
Gyo-Moon Chu
Swine Science and Technology Center, Gyeongnam National University of Science and Technology, Jinju, Korea
Young-Min Song
Department of Animal Resource Technology, Gyeongnam National University of Science and Technology, Jinju, Korea
Asian Journal of Animal and Veterinary Advances
Year: 2013 | Volume: 8 | Issue: 5 | Page No.: 691-702







ABSTRACT
As part of living organisms, animals require complex environmental conditions and they have evolved mechanisms to cope with stress caused by changing environmental conditions. Environmental stress includes but not limited to being separated from dams, adapting to novel surroundings, struggling for rank and housing conditions. These factors interacting with genetic background can reduce the growth performance, decrease immune response and elevate cortisol concentration of an animal leading to compromised productivity in mass. This study was carried out so as to investigate the effects of different seasons on the growth performance, blood characteristics and immune responses in fattening pigs. Crossbred barrows of approximately 85 kg body weight were divided into 4 groups for each season (6 pigsx3 pensx4 seasons) and the diet was given for 4 weeks. The blood samples were collected from 4 pigs in each pen 2 h after the last feeding. the temperature inside was 23.8, 35.7, 22.1 and 11.8°C in spring, summer, autumn and winter, respectively when blood samples were collected. Summer significantly lowered (p<0.05) Average Daily Gain (ADG) and feed efficiency as well as concentrations of platelets, Immunoglobulin M (IgM), Immunoglobulin G (IgG) and interferon-gamma (IFN-γ) compared with other seasons. Summer also significantly increased (p<0.05) concentrations of leukocytes, Lactate Dehydrogenase (LDH) and cortisol compared with other seasons. Therefore, these results indicated that the different seasons enhanced growth performance, blood characteristics and immune responses and summer was more stressful than spring, autumn and winter in fattening pigs.
PDF Abstract XML References Citation
Received: March 21, 2013;
Accepted: June 03, 2013;
Published: August 05, 2013
How to cite this article
Gyo-Moon Chu and Young-Min Song, 2013. Growth Performance, Blood Characteristics and Immune Responses of Fattening Pigs in Different Seasons. Asian Journal of Animal and Veterinary Advances, 8: 691-702.
DOI: 10.3923/ajava.2013.691.702
URL: https://scialert.net/abstract/?doi=ajava.2013.691.702
DOI: 10.3923/ajava.2013.691.702
URL: https://scialert.net/abstract/?doi=ajava.2013.691.702
INTRODUCTION
Growth function have been used extensively to represent changes in Body Weight (BW) with age, so that intake level and diet composition can be matched to possible growth and the genetic potential of animal can be evaluated (Lopez et al., 2000). Growth is influenced by genotype, environment and feed factors in pigs (Bastianelli and Sauvant, 1997). Environmental stress includes being separated from dams, adapting to novel surroundings and struggling for rank with each other (Oh et al., 2010) and it is caused by housing conditions and can reduce growth performance (Hyun et al., 2005; Wolter et al., 2000), decrease immune response (McGlone and Curtis, 1985) and elevate cortisol concentration (Arnone and Dantzer, 1980). Therefore, environmental stress can eventually compromise swine productivity.
Animal’s environment is complex and good environmental conditions are needed for life to continue. All living organisms have evolved mechanisms that enable them to cope with changing environmental conditions and stress responses (Salak-Johnson and McGlone, 2007). Generally, growing-finishing pigs were reported to optimally perform at temperatures between 18 and 21°C (Huynh et al., 2005). The pigs show increased respiration rate and rectal temperature and decreased feed intake at higher ambient temperature (Quiniou et al., 2001). Moreover, a cycling hot environment of 27-32°C for 28 days has been reported to induce elevated rectal temperature and several immunological effects in sows due to stress (Becker and Misfeldt, 1994). Therefore, heat stress may affect the immune system of pigs.
The response of the immune system is one of the mechanisms that organisms have developed to defend themselves against environmental challenges. It was reported that stressed animals showed a suppressed immune system that lead to enhanced susceptibility to diseases (Salak-Johnson and McGlone, 2007). Cytokines in animals are known to modify their growth and regulate the function of immune system. It is readily apparent that the regulatory jurisdiction of cytokines as interleukin-1 (IL-1), interleukin-6 (IL-6) and tumor necrosis factor-α (TNF-α) extends well beyond the immune system. Many types of cells possess receptors of several pro-inflammatory cytokines that directly regulate metabolism of nutrients and growth in animals (Spurlock, 1997). Glucocorticoids, prostaglandins and catecholamines are modulators of the immune system that are raised by cytokines and they affect cell metabolism and growth. In addition, cytokines were also found to affect numerous physiological events that have been identified and put into a hypothesis which states that pro-inflammatory cytokines coordinate a homeorhetic response by which nutrients are driven away from tissue growth during periods of suppressed immune system (Spurlock, 1997).
To our knowledge, no reports have shown a comparison between the growth performances, blood characteristics and immune system responses due to the different seasons in fattening pigs. Therefore, the present experiment was conducted to study the growth performance, blood characteristics and immune responses, such as antibodies and cytokines under the different season conditions in fattening pig.
MATERIALS AND METHODS
Animals and diet: The BW of fattening pigs used in this study was approximately 85 kg and the age was approximately 150 days at the initiation of the study. Seventy two Durocx(Yorkshire x Landrace) crossed barrows were assigned into the 4 different seasons of spring, summer, autumn and winter (6 pigsx3 pensx4 seasons) which were balanced for initial BW. The pigs were conventionally housed individually in pens of 5.0x10.0 m and had free access to water and diet. They had free access to water and diet for 4 weeks. Taking care of these animals was according to the Guide for the Care and Use of Laboratory Animals (Animal Care Committee for Gyeongnam National University of Science and Technology).
The basal diet formulation is shown in Table 1. The basal diet mainly contained approximately 51.80% corn, 14.59% wheat and 11.12% soybean meal, respectively. This was consisted of 14.00% crude protein, 3.13 Mcal kg-1 metabolism energy, 0.80% lysine, 0.70% calcium and 0.40% total phosphorus. The pigs fed basal diet and were fed approximately from 85-110 kg of average BW for 4 weeks. The pigs also were fed the same diet in spring, summer, autumn and winter.
Temperatures and sampling: Experiment period, the lowest, highest and average temperature inside and outside is shown in Table 2.
| Table 1: | Composition of experimental diet (as-fed basis) |
 | |
| 1Supplied per kg of diet: 8000 IU vitamin A, 4000 IU vitamin D, 1500 IU vitamin E, 150 mg vitamin K, 20 mg vitamin B12, 20 mg niacin, 10 mg thiamin, 20 mg pantothenic acid, 4.0 mg riboflavin, 0.6 mg folic acid, 0.03 mg Biotin, 60 mg Zn, 60 mg Fe, 25 mg Mn, 15 mg Cu, 0.25 mg Se and 0.20 mg I | |
| Table 2: | Temperature inside and outside of animal house in different seasons |
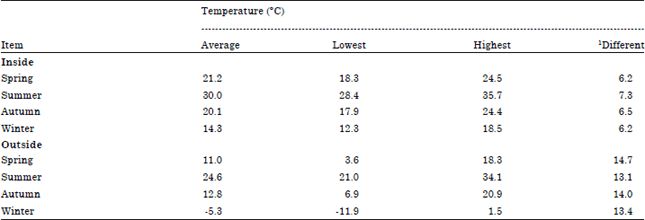 | |
| The inside humidity was 55-63% for experiment periods. 1Values showns the differences of highest and lowest temperature | |
Average temperature inside was 21.2, 30.0, 20.1 and 14.3°C in spring, summer, autumn and winter, respectively and Average humidity was approximately 55-63% in all seasons. The average temperature outside was 11.0, 24.6, 11.0 and -5.3°C in spring, summer, autumn and winter, respectively.
The BW was measured at the initiation and end of this experiment. Consumption amounts of diet were recorded every day at feeding trials. The feed efficiency was calculated as the Average Daily Gain (ADG) and feed intake. At the end of the experiment (one day before shipping), 2 h after the end of the last feed, blood samples were collected from jugular vein of 4 pigs in each pen (4 pigsx3 pensx4 seasons). The blood samples were collected at 15:00 o’clock in all seasons and the temperature inside was 23.8, 35.7, 22.1 and 11.8°C in spring, summer, autumn and winter, respectively.
Determination of blood corpuscle and chemical composition: The blood corpuscles as leukocytes, erythrocytes, hemoglobin, hematocrit, Mean Corpuscular Volume (MCV), Mean Corpuscular Hemoglobin (MCH), Mean Corpuscular Hemoglobin Concentration (MCHC) and platelets were determined using an automatic hematological analyzer (VET abc, Montpellier, France) within 2 h of blood sampling. Plasma was obtained by centrifugation at 2,500 g for 30 min at 4°C and stored at -20°C for determination of chemical composition. The chemical composition in plasma as total protein, albumin, Lactate Dehydronase (LDH), total cholesterol, triglyceride, Blood Urea Nitrogen (BUN), High Density Lipoprotein (HDL)-cholesterol and Low Density Lipoprotein (LDL)-cholesterol was determined using Blood Analyzer (Express Plus, Bayer, MA, USA). The cortisol concentration was determined using an Immulite (Diagnostic product Co., USA) method for chemiluminescent immuno-assay.
Determination of antibodies: Total porcine immunoglobulin was determined in serum as previously described in a report by Mizumachi et al. (2009). The blood was collected in a heparinized vacutainer tube (Becton Dickinson, San Jose, CA, USA) then centrifuged at 10,000 g for 2 min after clotting and stored individually at -80°C. Total porcine antibodies as immunoglobulin M (IgM), immunoglobulin G (IgG) and immunoglobulin A (IgA) were measured by a sandwich Enzyme-linked Immunosorbent Assay (ELISA). For the detection of IgM, IgG and IgA, microtiter plate wells (Maxisorp; Nunc, Roskilde, Denmark) were coated overnight at 4°C with goat anti-porcine IgM, IgG or IgA (Bethyl Laboratories, Montgomery, TX, USA). The coated wells were washed with Phosphate Buffer Saline (PBS) containing 0.05% Tween 20 (PBST) and blocked with 1% Bovine Serum Albumin (BSA) in PBS for 30 min. After washing with PBST, the approximately diluted samples were added and incubated for 2 h at Room Temperature (RT). Then the wells were treated with horseradish peroxidase-conjugated goat anti-porcine IgM, IgG or IgA (Bethyl Laboratories, USA) for 1 h at RT. The wells were washed and 3,3’,5,5’-Tetramethylbenzidine (TMB) solution (KPL, Gaithersburg, MD, USA) was added to each well as a substrate. After 30 min of incubation at RT, the reaction was stopped by the addition of 1 M dihydrogen phosphate (H2PO3). Absorbance was measured at 450 nm using a microplate reader (Original multi scan, Thermo, USA).
Determination of cytokine production: Determination of cytokine production in whole blood was described by Edfors-Lilja et al. (1998) and Mizumachi et al. (2009). Heparinized whole blood was diluted to 1:10 for the detection of IL-6, interleukin-8 (IL-8) and TNF-α and 1:50 for the detection of interferon-γ (IFN-γ) by the culture medium as a Roswell Park Memorial Institute-1640 (RPMI-1640) containing 50 μM 2-mercaptoethanol, 10 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES), 10 U mL-1 penicillin and 100 μg mL-1 streptomycin. The diluted samples were seeded into 24-well plates and cultured in a humid incubator. The incubator was maintained at 37°C and 5% CO2 gas. The culture supernatant was collected after 6 h for determination of IL-8 and TNF-α and was collected after 72 h for determination of IL-6 and IFN-γ. The cytokine levels were determined by commercial ELISA reagents (IL-6, IL-8 and TNF-α; R&D Systems, Minneapolis, MN, USA and IFN-γ; PharMingen, San Diego, CA, USA) according to the manufacturer’s procedures.
Statistical analyses: The data were analyzed in ANOVA using the General Linear Model (GLM) procedure of SAS (1999) and significant differences among the means were determined using the Duncan’s Multiple Range Test method at p<0.05 (Duncan, 1955).
RESULTS
The inside humidity was approximately 55-63% in all seasons and the animals had not been any diseases for experiment periods (data not shown).
The different seasons affected the growth performance and feed efficiency in fattening pigs (Table 3). The final BW and average daily gain were significantly lower (p<0.05) in summer than in spring, autumn and winter and it was not different between spring, autumn and winter. The finished body weight in summer was 100.67 kg while that of spring, autumn and winter were 107.83, 107.17 and 107.83 kg, respectively. The average daily gain was 0.815, 0.530, 0.810 and 0.850 kg day-1 for spring, summer, autumn and winter, correspondingly. Also, the average daily feed intake and feed efficiency were significantly lower (p<0.05) in summer than in spring, autumn and winter and it was also not different between spring, autumn and winter. The average daily intake was 2.010, 1.832, 2.057 and 2.357 kg for spring, summer, autumn and winter, sequentially. The feed efficiency was 0.406, 0.289, 0.394 and 0.361 for spring, summer, autumn and winter in order. There were significant differences (p<0.05) in average daily gain and feed efficiency between summer and other seasons.
The results of concentration of blood corpuscle are shown in Table 4. Although the concentrations of erythrocytes, hemoglobin, hematocrit, MCV, MCH and MCHC were not affected (p>0.05) by season, it did negatively affect the concentrations of leukocytes and platelets.
| Table 3: | Growth performance and feed efficiency of fattening pigs in different seasons |
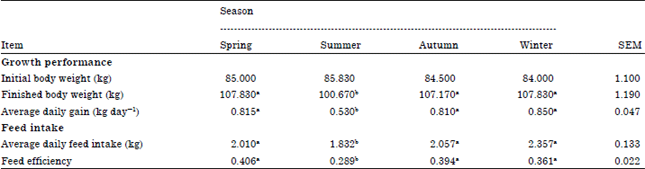 | |
| a-bValues in the same row with different superscripts differ at p<0.05 | |
| Table 4: | Blood corpuscles concentration of fattening pigs in different seasons |
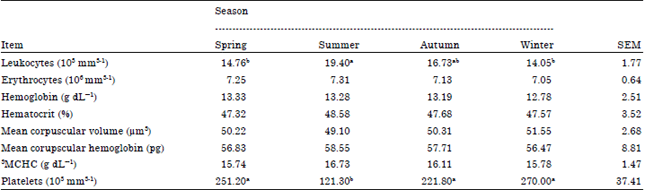 | |
| 3Mean corupscular hemoglobin concentration. a-bValues in the same row with different superscripts differ at p<0.05 | |
The leukocytes concentration was significantly lower (p<0.05) in spring (14.76x103 mm3-1) and winter (14.05x103 mm3-1) compared with summer (19.40x103 mm3-1) while the platelets concentration in summer (121.3x103 mm3-1) was significantly decreased (p<0.05) compared with other seasons and it was not different between spring (251.2x103 mm3-1), autumn (221.8x103 mm3-1) and winter (270.0x103 mm3-1).
The results of different seasons on chemical composition in plasma are shown in Table 5. The different seasons did not affect the concentration of total protein, albumin, total cholesterol, triglyceride, HDL-cholesterol and LDL-cholesterol while it significantly affected (p<0.05) the concentration of LDH, BUN and cortisol in the plasma. The LDH concentration was significantly higher (p<0.05) in summer (937.70 U L-1) than in other seasons and it did not differ between spring (799.2 U L-1), autumn (782.1 U L-1) and winter (798.7 U L-1). The BUN concentration was significantly higher (p<0.05) in summer (16.33 mg dL-1) than in winter (10.10 mg dL-1). The cortisol concentration in summer (5.67 μg dL-1) significantly increased (p<0.05) compared with other seasons and it did not differ between spring (2.83 μg dL-1), autumn (2.33 μg dL-1) and winter (2.57 μg dL-1). There were significant differences (p<0.05) in the concentrations of LDH and cortisol in the plasma between summer and other seasons.
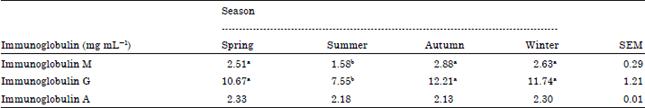
The effects of different seasons on antibodies as total serum IgM, IgG and IgA concentrations are shown in Table 6. Although the different seasons did not affect the concentration of total serum IgA, it affected the concentrations of total serum IgM and IgG. The concentrations of total serum IgM and IgG in summer (1.58 and 7.55 mg mL-1, respectively) were significantly decreased (p<0.05) compared with other seasons and it did not differ between spring (2.51 and 10.67 mg mL-1), autumn (2.88 and 12.21 mg mL-1) and winter (2.63 and 11.74 mg mL-1, respectively).
| Table 5: | Plasma chemical composition of fattening pigs in different seasons |
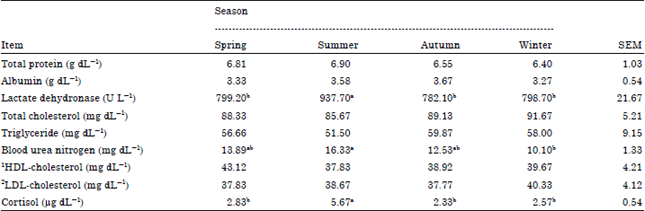 | |
| 1High density lipoprotein cholesterol. 2Low density lipoprotein cholesterol. a-bValues in the same row with different superscripts differ at p<0.05 | |
| Table 6: | Antibodies as IgM, IgG and IgA concentration of fattening pigs in different seasons |
 | |
| 1Not detected. a-bValues in the same row with different superscripts differ at p<0.05 | |
| Table 7: | Cytokine production of fattening pigs in different seasons |
 | |
| 1Not detected. a-bValues in the same row with different superscripts differ at p<0.05 | |
There were significant differences (p<0.05) in the concentration of total serum IgM and IgG between summer and other seasons.
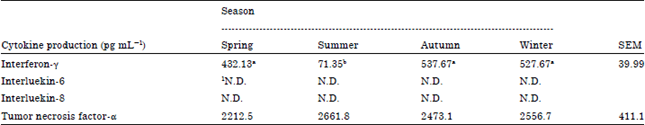
The effects of different seasons on cytokine production as concentrations of IFN-γ, IL-6, IL-8 and TNF-α in blood cells are shown in Table 7. The IFN-γ concentration of blood cells in summer (71.35 pg mL-1) was significantly decreased (p<0.05) compared with other seasons and it did not differ between spring (432.13 pg mL-1), autumn (537.67 pg mL-1) and winter (527.67 pg mL-1). The concentrations of IL-6 and IL-8 were not detectable in all seasons. The TNF-α concentration of blood cells was not significantly different (p>0.05) between spring, summer, autumn and winter. There was significant difference (p<0.05) in IFN-γ concentration of blood cells between summer and other seasons.
DISCUSSION
Season was considered to be an important risk factor for growth and temperature also was considered to be the most important stressor in multi-factorial variable (Lewis and Berry, 2006). In this study, average daily gain, daily feed intake and feed efficiency in summer decreased compared with other seasons. Kim et al. (2000) reported that the ADG and feed efficiency were decreased in summer than in winter in fattening pigs. Therefore, the results of growth performance and feed efficiency in this experiment were suggested by Kim et al. (2000).
Many studies reported that decreased BW, decreased antibody synthesis, weaned cellular immunity and lowered pathogenic resistance may be observed when animals are exposed to environmental stress (Kelly, 1980; McGlone, 1990; Murtaugh et al., 1996; McCracken et al., 1999). Moreover, many factors may influence the immune response of an animal due to stress. Stress may suppress, enhance or have no effect on the immune status of an animal. Many conflicting factors such as stressor, age, genetic and social status may affect animals. Stress may shift the balance from a helper T-cell 1 (Th1)-like to a helper T-cell 2 (Th2)-like response. If the immune system is in a predominantly Th2 state, the animal have enhanced protection against bacteria to which it has been previously exposed. If the immune system is skewed toward Th2, then viral and early pathogen immunity would be suppressed and the animals would be more likely to have an allergic or autoimmune disease. Therefore, a better understanding of the complexity of these relationships in farm animals will increase the likelihood that animal health and well-being will be improved (Salak-Johnson and McGlone, 2007).
Appropriate development of humoral and cellular functions of immune system is essential for optimum growth and performance of the piglets (Sinkora et al., 2002). Humoral immunity is the produced antibodies by B cells that cause the destruction of extracellular microorganisms and prevent the spread of intra-cellular infections. Cellular immunity involves in the activity of T-lymphocytes derived from the thymus gland. The T-lymphocytes directly destroy virally infected cells and can control the activity of other cells involved in the immune response. Expansion and activation of T cells may indicate the induction of a cellular immune response which promotes secretion of multiple pro-inflammatory cytokines such as IL-1, IL-6 and TNF-α. Also pro-inflammatory cytokine can disturb the immune system and decreases feed efficiency and growth rate in pigs (Colditz, 2002; Pie et al., 2004; Sinkora et al., 2002). Klasing and Johnstone (1991) reported that nutrient cause a somatic growth to support the immune system. In other words, pro-inflammatory cytokines cause a shift in nutrient partitioning away from skeletal muscle accretion and towards the metabolic responses necessary to support the immune system. Inflammatory cytokines stimulate the secretion of stress hormone, such as cortisol which decreases the secretion of growth hormone (Fan et al., 1994).
Summer was found to increase the concentration of leukocytes and decrease platelets in this experiment. Leukocytes are cells of the immune system found in the blood and lymphatic system and involved in defending the body against both infectious diseases and foreign materials (Maton et al., 2008). Leukocytes number is 10,000 to 15,000 per μL in normal pigs and increases commonly two-folds when the pigs have an active infection or stress (McGlone and Pond, 2002). Therefore, this result indicated that summer might have more stress than spring, autumn and winter in fattening pigs. Platelet is 2-3 μm in diameter and its lifespan is normally 5-9 days. Platelets are fragments of the cells in bone marrow, called megakaryocytes. Stimulated by the hormone thrombopoietin, platelets break off the megakaryocytes and enter the blood stream. While a disease or a genetic disorder can cause a lower number of platelets, a higher than normal count of platelets is known as thrombocytosis which can cause serious health risks (Campbell et al., 2008). The count of platelets in this study indicated that there might be less chance of disease in spring, autumn and winter than summer in fattening pigs.
Summer increased LDH and cortisol concentrations of plasma chemical composition compared with spring, autumn and winter in this experiment. Also, BUN concentration in summer increased compared with winter. LDH plays an important role in cellular respiration through catalyzation by which pyruvate from glucose is converted into usable energy as lactate in cells. When tissues are damaged by injury or disease, they release more LDH into the bloodstream. Moreover, conditions that can cause increased LDH in the blood include liver disease, heart attack, anemia, muscle trauma, bone fractures, etc. (Joseph et al., 2002). The results of this study suggest that summer cause more damage or stress than spring, autumn and winter in fattening pigs.
The BUN is the amount of urea nitrogen as a waste product of protein metabolism in the blood. There are many factors beside renal disease that can cause BUN alterations, including protein breakdown, hydration status and liver failure. Diseased or damaged kidneys cause an elevated BUN because the kidneys are less able to clear urea from the bloodstream (Johnson et al., 1972). The results of this study showed that winter caused less diseased or damaged kidneys than summer in fattening pigs.
Cortisol is a steroid hormone or glucocorticoid produced by the adrenal gland and released in response to stress. Its primary functions are to increase blood sugar by gluconeogenisis, suppress the immune system, aid in fat, protein and carbohydrate metabolism and decrease the bone formation (Tao, 2009). Moreover, cortisol prevents proliferation of T-cells by rendering the IL-2 or IL-1 and T-cell growth factor. Cortisol also has a negative-feedback effect on IL-1 which is useful in combating some diseases as hypothalamus (Palacios and Sugawara, 1982). The results of this study showed that summer caused more stress than spring, autumn and winter in fattening pigs.
Summer decreased the concentrations of IgM and IgG in serum compared with spring, autumn and winter while it did not affect IgA concentration. IgM is the first antibody formed when white blood cells are initially exposed to an antigen and it is a pentomer molecule with five linked antibodies. When exposed to an antigen for a second time, the pig builds very high levels of antibodies, mostly in the class of IgG. White blood cells switch from synthesizing IgM to synthesizing IgG after continued antigen exposure. Serum IgG is the major constituent of blood immune globulin and this antibody plays a major role in defending against antigens. Concentration of serum IgG increases over time as the immune system matures, but the increased serum IgG is suppressed in heat-stressed pigs (Murphy et al., 2008). Morrow-Tesch et al. (1994) reported that serum IgG concentration was affected in pigs that were simultaneously exposed to heat stress and social stress. IgA referred to as secretory antibody and IgA found at higher concentrations in the fluids of the mucous membranes (respiratory, gastrointestinal, reproductive tracts and eyes). Finally, IgD and IgE function as allergic and anaphylactic responses (McGlone and Pond, 2002). Supplemental probiotics and fermented liquid feed increased antibodies in pigs (Corthesy et al., 2007; Mizumachi et al. (2009). The results of this study indicated that spring, autumn and winter increased the immune responses as IgM and IgG and may prevent diseases or stress compared with summer in fattening pigs.
Summer greatly decreased the IFN-γ concentration of cytokine while it did not affect TNF-α concentration compared with spring, autumn and winter. IL-6 and IL-8 were not detectable in all seasons. T and B cells play an important role in the immune system response, in which T-cells control most of the B-cells producing antibodies. IFN-γ secreted by Th1 cells and IL-4 and IL-6 secreted by Th2 cells Mizumachi et al. (2009). In this experiment, the temperature of different seasons affected IFN-γ secretion from blood cells while it did not affect IL-6 secretion. Therefore, the temperature is expected to affect more Th1 type immune responses. Antigen-presenting cells induce production of different T-cell cytokines while mucosal cells induce predominantly Th2 cytokines (Everson et al., 1996). The temperature of different seasons might stimulate Th2 response via mucosal sites and it has been reported that some types of B cells appear regulated in a T-cell independent manner (Fagarasan and Honjo, 2000). These antibodies are produced in the absence of any exogenous antigenic exposure without the help of T cells and play an important role in the first line of defense against bacterial and viral infections (Mizumachi et al., 2009). In this experiment, T-cells-independent antibodies might have been produced in spring, autumn and winter.
CONCLUSION
The temperature of different seasons affected the growth performance and immune responses as antibodies and cytokine production in fattening pigs. Summer decreased the growth performances and feed efficiency in this study. Moreover, summer increased the concentrations of leukocytes and cortisol concentration while decreased the concentrations of IgA, IgG and IFN-γ in fattening pigs. Therefore, these results indicated that the seasons of spring, autumn and winter may decrease stress. Hence, this study suggests that, pigs raised in autumn to winter or winter to spring, provide healthier pigs with low cost, because stress makes pigs vulnerable to diseases that may need treatment and increase cost. Further investigations are required to clarify the effects of temperature of different seasons on the mechanisms of blood immune responses in pigs.
ACKNOWLEDGMENT
This study was supported by Priority Research Centers Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (2012-0006683). We would like also to thank Adoni Agricultural Union Corporation, Geochang, Korea for their kind help with the feeding experiment.
REFERENCES
- Arnone, M. and R. Dantzer, 1980. Does frustration induce aggression in pigs? Applied Anim. Ethol., 6: 351-362.
CrossRef - Bastianelli, D. and D. Sauvant, 1997. Modelling the mechanisms of pig growth. Livestock Prod. Sci., 51: 97-107.
CrossRef - Becker, B.A. and M.L. Misfeldt, 1994. Effect of hot environment on cell surface differentiation antigens of porcine peripheral lymphocytes from sows and litters. J. Thermal Biol., 19: 163-169.
CrossRef - Colditz, I.G., 2002. Effects of the immune system on metabolism: Implications for production and disease resistance in livestock. Livest. Product. Sci., 75: 257-268.
CrossRefDirect Link - Corthesy, B., H.R. Gaskins and A. Mercenier, 2007. Cross-talk between probiotic bacteria and the host immune system. J. Nutr., 137: 781S-790S.
Direct Link - Edfors-Lilja, I., E. Wattrang, L. Marklund, M. Moller, L. Andersson-Eklund, L. Andersson and C. Fossum, 1998. Mapping quantitative trait loci for immune capacity in the pig. J. Immunol., 160: 829-835.
Direct Link - Everson, M.P., D.S. McDuffie, D.G. Lemak, W.J. Koopman, J.R. McGhee and K.W. Beagley, 1996. Dendritic cells from different tissues induce production of different T cell cytokine profiles. J. Leukocyte Biol., 59: 494-498.
Direct Link - Fagarasan, S. and T. Honjo, 2000. T-independent immune response: New aspects of B cell biology. Science, 290: 89-92.
CrossRef - Fan, J., P.E. Molina, M.C. Gelato and C.H. Lang, 1994. Differential tissue regulation of insulin-like growth factor-I content and binding proteins after endotoxin. Endocrinology, 134: 1685-1692.
PubMed - Huynh, T.T.T., A.J.A. Aarnink, M.W.A. Verstegen, W.J.J. Gerrits, M.J.W. Heetkamp, B. Kemp and T.T. Canh, 2005. Effects of increasing temperatures on physiological changes in pigs at different relative humidities. J. Anim. Sci., 83: 1385-1396.
Direct Link - Hyun, Y., M. Ellis, S.E. Curtis and R.W. Johnson, 2005. Environmental temperature, space allowance and regrouping: Additive effects of multiple concurrent stressors in growing pigs. J. Swine Health Prod., 13: 131-138.
Direct Link - Johnson, W.J., W.W. Hagge, R.D. Wagoner, R.P. Dinapoli and J.W. Rosevear, 1972. Effects of urea loading in patients with far-advanced renal failure. Mayo Clin. Proc., 47: 21-29.
PubMed - Joseph, J., P. Badrinath, G.S. Basran and S.A. Sahn, 2002. Is albumin gradient or fluid to serum albumin ratio better than the pleural fluid lactate dehydroginase in the diagnostic of separation of pleural effusion? BMC Pulmonary Med., Vol. 2.
CrossRef - Morrow-Tesch, J.L., J.J. McGlone and J.L. Salak-Johnson, 1994. Heat and social stress effects on pig immune measures. J. Anim. Sci., 72: 2599-2609.
Direct Link - Kelly, K., 1980. Stress and immune functions: A bibliographics review. Ann. Vet. Res., 11: 445-478.
Direct Link - Klasing, K.C. and B.J. Johnstone, 1991. Monokines in growth and development. Poult. Sci., 70: 1781-1789.
CrossRefDirect Link - Lewis, N.J. and R.J. Berry, 2006. Effects of season on the behaviour of early-weaned piglets during and immediately following transport. Applied Anim. Behav. Sci., 100: 182-192.
CrossRef - Lopez, S., J. France, W.J. Gerrits, M.S. Dhanoa, D.J. Humphries and J. Dijkstra, 2000. A generalized Michaelis-Menten equation for the analysis of growth. J. Anim. Sci., 78: 1816-1828.
PubMed - McCracken, B.A., M.E. Spurlock, M.A. Roos, F.A. Zuckermann and H.R. Gaskins, 1999. Weaning anorexia may contribute to local inflammation in the piglet small intestine. J. Nutr., 129: 613-619.
PubMed - McGlone, J.J. and S.E. Curtis, 1985. Behavior and performance of weanling pigs in pens equipped with hide areas. J. Anim. Sci., 60: 20-24.
PubMedDirect Link - McGlone, J.J., 1990. Potential for improving animal health by modulation of behavior and immune function. Adv. Vet. Sci. Comp. Med., 35: 307-315.
PubMedDirect Link - Murtaugh, M.P., M.J. Baarsch, Y. Zhou, R.W. Scamurra and G. Lin, 1996. Inflammatory cytokines in animal health and disease. Vet. Immunol. Immunopathol., 54: 45-55.
CrossRef - Oh, H.K., H.B. Choi, W.S. Ju, C.S. Chung and Y.Y. Kim, 2010. Effects of space allocation on growth performance and immune system in weaning pigs. Livest. Sci., 132: 113-118.
CrossRefDirect Link - Palacios, R. and I. Sugawara, 1982. Hydrocortisone abrogates proliferation of T cells in autologous mixed lymphocyte reaction by rendering the interleukin-2 producer T cells unresponsive to interleukin-1 and unable to synthesize the T-cell growth factor. Scand. J. Immunol., 15: 25-31.
CrossRef - Pie, S., J.P. Lalles, F. Blazy, J. Laffitte, B. Seve and I.P. Oswald, 2004. Weaning is associated with an up regulation of expression of inflammatory cytokines in the intestine of piglets. J. Nutr., 134: 641-647.
PubMed - Quiniou, N., I. Noblet, J. van Milgen and S. Dubois, 2001. Modelling heat production and energy balance in group-housed growing pigs exposed to low or high ambient temperatures. Br. J. Nutr., 85: 97-106.
CrossRefDirect Link - Salak-Johnson, J.L. and J.J. McGlone, 2007. Making sense of apparently conflicting data: Stress and immunity in swine and cattle. J. Anim. Sci., 85: E81-E88.
CrossRefPubMedDirect Link - Sinkora, J., Z. Rehakova, M. Sinkora, B. Cukrowska and H. Tlaskalova-Hogenova, 2002. Early development of immune system in pigs. Vet. Immunol. Immunopathol., 87: 301-306.
PubMed - Spurlock, M.E., 1997. Regulation of metabolism and growth during immune challenge: An overview of cytokine function. J. Anim. Sci., 75: 1773-1783.
Direct Link - Wolter, B.F., M. Ellis, S.E. Curtis, E.N. Parr and D.M. Webel, 2000. Group size and floor-space allowance can affect weanling-pig performance. J. Anim. Sci., 78: 2062-2067.
Direct Link