
A.S. Ahmed
Department of Animal and Fish Production, College of Agriculture and Food Science, King Faisal University, Saudi Arabia
Asian Journal of Animal and Veterinary Advances
Year: 2015 | Volume: 10 | Issue: 1 | Page No.: 35-42







ABSTRACT
This study was conducted to assess the effect of genetic crossing using four chicken lines Hajar1 line, Hajar2 line, Hisex sires×Hajar1 dams cross (HiH1) and Hisex sires×Hajar2 dams cross (HiH2) on the maternal immunity against Newcastle Disease (ND) of chicks for the first 14 days of age. Each line consisted of 100 unvaccinated chicks represented both sexes equally in five replicates in a caging house system. Feed and water were provided ad libitum. Serum samples were collected at 3, 7, 10 and 14 days of age for ND titration. Spleen and bursa weight were evaluated at 14 days of age. The results indicated comparable ND antibody levels between all lines at 3 days of age. Cross lines chicks showed significantly low ND antibody level by 7, 10 and 14 days of age compared to local lines. Sex of the birds did not participate as a major factor that affect ND antibody level or glands weight. Bursa weight did not affected by genetic line while spleen weight demonstrated trivial change due to the genetic line effect. Although the literature provided little attention to the role of male partner in the maternal immunity efficiency, our results indicated the possible effect of male parents in modulating the maternal antibody efficiency to ND in their offspring. In addition, the genetic line and the male parent participation in maternal antibodies degradation process over the first 14 days of age should be considered.
PDF Abstract XML References Citation
Received: November 26, 2014;
Accepted: January 06, 2015;
Published: February 13, 2015
How to cite this article
A.S. Ahmed, 2015. Assessment of Maternal Immunity against Newcastle Disease in Offspring Chicks
as Affected by Parents Genetic Crossing. Asian Journal of Animal and Veterinary Advances, 10: 35-42.
DOI: 10.3923/ajava.2015.35.42
URL: https://scialert.net/abstract/?doi=ajava.2015.35.42
DOI: 10.3923/ajava.2015.35.42
URL: https://scialert.net/abstract/?doi=ajava.2015.35.42
INTRODUCTION
Maternal transfer of antibodies via egg yolk is very important to protect newly hatched chicks from common pathogens before their immune system mature (Hasselquist and Nilsson, 2009). By expected transfer percentage for IgY of 30% to chicks circulation, the maternal antibody transfer from dams to their chicks could be a good strategy to protect them in the first few days of age against specific pathogens (Hamal et al., 2006). Vaccination programs are highly varied based on different factors, maternal immunity is one of those important factors, for instance live vaccines are susceptible to the existing maternal antibody where, in the presence of high level maternal antibody in the first two weeks of age the vaccine may subjected to vaccine neutralization (Marangon and Busani, 2006). Vaccination while having a high level of maternal antibodies leads to vaccination failure and neutralization of living vaccine (Al-Natour et al., 2004). The level of maternal antibody decay and its half life time is an important information for designing a suitable vaccination program, the estimated half life time of ND maternal antibody level is 6.3 days (Gharaibeh and Mahmoud, 2013). Maternally transmitted immunoglobulins are catabolized by offspring for 14 days after hatching (Siwek and Knoll, 2005). Using broilers Jalil et al. (2009) studied the decline of maternal antibody level of NDV, they reported that the decline of maternal antibody titer in chicks was about half every 7 days. In another study, Hamal et al. (2006) reported that the transferred anti Newcastle disease virus antibody recorded the highest level in chicks plasma at 3 days of age then decreased considerably by the 7th day of age to vanished at 14 days of age in meat lines commercial chicken. Effect of the genetic line on maternal immunity has been demonstrated by Abdel-Moneim and Abdel-Gawad (2006), they explained that the variation in responsiveness to infectious diseases between native and crossbred chicken in response to IBDV depend partially on the inherited maternally derived antibodies in day old chicks. Genes of the neonates influence their innate defense system, the genes of the female help through maternal antibodies in the early days of chicks life (Siwek and Knoll, 2005). Differences in antibody level against NDV between meat lines of chicken were reported by Hamal et al. (2006) where, line 1 had higher antibody level to NDV than line 2. Not only the antibody level differences have been reported but also differences in Bursa and spleen weight. In a study of two broiler breeder strains Leandro et al. (2011) reported that breeder strain affected maternal antibody transfer to progeny and embryo development, they added that there are difference among the strains in bursa of fabricius and spleen growth. The spleen and bursa weight are affected by the genetic pool of the bird, the presence of the naked neck gene significantly decreased their weight compared to normally feathered birds while, the presence of the F gene increased the relative weight of bursa (Fathi et al., 2014). In the current study four chicken lines composed of two local lines and their crosses with commercial Hisex breeder sires were subjected to evaluation for maternal immunity against Newcastle disease through the first 14 days of age without exposure to any vaccination. This research study tried to assess the participation of the genetic pool modification via crossing approach with commercial line sires in maternal immunity against ND in early ages of the chicks. Moreover, to investigate the effect of sex of the birds in such process, we hypothesized that the maternal immune response to ND in the newly hatched chicks through the first couple weeks of age could be affected by female parents crossing with commercial sires due to the modulation of the host chicks genetic pool. In addition, offspring sex effect could interfere with the level of response.
MATERIALS AND METHODS
Parent lines and obtained chicks: The chicks used in the present study were obtained from four genetic line parents at the poultry experimental unit of King Faisal University Experimental Station as follow. Hajar1 line, Hajar2 line, Hisex siresxHajar1 dams cross (HiH1) and Hisex siresxHajar2 dams cross (HiH2). The Hajar1 and Hajar2 lines are local chicken lines, both lines are locally characterized lines for their phenotypic characters (Ahmed and Alabbad, 2014). In addition some genetic parameters of both lines have been reported (Alabbad, 2014). There were noticeable diversity in some productive and physiological parameters between the two lines (Ahmed et al., 2014). Forty dams and 4 sires were used in each line to produce the current experimental chicks. All parents received an identical vaccination program and same management treatments since one-day-old. At 45 weeks of age all dams were evaluated for ND antibody titer to ensure that there were not significant differences in dams serum antibody level. Average antibody titers against ND at 45 weeks of age were 8.21, 8.69, 8.43 and 8.52 for Hajar1, Hajar2, HiH1 and HiH2, respectively no significant differences were detected among lines. Each parent line chickens were placed randomly in four floor bens each of 10 dams and one sire in an open house system with ventilation fans, feeders, drinkers, wood shave litter and trap nests. Parents were exposed to 17 h day-1 of light during the experimental period. Feed and water were available ad libitum. Birds were fed a commercial layer diet. Fertile eggs were collected twice daily for 10 days and kept under 10°C until incubation and then eggs were placed into incubator. One hatch was obtained from all lines.
Experimental chicks: After hatch, one hundred chicks were obtained from each line, all chicks were subjected to vent sexing, wing banded and housed in five replicates per each line in a cage system for brooding. Feed and water were provided ad libitum. Cages were placed in a closed house system considering the optimum chicks brooding criteria. Feed and water were provided ad libitum. All chicks were left without any vaccination over the experimental period with daily close monitoring for chicks.
Parameters and data collection: All chicks were bled via the jugular vein at 3, 7, 10 and 14 days of age using a 0.5 mL insulin syringe with a 28.5-gauge needle. The serum samples were collected after centrifugation (3000 rpm, 3 min), stored at -20°C. Serum samples were collected to assess the maternal antibody against ND. Antibody titer was assayed using microtitre Hemagglutination Inhibition (HI) test as described by OIE (2009). At 14 days of age, after blood sampling, all birds were weighted individually and then 50% of all chicks in each line represented equally male and female birds were scarified, then the bursas of Fabricius and spleens were removed and their weight recorded individually. The research complied with King Faisal University animal care guidelines.
Statistical analysis: The data from this study were subjected to a two-way analysis of variance for the effect of line, sex and their interactions. Data were analyzed using the general linear model procedure of JMP IN 5.1 software (Sall et al., 2005). Statistical significance was considered as p≤0.05 throughout the paper.
RESULTS
Figure 1 demonstrates the antibody titer against ND of different chicks lines at 3, 7, 10 and 14 days of age. The means among all lines were comparable at 3rd day of age indicating that genetic line did not affect ND antibody titer at the very early age. Cross line HiH2 chicks recorded a significant (p<0.05) low ND antibody level compared to Hajar1 and Hajar2 lines for 7, 10 and 14 days of age. At 14 days of age both cross lines recorded the lowest significant (p<0.05) ND antibody titer. The ND antibody titer results of the effect of line-sex interactions in the different lines demonstrated in Table 1.
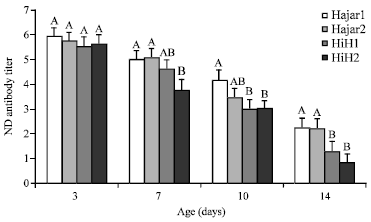 | |
| Fig. 1: | Effect of genetic line on maternal antibody titer against Newcastle Disease (ND) of chicks at 3, 7, 10 and 14 days of age, HiH1 and HiH2 are Hisex siresxHajar1 dams and Hisex siresxHajar2 dams crosses, respectively, A,BValues within a day with different superscript differ significantly (p≤0.05) |
Line-sex interaction did not affect the level of ND antibody titer at 3rd day of age. At 7 days of age female chicks of HiH2 cross line recorded a significant (p<0.05) lower ND antibody titer compared to Hajar1 and Hajar2 females while, males of all lines were not significantly different from each other neither at 7 days of age nor at 10 days of age. At 14 days of age male chicks of HiH2 cross line recorded significant (p<0.05) lower ND antibody titer compared to what Hajar1 and Hajar2 male birds recorded. No significant differences in ND antibody titer were recorded among female birds of all lines at 14 days of age. The results did not show any significant (p<0.05) differences between male and female chicks within the same genetic line at any of the measuring points. Bursa weight at 14 days of age did not affected by the genetic line as shown in Fig. 2. Mean spleens weight was significantly (p<0.05) higher in HiH2 cross line birds compared to Hajar2 birds.
| Table 1: | Effect of genetic line-sex interaction on maternal antibody titer against Newcastle Disease (ND) of chicks at 3, 7, 10 and 14 days of age |
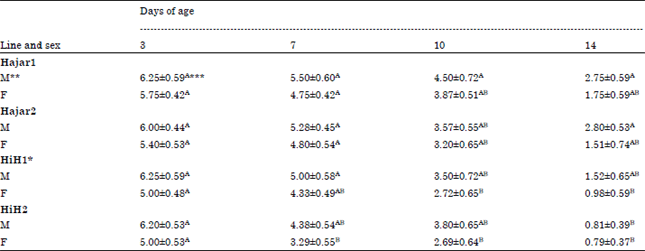 | |
| *HiH1 and HiH2 are Hisex siresxHajar1 dams and Hisex siresxHajar2 dams crosses, respectively, **M and F are male and female birds, respectively, ***A,BValues within a day with different superscript differ significantly (p≤0.05), The titer values are log2 of the reciprocal dilution, Sample size was minimum 96 and maximum 100 birds for each line at any measuring point | |
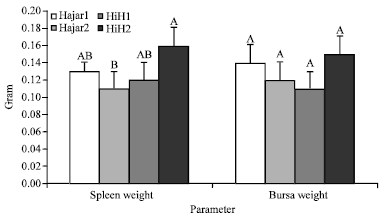 | |
| Fig. 2: | Effect of genetic line on spleen and bursa of fabricius weight of chicks at 14 days of age, HiH1 and HiH2 are Hisex siresxHajar1 dams and Hisex siresxHajar2 dams crosses, respectively, A,BValues within a parameter with different superscript differ significantly (p≤0.05) |
| Table 2: | Effect of genetic line-sex interaction on spleen and bursa of fabricius weight of chicks at 14 days of age |
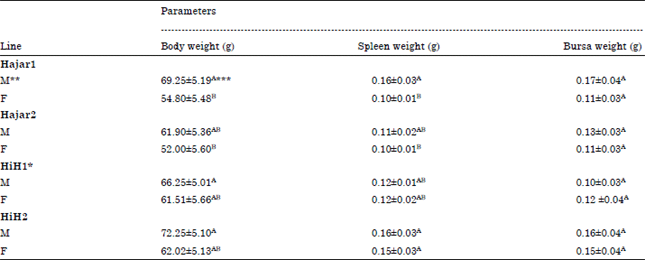 | |
| *HiH1 and HiH2 are Hisex siresxHajar1 dams and Hisex siresxHajar2 dams crosses, respectively, **M and F are male and female birds, respectively, ***A,BValues within a parameter with different superscript differ significantly (p≤0.05); Sample size was minimum 93 and maximum 99 birds in each line for body weight, while minimum 47 and maximum 50 birds in each line for spleen and bursa weight | |
However, all other lines except Hajar2 line did not show significant differences in birds spleen weight (Fig. 2). Male birds of all genetic lines had comparable body weight at 14 days of age and the same tendency was recorded for female birds too (Table 2). There were not any significant differences between both sexes in body weight at 14 days of age within each genetic lines except for Hajar1 line where, male birds recorded significant higher body weight than female birds (Table 2). Bursa weight of all birds did not affected by line-sex interaction whereas, spleen weight partially affected by line-sex interaction (Table 2). Male chicks of all lines recorded comparable spleen weight while, female chicks of HiH2 cross line recorded a higher spleen weight compared to Hajar1 and Hajar2 female birds. There were not any significant differences in spleen weight between male and female chicks within each line except for Hajar1 line where Hajar1 male birds had higher spleen weight than female birds of both Hajar1 and Hajar2 lines.
DISCUSSION
The current results pointed out to the potential role of sires in modulating the ability of the offspring to maintain maternal antibody levels against ND. This is important since most of the research studies focuses only on the maternal role in providing maternal antibody level that passively transfer to the chicks regardless father effect (Gharaibeh et al., 2008; Jalil et al., 2009; Addison et al., 2010; Maas et al., 2011; Ahmed, 2011; Gharaibeh and Mahmoud, 2013). The current research study suggested that the host chicks play an important role in managing the period required to decay the maternal antibody. This process inside the host chick affected by its genetic makeup which is inherited from both parents. The effect of genetic line on maternal antibody level in the late measuring points as shown in the current results suggested that the change in the genetic makeup through crossing may leads to a significant decrease in the maternal antibody level against ND at 14 days of age. Effect of genetic line has been previously reported by Leandro et al. (2011) using different commercial broiler breeder lines, they stated that the breeder line affected the transfer of maternal antibody and the development of bursa and spleen. The male parents effect have been studied in other avian species where, Saino et al. (2002) demonstrated that the allocation of maternal immunity in eggs partially affected by the quality of fathers in barn swallows birds. Not only the quantity of maternal antibody transfer is affected by genetic selection but also how long these antibodies maintained is under genetic control (Boa-Amponsem et al., 1997; Grindstaff et al., 2003).
The results demonstrated that sex of the chicks did not highly affect maternal antibody degradation, while genetic line factor was superior. Both sexes of HiH2 cross line chicks and HiH1 female chicks recorded low ND titer values especially at 14 days of age although their comparable body weight with all other lines as demonstrated in Table 2. The trivial effect of the chicken offspring sex on the maternal antibody level in the current study is disagreed with some previous reports which indicated a variation in egg components related to offspring sex in some other avian species (Cordero et al., 2001; Anderson et al., 1997). In addition, Saino et al. (2003) reported that barn swallow mothers allocate more amounts of maternal antibodies to eggs with a daughter than a son. No significant effect has been shown for genetic line or sex within line on chicks bursa weight, in addition spleen weight suggested to be related to the body weight than the genetic line or bird’s sex.
The differences in maternal antibody kinetic over the first 14 days of age depend on many factors included but not limited to the initial maternal antibody level, where the amount of IgY absorbed by the embryo is a small proportion, about 10%, of that deposited in the egg yolk (Davison et al., 2008). This proportion suggested to be affected by the embryo physiological capacity which is under the genetic control that inherited from both parents, previous reports demonstrated the effect of the breed on maternal antibody transfer to chicks (Bumstead et al., 1991; Hamal et al., 2006). Maternal antibodies degradation affected by different factors including protein metabolism (Saino et al., 2002) the protein metabolic rate is partially under genetic control that could play an important role in decay of maternal antibodies.
CONCLUSION
The current research study indicated the possible effect of male parents to modulate the maternal antibody level of their offspring. The results pointed out to the importance of considering the genetic line and the male parent participation in the process of maintaining maternal antibody level over the first 14 days of age. Further research is needed to understanding the specific role of male parent in such process.
ACKNOWLEDGMENTS
The author gratefully acknowledge deanship of scientific research at King Faisal University (KFU) for financial support of the current research work. Thank also extended to Agriculture Research Station at KFU for their support in conducting this research. The author also acknowledges the technical assistance of Abdullah Alnazr, AbdulRazak Al-Abbad, Gaafar Al Hamada, Zafer Hakami and all the staff at the poultry research unit.
REFERENCES
- Abdel-Moneim, A.S. and M.M.A. Abdel-Gawad, 2006. Genetic variations in maternal transfer and immune responsiveness to infectious bursal disease virus. Vet. Microbiol., 114: 16-24.
CrossRefDirect Link - Addison, B., R.E. Ricklefs and K.C. Klasing, 2010. Do maternally derived antibodies and early immune experience shape the adult immune response? Funct. Ecol., 24: 824-829.
CrossRefDirect Link - Ahmed, A.S., 2011. Antibody response in early hatched chicks as influenced by vaccination and different maternal antibody levels. Egypt. J. Anim. Prod., 48: 81-89.
Direct Link - Ahmed, A.S. and A.H. Alabbad, 2014. First Report about Growth, Partial Record Egg Production and Morphological Characters of a Newly Characterized Native Saudi Chicken Lines Hajar 1 and Hajar 2. In: Biotechnology and Conservation of Species from Arid Regions, Farooq, S.A., R.M.M. Abed and S. Baqir (Eds.). Vol. 1, Chapter 10, Nova Science Publisher Inc., USA., ISBN-13: 978-1631178696, pp: 105-114.
- Al-Natour, M.Q., L.A. Ward, Y.M. Saif, B. Stewart-Brown and L.D. Keck, 2004. Effect of different levels of maternally derived antibodies on protection against infectious bursal disease virus. Avian Dis., 48: 177-182.
CrossRefPubMedDirect Link - Anderson, D.J., J. Reeve and D.M. Bird, 1997. Sexually dimorphic eggs, nestling growth and sibling competition in American Kestrels Falco sparverius Funct. Ecol., 11: 331-335.
CrossRef - Cordero, P.J., J. Vinuela, J.M. Aparicio and J.P. Veiga, 2001. Seasonal variation in sex ratio and sexual egg dimorphism favouring daughters in first clutches of the spotless starling. J. Evol. Biol., 14: 829-834.
CrossRefDirect Link - Fathi, M.M., A. Galal, S. El-Safty and M. Mahrous, 2014. Naked neck and frizzle genes for improving chickens raised under high ambient temperature: II. Blood parameters and immunity. World's Poult. Sci. J., 70: 165-172.
CrossRefDirect Link - Gharaibeh, S., K. Mahmoud and M. Al-Natour, 2008. Field evaluation of maternal antibody transfer to a group of pathogens in meat-type chickens. Poult. Sci., 87: 1550-1555.
CrossRefDirect Link - Gharaibeh, S. and K. Mahmoud, 2013. Decay of maternal antibodies in broiler chickens. Poult. Sci., 92: 2333-2336.
CrossRefDirect Link - Grindstaff, J.L., E.D. Brodie and E.D. Ketterson, 2003. Immune function across generations: Integrating mechanism and evolutionary process in maternal antibody transmission. Proc. R. Soc. London Ser. B: Biol. Sci., 270: 2309-2319.
CrossRef - Hamal, K.R., S.C. Burgess, I.Y. Pevzner and G.F. Erf, 2006. Maternal antibody transfer from dams to their egg yolks, egg whites and chicks in meat lines of chickens. Poult. Sci., 85: 1364-1372.
CrossRefDirect Link - Hasselquist, D. and J.K. Nilsson, 2009. Maternal transfer of antibodies in vertebrates: Trans-generational effects on offspring immunity. Philos. Trans. R. Soc. London B: Biol. Sci., 364: 51-60.
CrossRefPubMedDirect Link - Jalil, M.A., M.A. Samad and M.T. Islam, 2009. Evaluation of maternally derived antibodies against Newcastle disease virus and its effect on vaccination in broiler chicks. Bangladesh J. Vet. Med., 7: 296-302.
CrossRefDirect Link - Leandro, N.M., R. Ali, M. Koci, V. Moraes and P.E. Eusebio-Balcazar et al., 2011. Maternal antibody transfer to broiler progeny varies among strains and is affected by grain source and cage density. Poult. Sci., 90: 2730-2739.
CrossRefDirect Link - Marangon, S. and L. Busani, 2007. The use of vaccination in poultry production. Rev. Sci. Technol. Off. Int. Epiz., 26: 265-274.
PubMedDirect Link - Maas, R., S. Rosema, D. van Zoelen and S. Venema, 2011. Maternal immunity against avian influenza H5N1 in chickens: Limited protection and interference with vaccine efficacy. Avian Pathol., 40: 87-92.
CrossRefDirect Link - Saino, N., R.P. Ferrari, R. Martinelli, M. Romano, D. Rubolini and A.P. Moller, 2002. Early maternal effects mediated by immunity depend on sexual ornamentation of the male partner. Proc. R. Soc. B: Biol. Sci., 269: 1005-1009.
CrossRefPubMedDirect Link - Saino, N., M. Romano, R.P. Ferrari, R. Martinelli and A.P. Moller, 2003. Maternal antibodies but not carotenoids in barn swallow eggs covary with embryo sex. J. Evol. Biol., 16: 516-522.
CrossRefDirect Link - Siwek, M. and E.F. Knoll, 2005. Genetic aspects of biological processes underlying the defense system in the neonate. Folia Biologica, 53: 39-43.
Direct Link