
Meiling Zhang
Anhui Provincial Laboratory of Local Livestock and Poultry Genetical Resource Conservation and Breeding, College of Animal Science and Technology, Anhui Agricultural University, Hefei City, Anhui Province 230036, China
Tao Gui
Anhui Provincial Laboratory of Local Livestock and Poultry Genetical Resource Conservation and Breeding, College of Animal Science and Technology, Anhui Agricultural University, Hefei City, Anhui Province 230036, China
Yunsheng Li
Anhui Provincial Laboratory of Local Livestock and Poultry Genetical Resource Conservation and Breeding, College of Animal Science and Technology, Anhui Agricultural University, Hefei City, Anhui Province 230036, China
Chao Wei
Anhui Provincial Laboratory of Local Livestock and Poultry Genetical Resource Conservation and Breeding, College of Animal Science and Technology, Anhui Agricultural University, Hefei City, Anhui Province 230036, China
Nian Zhan
Anhui Provincial Laboratory of Local Livestock and Poultry Genetical Resource Conservation and Breeding, College of Animal Science and Technology, Anhui Agricultural University, Hefei City, Anhui Province 230036, China
Zhen Chen
Anhui Provincial Laboratory of Local Livestock and Poultry Genetical Resource Conservation and Breeding, College of Animal Science and Technology, Anhui Agricultural University, Hefei City, Anhui Province 230036, China
Yuanliang Zhang
Anhui Provincial Laboratory of Local Livestock and Poultry Genetical Resource Conservation and Breeding, College of Animal Science and Technology, Anhui Agricultural University, Hefei City, Anhui Province 230036, China
Ya Liu
Anhui Provincial Laboratory of Local Livestock and Poultry Genetical Resource Conservation and Breeding, College of Animal Science and Technology, Anhui Agricultural University, Hefei City, Anhui Province 230036, China
Qing Jia
Anhui Provincial Laboratory of Local Livestock and Poultry Genetical Resource Conservation and Breeding, College of Animal Science and Technology, Anhui Agricultural University, Hefei City, Anhui Province 230036, China
Jianping Ding
Anhui Provincial Laboratory of Local Livestock and Poultry Genetical Resource Conservation and Breeding, College of Animal Science and Technology, Anhui Agricultural University, Hefei City, Anhui Province 230036, China
Xiaorong Zhang
Anhui Provincial Laboratory of Local Livestock and Poultry Genetical Resource Conservation and Breeding, College of Animal Science and Technology, Anhui Agricultural University, Hefei City, Anhui Province 230036, China
Yunhai Zhang
Anhui Provincial Laboratory of Local Livestock and Poultry Genetical Resource Conservation and Breeding, College of Animal Science and Technology, Anhui Agricultural University, Hefei City, Anhui Province 230036, China
Asian Journal of Animal and Veterinary Advances
Year: 2013 | Volume: 8 | Issue: 5 | Page No.: 713-722







ABSTRACT
This study was conducted to explore efficient and cost-effective protocols for isolation and culture of mammary epithelial cells from goats during gestation period (non-lactating). Tissues collected from the mammary glands of a Huanghuai white goat, at gestation day 60 were cultured in the presence of fetal bovine serum, insulin-transferrin-selenium and epidermal growth factor. Cell lines were purified using differential enzyme digestion, high density cultivation and continuous passaging. And subsequently characterized by immunofluorescence staining, cell growth curve, oil red O staining and karyotype analysis. In conclusion, a goat mammary epithelial cell line was successfully established and shown to exhibit normal conformation, growth characteristics and lactation function by using improved sample collection and in vitro culture methods. This study provides a new and efficient cell culture protocol, which may facilitate research into the development and function of goat mammary glands and the development of bioreactors.
PDF Abstract XML References Citation
Received: March 23, 2013;
Accepted: June 17, 2013;
Published: August 05, 2013
How to cite this article
Meiling Zhang, Tao Gui, Yunsheng Li, Chao Wei, Nian Zhan, Zhen Chen, Yuanliang Zhang, Ya Liu, Qing Jia, Jianping Ding, Xiaorong Zhang and Yunhai Zhang, 2013. Establishment and Characterization of a Huanghuai White Goat Mammary Gland Epithelial Cell Line Derived During the Gestational Period. Asian Journal of Animal and Veterinary Advances, 8: 713-722.
DOI: 10.3923/ajava.2013.713.722
URL: https://scialert.net/abstract/?doi=ajava.2013.713.722
DOI: 10.3923/ajava.2013.713.722
URL: https://scialert.net/abstract/?doi=ajava.2013.713.722
INTRODUCTION
Mammary gland is considered to be one of mysterious organ of the whole body. Because they can repeatedly undergo pubertal growth, functional differentiation and involution (Warri et al., 2007; Macias and Hinck, 2012). Mammary epithelial cells, which are the major components of mammary gland tissue, function in the synthesis and secretion of milk. Thus, Samiec and Skrzyszowska (2011) have proposed that in vitro culture of these cells provides an important resource for research. First, cultured mammary epithelial cells mimic mammary gland development and lactation at the cellular level which mainly provide experimental tools for investigation of mammary gland regulatory mechanisms. Second, commercialized mice mammary gland cancer cells(C-127) in Yi and Li (2010) research and human breast cancer cells (MCF-7) (Zhou et al., 2011) have been used to investigate rational and efficient approaches to the construction of mammary gland-specific expression vectors. However, whether mouse and human cell lines reflect the function of mammary gland-specific expression in large animals remains to be elucidated. However, the specific functions and signaling pathways are retained in primary cultures of mammary epithelial cells derived from large animals. In contrast, mammary epithelial cells of large animals such as cattle and sheep correlate closely with the expression levels of mammary gland-specific expression vectors in the mammary glands of large livestock (He et al., 2009). More importantly, sustained cultures of mammary epithelial cells that exhibit normal differentiation could be used as donor cells for nuclear transfer of exogenous proteins in the generation of transgenic animals. In this way, the success rate of mammary gland bioreactor generation could be greatly increased.
Mammary gland epithelial cells are highly differentiated. The optimal conditions and supplemented factors for culturing these cells in vitro are a subject for continuous investigation (Duo et al., 2006). Mammary epithelial cell lines have been successfully established from humans (Colomb et al., 1991), mice (Danielson et al., 1984), dairy cattle (Hu et al., 2009), sheep (Ilan et al., 1998) and goats (Pantschenko et al., 2000). However, these cell lines have rarely proved to be suitable for functional analyses of mammary gland-specific expression vectors due to the complexity of primary mammary epithelial cell culture preparation in large animals and difficulties associated with differential culture in vitro. Currently, no consistent protocol has been established that defines optimal tissue sources and cell culture conditions for collagenase pre-treatment and the combination of exogenous growth factors used in the medium of mammary epithelial cell lines. Previously reported methods vary in terms of the constituents of the culture medium and include many expensive hormones (such as prolactin, progesterone, hydrocortisone and estradiol) and cytokines (such as epidermal growth factor, insulin-like growth factor-1 and insulin). Therefore, the currently available protocols for the establishment of mammary epithelial cell lines are highly expensive.
Protocols describing the establishment of goat mammary epithelial cell lines reported to date predominantly use goat mammary gland tissue obtained during the lactation period as the primary resource (Ilan et al., 1998; Pantschenko et al., 2000) with few reports of tissue obtained during the earlier gestation period. Although a greater number of acini are present in the gestation period compared with the middle and later periods of lactation, the rate of cell proliferation is higher than that during the lactation period due to the rapid development of the mammary gland being at this stage. Therefore, this study aimed to separate mammary epithelial cells form mammary gland tissues obtained during the gestation period prior to lactation to optimize the isolation methods and conditions traditionally used for the culture of mammary epithelial cells.
MATERIALS AND METHODS
Reagents: DMEM/F-12, Dulbecco’s Phosphate Buffered Saline (DPBS) and Fetal Bovine Serum (FBS) were purchased from Hyclone (Logan, UT). Trypsin (0.25%) with EDTA and Insulin-transferrin-selenium (ITS) were from Invitrogen (Shanghai, China). Epidermal Growth Factor (EGF) was from Sigma (St. Louis, MO). β-Casein, Vimentin), Cytokeratin-18 (CK18), epithelial membrane antigen (EMA) antibodies, rabbit IgG SABC-FITC and mouse IgG SABC-Cy3 kits were from Wuhan Boster Bio-engineering Co. Ltd. (China). All other chemicals used in this study were from Solarbio (Beijing, China).
Cell isolation, culture and cryopreservation: Pregnant Huanghuai white goats (60 days of gestation) were kindly provided by Hefei Boda Livestock Technology Development Co. Ltd., (Hefei, China). Our investigations were approved and conducted according to the strict guidance of the Local Ethics Committee of Society for the Protection of Animals at the Anhui Agriculture University. The tissues were harvested from mammary glands under sterile conditions and cut into 1 mm explant cubes. These were then incubated in normal saline containing 400 IU mL-1 penicillin and 400 μg mL-1 streptomycin followed by washing with DPBS. Explants were transferred into 10 cm dish and humidified by serum. The dish was inverted and cultured at 37°C under an atmosphere of 5% CO2 for 10 h. Then, Culture medium (DMEM/F-12, 10% FBS, 1% ITS, 10 ng mL-1 EGF) was added to the upright dishes and made up to 5 mL At 80% confluence, the cells were trypsinized and cryopreserved in cryoprotectant (10% DMSO, 30% serum and 60% DMEM/F-12).
Purification and subculture of mammary epithelial cells: In order to remove fiber-like cells, the cells at 80% confluence were washed with DPBS twice. The cells were then incubated with trypsin-0.02% EDTA (2-3 min). Fibroblasts were detached first and removed. The remaining cells were further incubated with trypsin (with 0.02% EDTA-2Na) until all the cells were almost detached. Medium containing FBS was added to stop trypsinization and the cells were harvested after centrifugation. A pure population of Goat Mammary Epithelial Cells (GMEC) was expected obtained after continuous passages.
Growth curves: Cells (passage 5) were seeded (4x104/well) in 12-well plates and cultured in GMEC complete medium (DMEM/F-12, 10% FBS, 1% ITS, 10 ng mL-1 EGF) or GMEC medium without EGF (DMEM/F-12+10% FBS+1% ITS). Cell number and viability were determined daily in quadruplicate wells for eight days using the trypan blue exclusion method and growth curves over time were generated.
Karyotyping analysis: Karyotyping analysis was performed as described previously (Tai et al., 2011). Briefly, GMECs at passages 5, 10 and 25 were harvested at 80% confluence. Cells were then incubated with colchicine (0.2 μg mL-1) for 4 h, released by trypsinization and fixed with ice-cold methanol before being spotted onto ice-cold glass slides. The slides were dried and stained with Giemsa solution for 10 min before being washed with distilled water and dried at room temperature. Chromosomes were visualized and detected with a phase-contrast microscope (Olympus IX71, Japan) and analyzed with Image J software (NIH, USA).
Immunostaining: Immunofluorescence staining was performed using SABC kits according to the instructions provided by the manufacturer instruction. Briefly, GMECs (passage 5) grown on glass coverslips in 12-well plates were fixed in 10% acetic acid, 50% ethanol for 40 min at 4°C and washed with phosphate buffered saline (PBS). Primary antibodies (β-Casein, Vimentin, CK18, EM; diluted 1:50) were applied for 1 h at 37°C followed by incubation with FITC- or Cy3-labeled secondary antibodies (dilution, 1:100) and 4',6-diamidino-2-phenylindole (DAPI). Immunofluorescence images were recorded with an Olympus AX70 epifluorescence microscope (Olympus, Tokyo, Japan).
Oil red O staining: GMECs at passages 2, 4, 9 and 16 were fixed for 15 min in 4% formaldehyde, washed with DPBS and stained with Oil red O for 10 min at room temperature. The stained cells were visualized under an inverted fluorescence microscope.
Statistical analysis: All experiments were performed in quadruplicate (minimum). SPSS 17.0 statistical software was used for the statistical analysis. Statistical evaluation of the raw data was performed using chi-square tests. Values of p<0.05 were considered statistically significant.
RESULTS
Primary culture, purification and subculture of mammary epithelial cells: Extravasation of a small number of cells from explant tissues in culture was observed at day 5, while numerous cells were observed at day 8 (Fig. 1a). Subsequently, cultures consisted of a mixed population of fibroblast-like cells and mammary epithelial cells, with obvious boundaries (Fig. 1b). After two to four passages, a higher purity of mammary epithelial cells was obtained. Purified fibroblasts exhibited a typical long spindle shape (Fig. 1c), which was transformed to a uniform short spindle shape after purification (Fig. 1d). During the early stages of contact inhibition, the cells accumulated in single layers to form a round and flat cake (Fig. 1e), while at later stages, net-like structures (Fig. 1f) or domed ones, presenting a papillary shape (Fig. 1g). After passage 8, more pebble-shaped cells were observed and fewer short spindle shaped cells (Fig. 1h). After cryopreservation, the thawed cells grew well in culture with low levels of cell death. High rates of proliferation and normal growth patterns were observed in purified cells up to passage 30 (Fig. 1i).
 | |
| Fig. 1(a-i): | Morphological observation of Huanghuai white goat mammary epithelial cells, (a) Mammary epithelial cells cultured from mammary tissue explants (day 4) (40x), (b) Mixed cultures of mammary epithelial cells with clear boundaries (arrow) (40x), (c) Purification of fibroblasts from mammary epithelial cells (40x), (d) Purified mammary gland epithelial cells; short spindle form (100x), (e) Purified mammary epithelial cells, round, flat form (100x), (f) Net-like structure formed after attachment-inhibited growth (arrow) (100x), (g) Papillary structure formed after attachment-inhibited growth (arrow) (40x), (h) The main formation; pebble-shaped, short spindle (100x) and (i) Mammary epithelial cells at passage 25 (100x) |
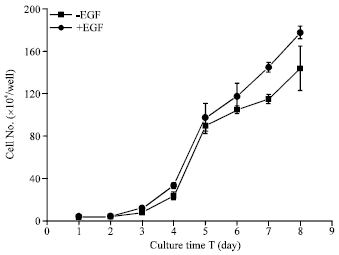 | |
| Fig. 2: | Huanghuai white goat mammary gland epithelial cell growth curve |
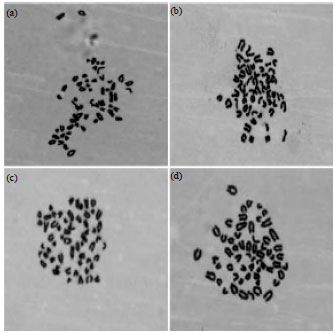 | |
| Fig. 3(a-d): | Karyotypes of Huanghuai white goat mammary gland epithelial cell lines (1,000x magnification), Chromosome number, 60, (a) Passage 5, (b) Passage 10, (c) Passage 15 and (d) Passage 20 |
Cell growth curves: Low levels of proliferation were detected in mammary epithelial cells during the first four days of in vitro culture, followed by a logarithmic increase from the day 4 to day 6. From day 6 to day 8, proliferation continued at a decreased rate. Contact inhibition of growth was observed at day 8 in 12-well plates. Growth curves of mammary epithelial cells showed that GMEC proliferation was enhanced in the presence of EGF (Fig. 2).
Chromosome karyotype: Analysis of GMECs at passages 5, 10 15 and 20 (Fig. 3) revealed the presence of 60 chromosomes, which was consistent with the normal goat karyotype. This demonstrated that the number of chromosomes in mammary epithelial cells was not altered during the course of the subculturing process.
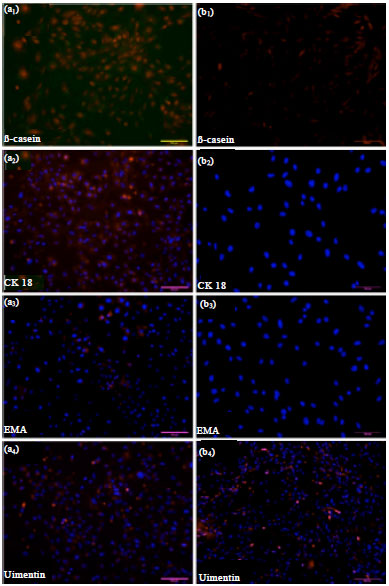 | |
| Fig. 4(a-b): | (a) Immunofluorescent staining of the mammary gland epithelial cell monolayer (Passage 5), a1-a4 (200x magnification) Positive staining of β-Casein/ CK18/EMA/Vimentin shown with merged PI/DAPI nuclear staining (b) Immunofluorescent staining of goat fibroblast cells, b1-b3 (200x magnification), b4 (100x magnification) Staining of β-Casein/CK18/EMA/Vimentin shown with merged PI/DAPI nuclear staining |
Thus, the genetic material of these cells was shown to be relatively stable and the GMEC line was established without transformation.
Immunofluorescence staining: Immunofluorescent staining confirmed expression of cytokeratin-18, EMA, vimentin and β-casein after mammary epithelial cell purification. In the negative group (fiber-like cells), cytokeratin-18, EMA and β-casein were not expressed, while expression of vimentin was strongly positive (Fig. 4).
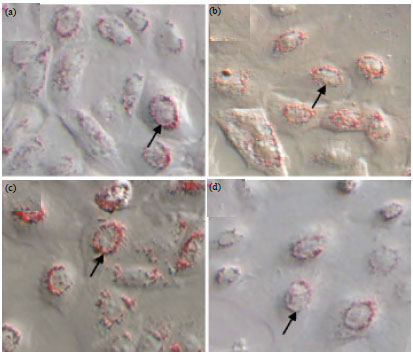 | |
| Fig. 5(a-d): | Oil red O staining of Huanghuai white goat mammary gland epithelial cells (200x magnification), (a) Passage 2, (b) Passage 4, (c) Passage 9 and (d) Passage 16 |
Oil red O staining: Oil red O staining revealed diffuse distribution of considerable numbers of bead-like fat droplets of varying sizes throughout the cytoplasm. Lipid droplets connected into a whole and flower rings formed around the lipid droplets (Fig. 5a). The proportion of mammary epithelial cells, which synthesize the lipid droplets, gradually decreased with increasing passage number. The most significant decrease was observed after passage 16 (Fig. 5b-d).
DISCUSSION
To date, many reports have described the isolation of mammary epithelial cells from mammary gland tissues obtained during the lactation period. For example, Zheng et al. (2005) established mammary epithelial cell lines from dairy goats obtained during the lactation period. At day 7, cell extravasation was observed with normal growth retained at passage 15. Similarly, Ouyang and Qian (1999) observed extravasation of individual cells from explants at day 5, while at day 6, most mammary epithelial cells appeared around explants. Wang et al. (2010) established goat mammary epithelial cell lines from mammary gland tissues obtained at day 40 of the lactation period and extravasation of numerous mammary epithelial cells was observed at day 10. In this study, goat mammary gland tissues were obtained at day 60 of the gestation period. During the early phase of explant culture, the methods of a serum-assisted adherence and the dish was inverted for cultivation were adopted. When the adherence was firm after 12 h, tissue explants were cultured with medium in an upright dish. Cellular extravasation was observed at day 4, with the formations of a large halo of mammary epithelial cells around most explants at day 5. Cells reached 90% confluence at day 11.
It is worth noting that, in previous reports, there were no consistent situations when isolation and culture were performed whether to use extracellular matrixes such as rat tail collagen, etc., or not. For instance, Zheng et al. (2005) reported outgrowth of mammary epithelial cells from primary explants cultured in suited medium on collagen-coated plates. These cells grew well in the absence of collagen in subcultures. In contrast, Ouyang and Qian (1999) reported an absolute requirement for collagen for the growth of GMECs. Duo successfully established dairy cow mammary epithelial cells in the absence of collagen and found that the effect of collagen on the transcription of the β-casein gene in these cells was not remarkable. Ahn et al. (1995) used plastic stents without an extracellular matrix to establishing cow mammary epithelial cells lines that were shown to secrete αs1-, β-, κ-casein in vitro. Wicha et al. (1979) postulated that collagen promoted the growth and attachment of mammary epithelial cells onto the culture flask. Terramani et al. (2000) hypothesized that serum-containing-medium could provide sufficient adhesion factors, such as fibrin and vitronectin, without the necessity for an extracellular matrix. Rat tail collagen is mainly composed of type IV collagen and certain growth factors. The method for isolation of the rat tail collagen is challenging and its role the generation of mammary epithelial cell lines is not clear. In this study, rat tail collagen was not included in the method used for the isolation and culture of GMECs. Explants were cultured in the absence of medium for 10 h to imitate the function of collagen in primary culture in promoting the attachment of epithelial cells during the early phase. It was anticipated that this approach would promote the attachment of explants and facilitate epithelial cell extravasation. The results of this study confirmed that the use of mammary gland tissue obtained during the gestation period as a primary source for the establishment of mammary epithelial cell lines, does not require the use of an extracellular matrix.
Tong et al. (2012) established Guanzhong dairy goat mammary epithelial cell lines using a cultivation system consisting of DMEM/F-12 basic medium supplemented with 10% serum, ITS, EGF and IGF-1 biotic factors. Cao et al. (2008) established Saanen dairy goat mammary epithelial cell lines using a similar cultivation system consisting of DMEM/F-12 basic medium supplemented with 10% serum, estradiol, hydrocortisone, EGF, IGF-1 and ITS. Peng et al. (2005) found that ITS was required for primary culture and subculture of cow mammary epithelial cells while EGF was not required but promoted significant proliferation of these cells. However, Colomb et al. (1991) demonstrated that estradiol and EGF were necessary for normal in vitro culture of human breast cells. The high cost of materials such as hormones and growth factors renders long-term in vitro culture of these cells extremely expensive. The cultivation system used in this study consisted of DMEM/F-12 basic medium supplemented with 10% serum, ITS and EGF biotic factors to improve the culture, thus reducing the cost of in vitro cell culture. It can be speculated that ITS and EGF promote cell proliferation while inhibiting the main function of differentiation in breast cells cultured in vitro.
The purification of GMECs was based on the differential sensitivity of mammary epithelial cells and fibroblasts to trypsin during the first four to five passages. In the course of subculturing, the proliferation rate of mammary epithelial cells at low density was lower, with a larger cell size and clustering than that observed at higher density cultures, in which cells were a consistent shape and high vigour. It can be speculated that these differences relate to stronger interactions among cells at a high density and the depression of autonomous "dedifferentiation". Furthermore, cell death in continuous subculture was lower and viability was higher, than that observed in recovery cultures after cryopreservation.
In this study, continuous subculture was performed at a passage ratio of 1:2. Results showed that prior to passage 10, cells required subculturing every other day, between passages 10 and 15, at 1.5 day intervals and every 2 to 3 days between passages 15 and 25. Cells observed up to passage 30 exhibited normal cell morphology and good viability although the cell volume was slightly enlarged. In subcultured cells (passages 2, 4, 9 and 16), the synthesis of milk fat and the expression of β-casein were detected, indicating lactation function in the established mammary epithelial cells.
This study successfully demonstrated a method for the establishment of mammary epithelial cell lines that retain lactation function derived from goat mammary gland tissue during the gestation period, using a cultivation system without extracellular matrix. Furthermore, compared to similar studies on tissues obtained during the lactation period, this method demonstrated earlier extravasation of mammary epithelial cells and more rapid proliferation.
CONCLUSION
The present study demonstrated a successful method for the establishment of mammary epithelial cells from goat mammary gland tissue of gestation stage, simplified the separation procedures of primary cell lines, reduced the cost of culture and formed the basis of further research into mammary gland development at the cellular levels and the mechanism underlying the regulation of lactation.
ACKNOWLEDGMENT
This study is supported by grants from by National High-Tech RandD Program (2011AA100307-04) and National Transgenic Breeding Program (No.2011ZX08008-005; No. 2009ZX08008-007B).
REFERENCES
- Ahn, J.Y., N. Aoki, T. Adachi, Y. Mizuno, R. Nakamura and T. Matsuda, 1995. Isolation and culture of bovine mammary epithelial cells and establishment of gene transfection conditions in the cell. Biosci. Biotechnol. Biochem., 59: 59-64.
PubMedDirect Link - Colomb, E., P. Berthon, C. Dussert, F. Calvo and P.M. Martin, 1991. Estradiol and EGF requirements for cell-cycle progression of normal human mammary epithelial cells in culture. Int. J. Cancer, 49: 932-937.
CrossRef - Danielson, K.G., C.J. Oborn, E.M. Durban, J.S. Butel and D. Medina, 1984. Epithelial mouse mammary cell line exhibiting normal morphogenesis in vivo and functional differentiation in vitro. Proc. Natl. Acad. Sci. USA., 81: 3756-3760.
Direct Link - Duo, S.G., Y.J. Wu, F.H. Luo and S. Bou, 2006. Isolation, culture and biological characteristics of bovine mammary epithelial cells. Zool. Res., 27: 299-305.
Direct Link - He, Y.L., Y.H. Wu, X.N. He, F.J. Liu, X.Y. He and Y. Zhang, 2009. An immortalized goat mammary epithelial cell line induced with human telomerase reverse transcriptase (hTERT) gene transfer. Theriogenology, 71: 1417-1424.
CrossRef - Ilan, N., I. Barash, E. Gootwine and M. Shani, 1998. Establishment and initial characterization of the ovine mammary epithelial cell line NISH. In vitro Cell. Dev. Biol. Anim., 34: 326-332.
CrossRef - Macias, H. and L. Hinck, 2012. Mammary gland development. Wiley Interdiscip Rev. Dev. Biol., 4: 533-557.
PubMedDirect Link - Peng, X.R., Y.M. Zheng and Y. Zhang, 2005. Study on the appearance of vitro-cultured bovine mammary epithelial cells. J. Northwest Sci-Tech Univ. Agric. For., 33: 13-17.
Direct Link - Samiec, M. and M. Skrzyszowska, 2011. The possibilities of practical application of transgenic mammalian species generated by somatic cell cloning in pharmacology, veterinary medicine and xenotransplantology. Pol. J. Vet. Sci., 14: 329-340.
CrossRef - Tai, D.P., L. Kun, Terigele, D.X. Hou, Y. Li and Y. Li, 2011. The biological characteristic analysis on three different chicken embryo tissue cells cultured in vitro. Chin. J. Cell Biol., 33: 988-993.
Direct Link - Terramani, T.T., D. Eton, P.A. Bui, Y. Wang, F.A. Weaver and H. Yu, 2000. Human macrovascular endothelial cells: Optimization of culture conditions. In vitro Cell. Dev. Biol.-Anim., 36: 125-132.
Direct Link - Tong, H.L, Q.Z. Li, X.J. Gao and D.Y. Yin, 2012. Establishment and characterization of a lactating dairy goat mammary gland epithelial cell line. In vitro Cell. Dev. Biol. Anim., 48: 149-155.
CrossRef - Warri, A.M., N.M. Saarinen and S.I. Makela, 2007. Can modulation of mammary gland development by dietary factors support breast cancer prevention? Horm. Res., 68: 248-260.
PubMedDirect Link - Wicha, M.S., L.A. Liotta, S. Garbisa and W.R. Kidwell, 1979. Basement membrane collagen requirements for attachment and growth of mammary epithelium. Exp. Cell Res., 124: 181-190.
CrossRef - Yi, N. and N. Li, 2010. Transient expression of chicken antimicrobial peptides by mouse mammary carcinoma cells C127. Protein Pept. Lett., 17: 1517-1523.
PubMed - Zheng, Y.M., Z.X. An, X.R. Peng, Y.Q. Shi and Y. Zhang, 2005. Establishment of goat mammary epithelial cell line and expression of EGFP gene. J. Agric. Biotechnol., 13: 322-325.
Direct Link - Zhou, Y., L. Ren, J. Zhu, S. Yan and H. Wang et al., 2011. Construction of a recombinant human FGF1 expression vector for mammary gland-specific expression in human breast cancer cells. Mol. Cell. Biochem., 354: 39-46.
CrossRef - Hu, H., J. Wang, D. Bu, H. Wei, L. Zhou, F. Li and J.J. Loor, 2009. In vitro culture and characterization of a mammary epithelial cell line from Chinese Holstein dairy cow. PLoS One, Vol. 4.
CrossRef