
S.R. Mishra
Physiology and Climatology, Indian Veterinary Research Institute, Izatnagar, Bareilly, Uttar Pradesh, 243122, India
Jaya Bharati
Physiology and Climatology, Indian Veterinary Research Institute, Izatnagar, Bareilly, Uttar Pradesh, 243122, India
M.K. Bharti
Physiology and Climatology, Indian Veterinary Research Institute, Izatnagar, Bareilly, Uttar Pradesh, 243122, India
G. Singh
Physiology and Climatology, Indian Veterinary Research Institute, Izatnagar, Bareilly, Uttar Pradesh, 243122, India
M. Sarkar
Physiology and Climatology, Indian Veterinary Research Institute, Izatnagar, Bareilly, Uttar Pradesh, 243122, India
Asian Journal of Animal and Veterinary Advances
Year: 2015 | Volume: 10 | Issue: 8 | Page No.: 433-442







ABSTRACT
Fibroblast Growth Factor (FGF) family is considered as local cytokines produced from many physiological systems including ovary. The FGFs regulate many cellular processes like angiogenesis, cell proliferation, cell survival, organogenesis and embryonic development including ovarian folliculogenesis. Fibroblast Growth Factor 10 (FGF10) is one of most important member in FGF family which mostly mediates epithelial cell motility, differentiation, migration and wound healing in many cellular system. The FGF10 is also known to regulate the ovarian follicular dynamics in an autocrine and paracrine manner. Therefore, the present study was undertaken aimed with an objective to evaluate the expression and localization of FGF10 in different follicular size or groups during various stages of follicular development in buffalo ovary. The mRNA and protein expression of FGF10 transcripts did not differ significantly in all follicular size during follicular development. The FGF10 was exclusively localized in Granulosa Cell (GC) and Theca Interna (TI) showing a stage specific immunoreactivity throughout the follicular developmental process in buffalo ovary. The present findings suggest the species specific expression and localization pattern of FGF10 in buffalo ovarian follicle.
PDF Abstract XML References Citation
How to cite this article
S.R. Mishra, Jaya Bharati, M.K. Bharti, G. Singh and M. Sarkar, 2015. Expression and Localization of Fibroblast Growth Factor 10 (FGF10) in Ovarian Follicle During Different Stages Development in Buffalo. Asian Journal of Animal and Veterinary Advances, 10: 433-442.
DOI: 10.3923/ajava.2015.433.442
URL: https://scialert.net/abstract/?doi=ajava.2015.433.442
DOI: 10.3923/ajava.2015.433.442
URL: https://scialert.net/abstract/?doi=ajava.2015.433.442
INTRODUCTION
Water buffalo (Bubalus bubalis) is one of the important livestock animal mostly reared for milk, meat and draft purpose found in many Asian countries including India. Indian buffaloes contribute more than half of world’s total buffalo milk production (FAO., 2012). In spite of high productive potential certain inherent reproductive problems like delayed puberty, silent heat with poor expression of oestrus, poor conception rate and long post partum anoestrus period limit the reproductive efficiency and productivity of buffalo (Perera, 2008). Reproductive acyclicity characterized by failure of ovulation and lack of development corpus luteum subsequently affects pregnancy in buffalo (El-Wishy, 2007; Ali et al., 2009; Nam and Aiumlamai, 2010; Zicarelli, 2010). Therefore, understanding the exact mechanism of ovarian dynamics might open up new insight for the treatment of infertility which could improve the reproductive performance in buffalo.
Fibroblast Growth Factor (FGF) family consists of 23 closely related members characterized by a conserved central domain which mediate their biological activities by binding with transmembrane Receptor Tyrosine Kinase (RTK), FGFR1-FGFR4 (Powers et al., 2000; Itoh and Ornitz, 2004; Oulion et al., 2012). FGFs control various cellular processes including angiogenesis, cell proliferation, differentiation, cell survival, organogenesis and embryonic development (Basilico and Moscatelli, 1992; Thisse and Thisse, 2005; Bottcher and Niehrs, 2005; Turner and Grose, 2010).
Ovarian folliculogenesis is a complex process involving proliferation and differentiation of the follicular cells i.e., Granulosa Cells (GC) and Theca Interna (TI) (Armstrong and Webb, 1997; McGee et al., 1999) which is mostly regulated by steroids, growth hormone, gonadotropins and a variety of intra ovarian cytokines including FGFs (Webb et al., 2003). The FGFs modulates a wide variety of ovarian function including proliferation, differentiation, steroidogenesis and apoptosis of GC in many farm animal species including human suggesting their role in regulation of ovarian folliculogenesis (Lavranos et al., 1994; Vernon and Spicer, 1994; Parrott and Skinner, 1997; Roberts and Ellis, 1999; Berisha et al., 2004; Buratini et al., 2007; Matos et al., 2007; Machado et al., 2009; Portela et al., 2010).
Fibroblast growth factor 10 (FGF10), also known as Keratinocyte Growth Factor 2 (KGF2) is a 19 kDa protein which promote epithelial cell motility, differentiation, migration and wound healing (Ware and Matthay, 2002). FGF10 transcript level was significantly decreased in theca cells of mature follicles in bovine (Buratini et al., 2007; Gasperin et al., 2012), goat (Chaves et al., 2010) and in theca and stromal cells of human ovary (Taniguchi et al., 2008). FGF10 mRNA was significantly more expressed in subordinate follicle than the dominat follicle in cattle ovary (Gasperin et al., 2012). FGF10 mRNA was expressed in oocytes and theca cells while acts on GC to inhibit steroidogenesis in bovine antral follicles (Buratini et al., 2007; Zhang et al., 2010; Caixeta et al., 2013). FGF10 protein was localized in GC, TI and oocytes of bovine Preovulatory Follicle (PF) (Buratini et al., 2007). The FGF10 was involved in preantral follicular development in human ovary (Oron et al., 2012).
Based on these previous evidences, FGFs are the principal regulators ovarian steroidogensis and follicular development. However, to the best of our knowledge, no such information is available regarding the expression and localization of FGF10 in different stages of follicular development in bubaline ovary. Therefore, the current study was aimed with an objective to investigate mRNA and protein expression along with the immunohistochemical localization of FGF10 in bubaline follicle during different stages follicular development.
MATERIALS AND METHODS
Collection of follicles and preparation: Entire reproductive tracts of buffalo cows were collected from local abattoir and transported on ice within 10-20 min to laboratory. The stage of estrous cycle was defined by macroscopic observation of ovaries i.e., colour, consistency, corpus luteum stage, number and size of follicles (Sarkar et al., 2010). Only follicles which appeared healthy (well vascularised with transparent follicular wall and fluid) with diameter >3 mm were used for this study. Large follicles (>14 mm) were collected only after CL regression, with signs of mucus production in uterus and cervix and were considered as Preovulatory Follicle (PF). Follicles were dissected out from ovarian stroma and surrounding tissue (theca externa) was carefully removed with forceps under a stereo microscope (Sarkar et al., 2010) and surface diameter was determined. After aspiration of FF, each follicle was bisected and inside wall was gently scraped and flushed with Ringer’s solution to separate the Granulosa Cells (GC) and remaining follicle wall after GC separation was considered as Theca Interna (TI). GC and TI isolated from each follicle were transferred into separate tubes and labelled. GCs in flushing solution were centrifuged at 3000 g for 10 min at 4°C. TI and GC pellet were separately snap frozen in liquid nitrogen and stored at -80°C until RNA and protein isolation. FF was stored at -20°C until determination of progesterone (P4) and estradiol (E2). Because healthy follicles have relatively constant P4 concentrations in FF, only follicles with P4 below100 ng mL–1 were used for evaluation to exclude atretic follicles (Sarkar et al., 2010).
Follicle classification: Follicles were classified according to the E2 content in FF as follows, (a) <0.5 (b) 0.5-5 (c) 5-40 and (d) >180 ng mL–1. The corresponding size of follicles were in range of (a) 4-6 mm (F1/Group-I) (b) 7-9 mm (F2/Group-II) (c) 10-13 mm (F3/Group-III) and (d) >14 mm (F4/Group-IV/PF). The 50 ovaries, each with follicles were used to extract 10 follicles/group for RNA extraction, Western Blot (WB) and Immunohistochemistry (IHC) studies.
Hormone determination: Concentrations of P4 and E2 in FF were estimated by P4125 I RIA kit (Cat No. IM1188) and E2125 I RIA kit (Cat No. A21854) (Immunotech, Czech Republic) as per manufacturer’s instruction. The measurable range was 0.05-50 ng mL–1 for P4 and 6-5000 pg mL–1 for E2. The FF was diluted 1:100 with PBS. The intra and inter-assay coefficient of variation for P4 and E2 were 6.1 and 7.4% for P4 and 9.4 and 11.1% for E2, respectively.
Primers: Primers were designed by using DNASTAR-lasergene (version 6) software. Primers used in this study were synthesized by Eurofins genomics, Loxembourg, Europe. The details of primers used are given in Table 1.
Quantitative RT-PCR (Real time-Polymerase Chain Reaction) analysis: Total RNA from GC and TI (n = 10 follicles/group) from all follicular groups and cultured GCs was isolated by TRIzol reagent (Invitrogen) according to manufacturer’s instruction. Total RNA was treated with DNase-1 (Invitrogen) to exclude any possible genomic DNA contamination. Constant amounts of 1 μg of total RNA from GC and TI (n = 10/group) were reverse transcribed to complementary DNA (cDNA) by using iScriptTM Select cDNA Synthesis Kit (Bio-Rad Laboratories, USA; Cat No#170-8891) at 25°C for 5 min, 42°C for 30 min and 85°C for 5 min as per manufacturer’s instruction. The resulting cDNA was used as template in qRT-PCR study. The qRT-PCR of all target genes and 2 housekeeping genes (β2-microglobulin, β2M and ribosomal Protein S 15A, RPS15A) were performed in duplicate by using SsoFast™ EvaGreen® Supermix kit (Bio-Rad Laboratories, USA; Cat No. 172-5201AP) in Stratagene MX3005P qPCR System (Agilent technologies) as per manufacturer’s instructions. Total reaction volume of 10 μL, which consists of 0.5 μL of cDNA+0.25 μL forward primer (0.25 μM)+0.25 μL reverse primer (0.25 μM)+5 μL of SsoFast™ EvaGreen® Supermix+4 μL PCR graded nuclease free water was subjected for qPCR study as per standard real time PCR protocol. The following qPCR protocol was followed for all genes in present study (Two step protocol): segment 1 (single cycle, enzyme activation -95°C for 30 sec), segment 2 (40 cycles, denaturation -95°C for 5 sec, annealing/extension -60°C for 10 sec) followed by a melting step by slow heating from 62-95°C with a rate of 0.58°C/second with a continuous fluorescence measurement and final cooling down at 4°C.
| Table 1: | Target genes, primer sequences (5’→3’) and amplicon length for qRT-PCR used in this study |
 | |
| FGF10: Fibroblast growth factor 10, EMBL: European molecular biology laboratory | |
The Cycle threshold (Ct) values were used for relative expression study. Standard curve to check primer efficiencies were determined by amplification of a standardized dilution series of cDNA. F1 follicle (4-6 mm) was used as calibrator to obtain relative mRNA expression. β2M and RPS15A were taken as housekeeping gene and geometric mean of Ct values of β2M and RPS15A was used as Ct of reference gene. Relative quantification of mRNA was obtained by Pfaffl method Pfaffl (2001). Efficiency of primers used in this study is given in Table 1.
Antibodies: WB and IHC studies were performed by using, Rabbit polyclonal FGF10 (sc-7917, Lot#E0912, Santa Cruz Biotechnology®, Inc., USA), Goat polyclonal GAPDH (sc-20357, Lot#E2413, Santa Cruz Biotechnology®, Inc., USA), Goat anti-rabbit IgG-HRP (sc-2004, Lot#B1711, Santa Cruz Biotechnology®, Inc., USA), Mouse anti-goat IgG-HRP (sc-2354, Lot#G0910, Santa Cruz Biotechnology®, Inc., USA), Goat anti-rabbit IgG-FITC (sc-2012, Lot#I1010, Santa Cruz Biotechnology®, Inc., USA).
Western blot: Total protein extraction was done (Chouhan et al., 2013) by triturating different groups of follicular tissue (n = 10/group) with liquid nitrogen and suspended in Tissue-PELBTM (G. Biosciences, USA) with halt protease inhibitor cocktail (Thermo Scientific). Finally, different group of tissue samples were homogenized and centrifuged at 12,000 g. The supernatant containing mostly total soluble protein was estimated by Bradford protein assay. The supernatant was diluted with 2X gel loading buffer (65.8 mM Tris-HCl, pH 6.8, 2.1% SDS, 26.3% (w/v) glycerol, 0.01% BPB) and boiled for 10 min. Protein samples (40 μg) were subjected to 12 or 14% SDS-PAGE, electrotransferred onto polyvinylidene-difluoride (PVDF) membrane and blocked with 5% Bovine Serum Albumin (BSA) before incubation with primary antibodies at 1:500 dilutions overnight at 4°C. Then membrane was washed thrice with PBST (1X PBS+0.01% Tween-20) for 5 min. Secondary antibodies conjugated with horseradish peroxidase (HRP) were added to the immunoblot and incubated at 37°C for 1 h. Again the blot washed thrice in PBST solution and developed by using DAB system (Dilution buffer+3, 3’-diaminobenzidine tetrahydrochloride+0.06% H2O2) (GeNeiTM, Merck, USA) for 10 min. Bands were visualized under white light and recorded on gel documentation system (Minibus Pro; DNR Bio-Imaging Systems). Densitometry of immunospecific bands was done using Image-J 1.43U software (National Institute of Health, USA). The experiment was replicated thrice for each protein.
Immunohistochemistry: Intact follicles were carefully dissected from the ovaries and surrounding stroma along with connective tissue was removed. Freshly separated follicular tissues from different groups were fixed with 10% NBF, dehydrated through series of graded alcohols, paraffin-embedded, serially-sectioned (5 μm), mounted on Mayer’s albumin coated slides and dried at 37°C overnight (Chouhan et al., 2013). Deparaffinization was done in xylene followed by rehydration in series of graded alcohols at room temperature. Epitope retrieval was done with sodium citrate buffer (10 mM sodium citrate, pH 6.0, 0.05% Tween-20). Then slides were washed thrice with PBST for 5 min and blocked with 5% BSA at 37°C for 2 h. Subsequently, sections were treated with primary antibodies at a dilution of 1:100. Primary antibodies were detected by fluorescent conjugated secondary antibodies. Slides were washed again and subjected with DAPI (0.4 μg mL–1, 4’, 6-diamidino-2-phenylindole dihydrochloride) in PBS to stain nuclei of cells in follicular sections. Negative control slides were processed under similar conditions except primary antibody treatment. Fluorescently stained sections were mounted with antifade mounting media (MP Biomedicals) and images were captured by using an Axio Observer Z1 microscope (Carl Zeiss Micro Imaging GmbH, Germany).
Statistical analyses: All experimental data are shown as mean ± standard error of mean (SEM). The statistical significance of differences, in mRNA and protein expression of FGF10 across different follicular groups were assessed by using SPSS.17 software by one-way analysis of variance (one way ANOVA) followed by Tukey’s Honestly Significant Differences (HSD) as multiple comparison test (GraphPad PRISM v. 3.0; GraphPad Software, Inc., San Diego, CA). Differences were considered significant if p<0.05.
RESULTS AND DISCUSSION
Expression of mRNA for FGF10 in follicle: FGF10 was found to be expressed in a regulated manner with stage specific differences in expression pattern in different stages of follicular development in bubaline follicle. The mRNA expression of FGF10 in buffalo follicle was consistent in different follicular size or groups however, the relative expression varied according to the follicle size or groups. FGF10 transcript level did not differ significantly in both GC (Fig. 1a) and TI (Fig. 1b) throughout the stages of follicular development.
In present study, our data regarding FGF10 expression pattern is not in line of agreement with earlier finding in human (Taniguchi et al., 2008), goat (Chaves et al., 2010) and cattle follicle (Buratini et al., 2007; Gasperin et al., 2012) where FGF10 mRNA expression was found to be decreased in mature or healthy follicle of human, goat and bovine ovary. In another study in human preantal follicle (Oron et al., 2012) FGF10 mRNA was observed regularly which is in opposition with our present data in relation to FGF10 expression pattern in bubaline follicle. This variation in FGF10 expression could be due to the species specific expression pattern of FGF10 in different follicular groups of buffalo follicle or due to the time of tissue sample collection.
Western blot analysis: FGF10 protein expression was observed in a regulated manner during all stages of follicular development and was validated by WB analysis. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as housekeeping gene. Major bands are shown in Fig. 2.
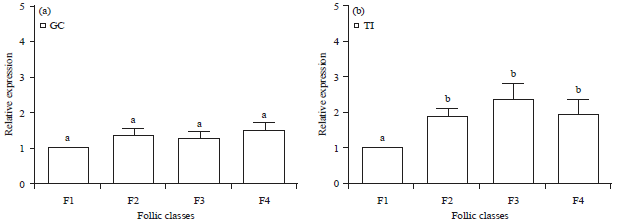 | |
| Fig. 1(a-b): | Relative mRNA expression of (a) FGF10-GC and (b) FGF10-TI in different follicular groups of buffalo. F1 was taken as calibrator. All values are shown as Mean±SEM. Different superscripts are statistically different values (p<0.05), FGF10: Fibroblast growth factor 10, GC: Granulosa cell, TI: Theca interna |
 | |
| Fig. 2(a-b): | Western blot band of (a) FGF10 and (b) GAPDH in different follicular groups of buffalo, FGF10: Fibroblast growth factor 10, GAPDH: Glyceraldehyde-3-phosphate dehydrogenase |
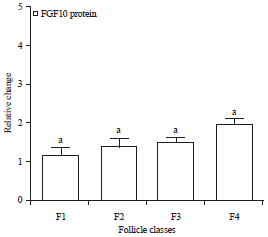 | |
| Fig. 3: | Relative change in protein expression (Mean±SEM, Band Densitometry) of FGF10 in different follicular groups of buffalo. Different superscripts denote statistically different values (p<0.05). F1 was taken as calibrator. GAPDH was used as reference protein. FGF10: Fibroblast growth factor 10, GAPDH: Glyceraldehyde-3-phosphate dehydrogenase |
The molecular weights of FGF10 and GAPDH were approximately ~19 and ~37 kDa respectively. FGF10 (Fig. 3) expression did not differ significantly in all follicular groups.
The protein expression of FGF10 in buffalo follicle was consistent throughout the follicular developmental however, their relative change varied according to the follicle size or groups. The protein expression of FGF10 did not differ significantly in all stages of follicular development. This type of variation in protein expression pattern of FGF10 might be due to its transcript level during different stages of follicular development in buffalo observed in this current study.
Immunohistochemistry of follicle: The immunoreactivity of FGF10 was observed by using IHC with a stage specific difference in reactivity. The localization of FGF10 was discernible in various cell types in different follicular groups. The intensity of immunostaining was varied in all follicular groups and number of positive cells was influenced by different stages of estrous cycle. FGF10 was exclusively localized in GC and TI in different follicular groups. The most intense immunostaining of FGF10 (Fig. 4) was observed in F4 or PF. The negative controls, with isotype IgG and without primary antibodies, showed only a weak background staining.
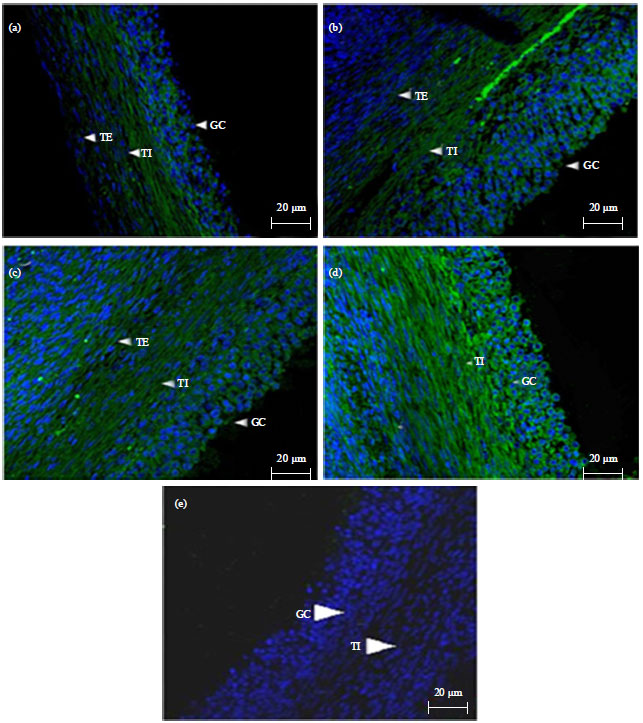 | |
| Fig. 4(a-e): | Immunohistochemical localization of FGF10 in different follicular groups of buffalo. Immunoreactivity of FGF10 in different groups of follicle was stained with FITC and merged with DAPI counter-stain (blue), indicating the nuclei of all cells in sections. Representative pictures showing immunoreactivity against FGF10 protein (a) F1, (b) F2, (c) F3, (d) F4 and (e) Negative control. Negative control sections are presented without primary antibody labelling. FGF10 protein was consistently found in GC and TI with stage specific immunostaining. Scale bar = 20 μM. FGF10: Fibroblast growth factor 10, GC: Granulosa cell, TI: Theca interna, TE: Theca externa, EC: Endothelial cell, FITC: Fluorescein isothiocyanate, DAPI, 4',6-diamidino-2-phenylindole dihydrochloride |
Our data regarding mRNA and protein expression was well supported by immunolocalization of FGF members in different follicular groups of buffalo. FGF10 was localized exclusively in GC, TI and ECs with stage specific intensity of immunostaining in various follicular groups of buffalo ovarian follicle. Our data regarding FGF10 localization in bubaline follicle coincides with the previous reports in cattle (Buratini et al., 2007) where FGF10 was localized in GC, TI and oocytes of cattle PF. In another study in human ovary (Oron et al., 2012), FGF10 protein was localized in GC and oocytes which corroborate with our current result of FGF10 protein localization in buffalo follicle in different stages of follicular development.
CONCLUSION
In conclusion, the present study is the first to demonstrate the dynamics of expression and localization of FGF10 in ovarian follicle during different stages of follicular development in buffalo. Our data clearly suggests the expression dynamics and localization of FGF10 in various stages of follicular development which indicates its involvement in ovarian folliculogenesis process in buffalo.
ACKNOWLEDGMENT
We thank Department of Biotechnology, New Delhi, India and Director, Indian Veterinary Research Institute, Izatnagar, India for funding of this work.
REFERENCES
- Ali, A., A.K. Abdel-Razek, R. Derar, H.A. Abdel-Rheem and S.H. Shehata, 2009. Forms of reproductive disorders in cattle and buffaloes in Middle Egypt. Reprod. Dom. Anim., 44: 580-586.
CrossRefDirect Link - Armstrong, D.G. and R. Webb, 1997. Ovarian follicular dominance: The role of intraovarian growth factors and novel proteins. Rev. Reprod., 2: 139-146.
CrossRefPubMedDirect Link - Basilico, C. and D. Moscatelli, 1992. The FGF family of growth factors and oncogenes. Adv. Cancer Res., 59: 115-165.
PubMed - Berisha, B., F. Sinowatz and D. Schams, 2004. Expression and localization of Fibroblast Growth Factor (FGF) family members during the final growth of bovine ovarian follicles. Mol. Reprod. Dev., 67: 162-171.
CrossRefDirect Link - Bottcher, R.T. and C. Niehrs, 2005. Fibroblast growth factor signaling during early vertebrate development. Endocr. Rev., 26: 63-77.
CrossRefDirect Link - Buratini, Jr. J., M.G.L. Pinto, A.C. Castilho, R.L. Amorim and I.C. Giometti et al., 2007. Expression and function of fibroblast growth factor 10 and its receptor, fibroblast growth factor receptor 2B, in bovine follicles. Biol. Reprod., 77: 743-750.
Direct Link - Caixeta, E.S., M.L. Sutton-McDowall, R.B. Gilchrist, J.G. Thompson and C.A. Price et al., 2013. Bone morphogenetic protein 15 and fibroblast growth factor 10 enhance cumulus expansion, glucose uptake and expression of genes in the ovulatory cascade during in vitro maturation of bovine cumulus-oocyte complexes. Reproduction, 146: 27-35.
CrossRefDirect Link - Chaves, R.N., I.B. Lima-Verde, J.J.H. Celestino, A.B.G. Duarte and A.M.C.V. Alves et al., 2010. Fibroblast growth factor-10 maintains the survival and promotes the growth of cultured goat preantral follicles. Domestic Anim. Endocrinol., 39: 249-258.
CrossRefDirect Link - Chouhan, V.S., R.P. Panda, V.P. Yadav, V. Babitha and F.A. Khan et al., 2013. Expression and localization of vascular endothelial growth factor and its receptors in the corpus luteum during oestrous cycle in water buffaloes (Bubalus bubalis). Reprod. Domest. Anim., 48: 810-818.
CrossRefDirect Link - El-Wishy, A.B., 2007. The postpartum buffalo: II. Acyclicity and anestrus. Anim. Reprod. Sci., 97: 216-236.
CrossRefDirect Link - Gasperin, B.G., R. Ferreira, M.T. Rovani, J.T. Santos, J. Buratini, C.A. Price and P.B.D. Goncalves, 2012. FGF10 inhibits dominant follicle growth and estradiol secretion in vivo in cattle. Reproduction, 143: 815-823.
CrossRefDirect Link - Itoh, N. and D.M. Ornitz, 2004. Evolution of the Fgf and Fgfr gene families. Trends Genet., 20: 563-569.
CrossRefDirect Link - Lavranos, T.C., H.F. Rodgers, I. Bertoncello and R.J. Rodgers, 1994. Anchorage-independent culture of bovine granulosa cells: The effects of basic fibroblast growth factor and dibutyryl cAMP on cell division and differentiation. Exp. Cell Res., 211: 245-251.
CrossRefDirect Link - Machado, M.F., V.M. Portela, C.A. Price, I.B. Costa, P. Ripamonte, R.L. Amorim and J. Buratini Jr., 2009. Regulation and action of fibroblast growth factor 17 in bovine follicles. J. Endocrinol., 202: 347-353.
CrossRefDirect Link - Matos, M.H.T., R. van den Hurk, I.B. Lima-Verde, M.C.A. Luque and K.D.B. Santos et al., 2007. Effects of fibroblast growth factor-2 on the in vitro culture of caprine preantral follicles. Cells Tissues Organs, 186: 112-120.
CrossRefDirect Link - McGee, E.A., S.Y. Chun, S. Lai, Y.E. He and A.J.W. Hsueh, 1999. Keratinocyte growth factor promotes the survival, growth and differentiation of preantral ovarian follicles. Fertil. Steril., 71: 732-738.
CrossRefDirect Link - Oron, G., B. Fisch, X.Y. Zhang, R. Gabbay-Benziv and G. Kessler-Icekson et al., 2012. Fibroblast growth factor 10 in human ovaries. Reprod. BioMed. Online, 25: 396-401.
CrossRefDirect Link - Oulion, S., S. Bertrand and H. Escriva, 2012. Evolution of the FGF gene family. Int. J. Evol. Biol.
CrossRefDirect Link - Parrott, J.A. and M.K. Skinner, 1997. Developmental and hormonal regulation of keratinocyte growth factor expression and action in the ovarian follicle. Endocrinology, 139: 228-235.
CrossRefDirect Link - Perera, B.M.A.O., 2008. Reproduction in domestic buffalo. Reprod. Domestic Anim., 43: 200-206.
CrossRefDirect Link - Pfaffl, M.W., 2001. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res., 29: e45-e45.
CrossRefPubMedDirect Link - Portela, V.M., M. Machado, J. Buratini Jr., G. Zamberlam, R.L. Amorim, P. Goncalves and C.A. Price, 2010. Expression and function of fibroblast growth factor 18 in the ovarian follicle in cattle. Biol. Reprod., 83: 339-346.
PubMedDirect Link - Powers, C.J., S.W. McLeskey and A. Wellstein, 2000. Fibroblast growth factors, their receptors and signaling. Endocr. Relat. Cancer, 7: 165-197.
CrossRefDirect Link - Roberts, R.D. and R.C. Ellis, 1999. Mitogenic effects of fibroblast growth factors on chicken granulosa and theca cells in vitro. Biol. Reprod., 61: 1387-1392.
PubMedDirect Link - Sarkar, M., S. Schilffarth, D. Schams, H.H.D. Meyer and B. Berisha, 2010. The expression of leptin and its receptor during different physiological stages in the bovine ovary. Mol. Reprod. Dev., 77: 174-181.
CrossRefDirect Link - Taniguchi, F., T. Harada, T. Iwabe, Y. Ohama, Y. Takenaka and N. Terakawa, 2008. Aberrant expression of keratinocyte growth factor receptor in ovarian surface epithelial cells of endometrioma. Fertil. Steril., 89: 478-480.
CrossRefDirect Link - Thisse, B. and C. Thisse, 2005. Functions and regulations of fibroblast growth factor signaling during embryonic development. Dev. Biol., 287: 390-402.
CrossRefPubMedDirect Link - Turner, N. and R. Grose, 2010. Fibroblast growth factor signalling: From development to cancer. Nat. Rev. Cancer, 10: 116-129.
CrossRefDirect Link - Vernon, R.K. and L.J. Spicer, 1994. Effects of basic fibroblast growth factor and heparin on follicle-stimulating hormone-induced steroidogenesis by bovine granulosa cells. J. Anim. Sci., 72: 2696-2702.
PubMedDirect Link - Ware, L.B. and M.A. Matthay, 2002. Keratinocyte and hepatocyte growth factors in the lung: Roles in lung development, inflammation and repair. Am. J. Physiol.-Lung Cell. Mol. Physiol., 282: L924-L940.
CrossRefDirect Link - Webb, R., B. Nicholas, J.G. Gong, B.K. Campbell, C.G. Gutierrez, H.A. Garverick and D.G. Armstrong, 2003. Mechanisms regulating follicular development and selection of the dominant follicle. Reproduction, 61: 71-90.
PubMedDirect Link - Zhang, K., P.J. Hansen and A.D. Ealy, 2010. Fibroblast growth factor 10 enhances bovine oocyte maturation and developmental competence in vitro. Reproduction, 140: 815-826.
CrossRefDirect Link - Zicarelli, L., 2010. Enhancing reproductive performance in domestic dairy water buffalo (Bubalus bubalis). Soc. Reprod. Fertil. Suppl., 67: 443-455.
PubMedDirect Link