a unique multicopy element, ISMap02, for detection of Mycobacterium avium subsp. paratuberculosis in fecal samples. J. Clin. Microbiol., 43: 4744-4750.
CrossRefDirect Link - Steuer, P., J. de Waard, M.T. Collins, E.P. Troncoso, O.A. Martinez, C. Tejeda and M.A. Salgado, 2015. Identification of contaminating bacteria when attempting to isolate Mycobacterium avium subsp. paratuberculosis (MAP) from bovine faecal and tissue samples using the BACTEC MGIT 960 system. Archivos Medicina Veterinaria, 47: 97-100.
CrossRefDirect Link - Stevenson, K., 2015. Genetic diversity of Mycobacterium avium subspecies paratuberculosis and the influence of strain type on infection and pathogenesis: A review. Vet. Res., Vol. 46.
CrossRefDirect Link - Stratmann, J., K. Dohmann, J. Heinzmann and G.F. Gerlach, 2006. Peptide aMptD-mediated capture PCR for detection of Mycobacterium avium subsp. paratuberculosis in bulk milk samples. Applied Environ. Microbiol., 72: 5150-5158.
CrossRefDirect Link - Stratmann, J., B. Strommenger, K. Stevenson and G.F. Gerlach, 2002. Development of a peptide-mediated capture PCR for detection of Mycobacterium avium subsp. paratuberculosis in milk. J. Clin. Microbiol., 40: 4244-4250.
CrossRefDirect Link - Strommenger, B., K. Stevenson and G.F. Gerlach, 2001. Isolation and diagnostic potential of ISMav2, a novel insertion sequence-like element from Mycobacterium avium subspecies paratuberculosis. FEMS Microbiol. Lett., 196: 31-37.
CrossRefDirect Link - Tasara, T., L.E. Hoelzle and R. Stephan, 2005. Development and evaluation of a Mycobacterium avium subspecies paratuberculosis (MAP) specific multiplex PCR assay. Int. J. Food Microbiol., 104: 279-287.
CrossRefPubMedDirect Link - Tasara, T. and R. Stephan, 2005. Development of an F57 sequence-based real-time PCR assay for detection of Mycobacterium avium subsp. paratuberculosis in milk. Applied Environ. Microbiol., 71: 5957-5968.
CrossRefDirect Link - Van Soolingen, D., P.W.M. Hermans, P.E. de Haas, D.R. Soll and J.D. van Embden, 1991. Occurrence and stability of insertion sequences in Mycobacterium tuberculosis complex strains: Evaluation of an insertion sequence-dependent DNA polymorphism as a tool in the epidemiology of tuberculosis. J. Clin. Microbiol., 29: 2578-2586.
PubMedDirect Link - Vansnick, E., P. de Rijk, F. Vercammen, D. Geysen, L. Rigouts and F. Portaels, 2004. Newly developed primers for the detection of Mycobacterium avium subspecies paratuberculosis. Vet. Microbiol., 100: 197-204.
CrossRefPubMedDirect Link - Vansnick, E., P. de Rijk, F. Vercammen, L. Rigouts, F. Portaels and D. Geysen, 2007. A DNA sequence capture extraction method for detection of Mycobacterium avium subspecies paratuberculosis in feces and tissue samples. Vet. Microbiol., 122: 166-171.
CrossRefDirect Link - Whitlock, R.H., S.J. Wells, R.W. Sweeney and J.V. Tiem, 2000. ELISA and fecal culture for paratuberculosis (John's disease): Sensitivity and specificity of each method. Vet. Microbiol., 77: 387-398.
CrossRefPubMedDirect Link - Zare, Y., G.E. Shook, M.T. Collins and B.W. Kirkpatrick, 2014. Short communication: Heritability estimates for susceptibility to Mycobacterium avium subspecies paratuberculosis infection defined by ELISA and fecal culture test results in Jersey cattle. J. Dairy Sci., 97: 4562-4567.
CrossRefDirect Link - Rodriguez-Lazaro, D., M. D'Agostino, A. Herrewegh, M. Pla, N. Cook and J. Ikonomopulos, 2005. Real-time PCR-based methods for detection of Mycobacterium avium subsp. paratuberculosis in water and milk. Int. J. Food Micorbiol., 101: 93-104.
CrossRefPubMedDirect Link