
M. Rozihan
Department of Aquaculture, Faculty of Agriculture, Universiti Putra Malaysia, 43400 UPM, Serdang, Selangor, Malaysia
E. Ismail
School of Bioscience and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600 UKM Bangi, Selangor D.E., Malaysia
B. Akbar John
Department of Biotechnology, Kulliyyah of Science, International Islamic University Malaysia, Jalan Sulthan Ahmed Shah, Bander Indera Mahkota, 25200, Kuantan, Pahang, Malaysia
K.C.A. Jalal
Department of Biotechnology, Kulliyyah of Science, International Islamic University Malaysia, Jalan Sulthan Ahmed Shah, Bander Indera Mahkota, 25200, Kuantan, Pahang, Malaysia
A. Mohd-Adnan
School of Bioscience and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600 UKM Bangi, Selangor D.E., Malaysia
Asian Journal of Animal and Veterinary Advances
Year: 2013 | Volume: 8 | Issue: 1 | Page No.: 63-72







ABSTRACT
The genetic structure of horseshoe crab (Tachypleus gigas) populations were examined using 18 microsatellite markers. Samples were collected from 2 stations of east coast of Peninsular Malaysia and one station from East Malaysia. All the observed allelic frequency showed no significant variation between sampling stations (p>0.05). Mean allelic richness (Ar) was greater in Sarawak population (4.83) followed by Pahang (4.24) and Terengganu (3.57) samples with the overall mean allelic richness of 4.21. Mean inbreeding coefficient (FIS) value was higher in Sarawak population (0.37) followed by Terengganu (0.31) and Pahang population (0.17). The genetic differentiation (FST value) and genetic distance between geographically closer populations was smaller compared to geographically isolated populations. Due to the dwindling population size of horseshoe crabs in Malaysian coast line, present data provides new information in assisting proper management and conservation of this living fossil.
PDF Abstract XML References Citation
Received: July 03, 2012;
Accepted: July 06, 2012;
Published: December 26, 2012
How to cite this article
M. Rozihan, E. Ismail, B. Akbar John, K.C.A. Jalal and A. Mohd-Adnan, 2013. Genetic Diversity of Horseshoe Crab (Tachypleus gigas) in Malaysia
Revealed using Microsatellite Markers. Asian Journal of Animal and Veterinary Advances, 8: 63-72.
DOI: 10.3923/ajava.2013.63.72
URL: https://scialert.net/abstract/?doi=ajava.2013.63.72
DOI: 10.3923/ajava.2013.63.72
URL: https://scialert.net/abstract/?doi=ajava.2013.63.72
INTRODUCTION
Horseshoe crabs are well known creatures remarkably retaining their genetic makeup virtually unchanged over millions of years (John et al., 2010). Numerous studies were attempted to explore their phylogeny and phylogeography of four extant species using mitochondrial DNA markers (Xia, 2000; Kamaruzzaman et al., 2011) while only few studies were done to investigate their population structure and genetic diversity using microsatellite markers (Li et al., 2009; Faurby et al., 2010). It was noted that all those studies concentrated mostly on specific species such as Tachypleus tridentatus and Limulus polyphemus while the population genetic studies on T. gigas is still limited especially in Malaysia (Rozihan and Ismail, 2011, 2012).
Tachypleus gigas population is established in both Peninsular Malaysia and east Malaysian coast. They migrate to the nesting grounds during full and new moon period to lay their eggs in loosely packed sediments in intertidal area (Akbar John et al., 2012). Field observation revealed the co-occurrence of T. gigas and C. rotundicauda in some nesting grounds along the east coast of Peninsular Malaysia. At present, the collection of T. gigas samples from the wild habitat and nesting grounds are becoming tedious because of dwindling population size due to habitat degradation besides their commercial fishery and biomedical importance (John et al., 2011). Hence, adequate management of this vulnerable species requires knowledge for their conservation, including information about their ecology and population structure. Here, this study presented the genetic differentiation within and among the T. gigas population noted in east coast of Peninsular Malaysia and East Malaysia using microsatellite molecular markers.
MATERIALS AND METHODS
Sample collection and microsatellite genotyping: A total of 31 horseshoe crab samples were collected from 3 states (Terengganu (N 05°41.000' E 102°42.594') = 11 samples; Pahang (N 03° 31.988' E 103°27.534') = 12 samples and Sarawak (N 01°38.944' E 110°28.508') = 8 samples) (Fig. 1). Samples were identified morphologically, sexed and weighed. A sterile scissor was used to excise 2x2 cm of book gill tissue and preserved in the 95% ethanol for further analysis and horseshoe crab samples were released back into the water.
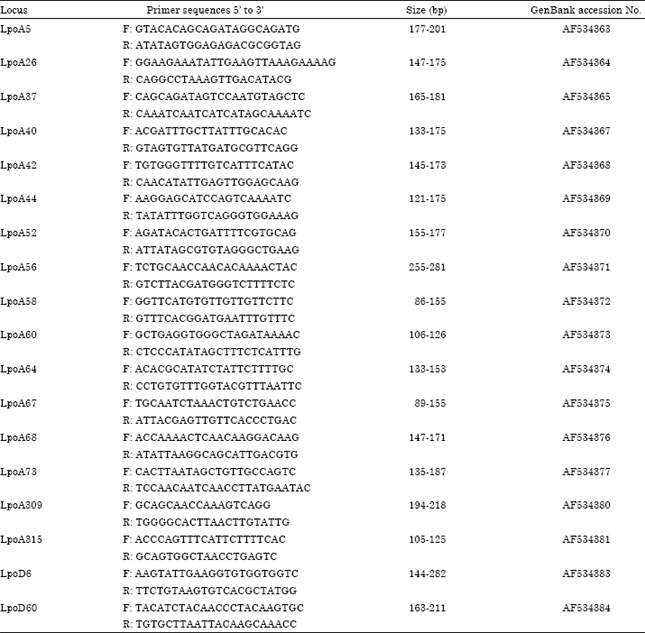
Total DNA was isolated from gill tissue using Promega® Wizard Genomic DNA Purification Kit. 18 microsatellite loci developed for American conspecific species (Limulus polyphemus) by King and Eackles (2004) were used in the amplification of scorable bands (Table 1). The amplified products were gel eluded using 5% Polyacrylamide gel. Alleles were sized using BioCapt software using 50 bp allelic ladder for consistent scrolling of alleles.
Data analysis: Van Oosterhout et al. (2004) method were adopted to check presence of null alleles and its dominance. GENEPOP 3.3v software was used to check the allelic frequencies and independence of genotypes among loci (Raymond and Rousset, 1995). FSTAT 2.93v was used to calculate genetic diversity parameters such as observed (Ho), expected heterozygosities (He) (under Hardy-Weinberg equilibrium) and Ar (allelic diversity) (Goudet, 1995). Pairwise FST values among sites and their significant variation among the sites (Analysis of Molecular Variance, AMOVA) were calculated using ARLEQUIN 3.1v (Schneider et al., 2000).
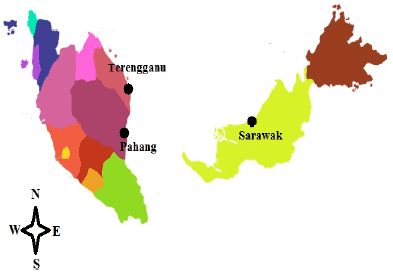 | |
| Fig. 1: | Location of the sampling are along the East Malaysian coast |
| Table 1: | Primer sequences of 18 polymorphic microsatellite loci that were used to characterize the three T. gigas populations in Malaysia |
 | |
RESULTS AND DISCUSSION
Allele frequency: A total of 118 alleles were detected at 18 microsatellite locus noted in horseshoe crab populations. Mean number of allele per locus was 3.7±0.4 in Terengganu, 5±0.9 in Pahang and 5.4±0.8 in Sarawak samples. It is well documented that high mutation rate and presence of more number of allelic variations in microsatellite sequence in a population could be useful in predicting population structure (Goldstein et al., 1999). The existence of flanking region in all the microsatellite loci was apparent in this study which eventually reflected polymorphisms in all the locus (Table 2). Similar situation was observed in American conspecific (King and Eackles, 2004).
Mean allele frequency of 18 microsatellite loci in horseshoe crab population showed no significant variation between sampling stations (p>0.05) which clearly revealed the existence of more than one population of horseshoe crab in east coast of Malaysia.
| Table 2: | No. of allele and its frequency observed at 18 microsatellite loci from three populations of T. gigas |
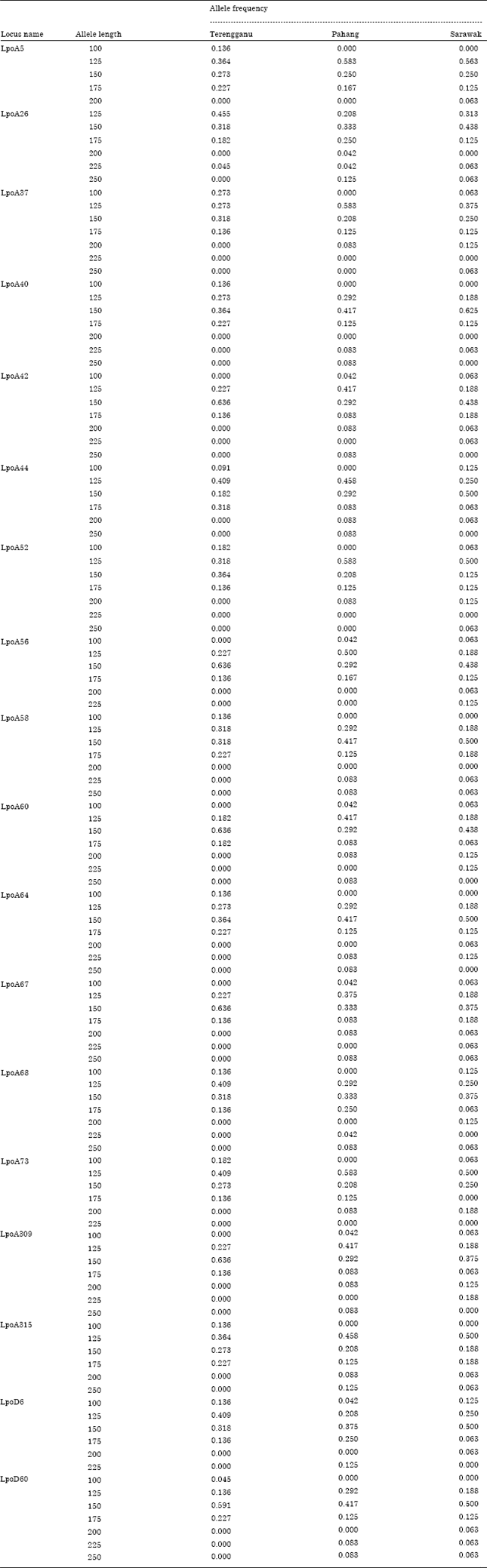 | |
In general, horseshoe crabs have limited migratory capacity and they migrate to shallow area for reproduction and the juveniles tend to stay at the natal beach for feeding (Sekiguchi, 1988; Chen et al., 2004). This might probably be the reason why they displayed genetic subdivision between populations and isolation by distance. Mean allelic richness (Ar) was greater in Sarawak population (4.83) followed by Pahang samples (4.24) and Terengganu crabs (3.57) with the overall mean allelic richness of 4.21 in east coast Malaysian samples. Mean number of alleles identified at a population is considered to be a good indicator of genetic variability (Leberg, 2002). Compared to the previous report on L. polyphemus microsatellite analysis by Orti et al. (1997), present study revealed less allelic variation in T. gigas population in East coast of Malaysia.
The observed heterozygosity (Ho) ranged from 0.25-0.75 in Sarawak, 0.33-0.75 in Pahang and 0.27-0.64 in Terengganu populations. Expected heterozygosity (He) was ranging from 0.62-0.86 in Sarawak population, 0.6-0.81 in Pahang samples and 0.56-0.76 in Terengganu crabs. Mean homo and heterozygosity values in Sarawak population were 0.48 and 0.72 which is slightly greater than Terengganu population (Ho = 0.47; He = 0.69) and lesser than Pahang samples (Ho = 0.59; He = 0.72). The fairly low heterozygosity value in Pahang population might be attributed to less number of allelic variations and high level of inbreeding (Aliabadi et al., 2008).
| Table 3: | Genetic variability at 18 microsatellite loci from three populations of T. gigas |
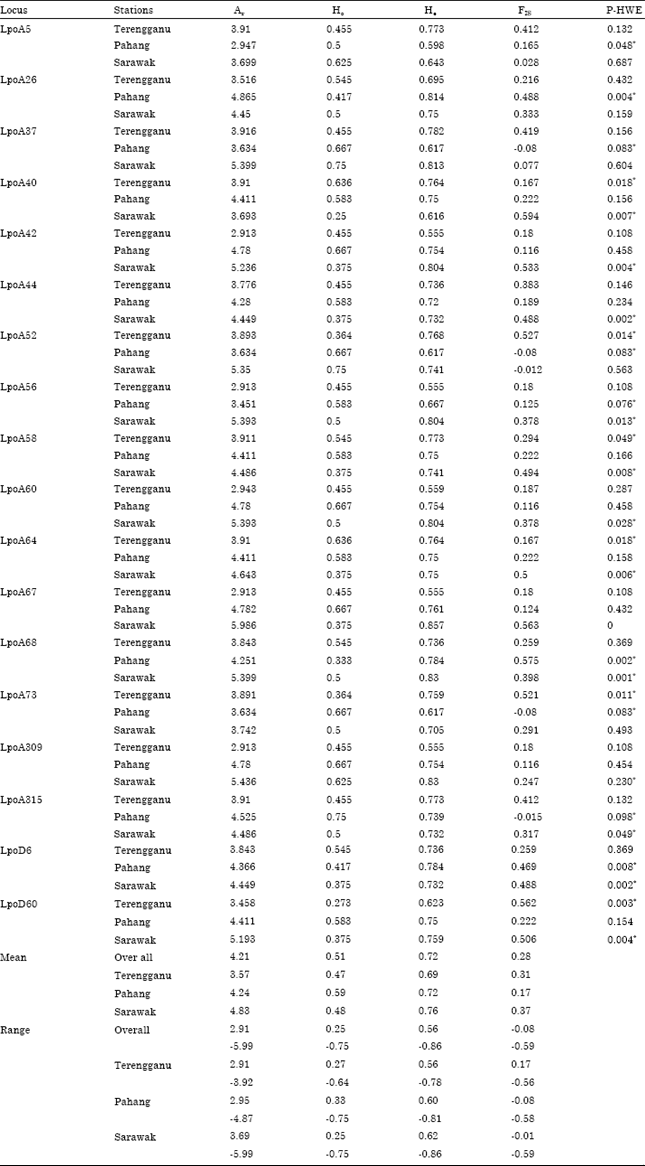 | |
| Ar: Allele richness, Ho: Observed heterozygosity, He: Expected heterozygosity, FIS: Inbreeding coefficient, HWE: Hardy-Weinberg equilibrium (*indicates statistical significance at 95% confidence interval) | |
Deviation from hardy-Weinberg equilibrium at the locus level had shown significant departure in multiple loci (Table 3). Mean inbreeding coefficient (FIS) value was higher in Sarawak population (0.37) followed by Terengganu (0.31) and Pahang population (0.17).
Genetic variation among sampling stations: The genetic differentiation (FST value) between geographically closer populations was smaller compared to geographically isolated populations. Lower FST value was recorded between Pahang and Terengganu samples (0.1441) followed by Terengganu and Sarawak population (0.5496) and Pahang/Sarawak samples (0.8469). However, the values were not statistically significant (p>0.05). Pairwise genetic distance value was lowest between Pahang/Sarawak population (-0.0113) followed by Terengganu/Sarawak (0.0047) and Pahang/Terengganu (0.0326) (Table 4). This observation contradicts with previous study on T. gigas genetic variation by Rozihan and Ismail (2012). Probable reasons are it might be due to the differential genetic cues in mitochondrial and microsatellite markers besides the number of samples collected from each sites could not reveal distinct genetic distance between geographically separated populations.
Analysis of Molecular Variance (AMOVA) using 18 microsatellite loci showed greater percentage variance among individuals of the same population (28.57%) (p<0.01) compared to the individuals of different population (7.47%) (Table 5). AMOVA test revealed the significant difference in sampled population might be due to rapid mutational rate in microsatellite loci (Wirgin et al., 2002). Probably the species specific microsatellite markers will reveal more genetic information on the population pattern and inbreeding rate of T. gigas population in Malaysian coast line.
| Table 4: | Estimates of pairwise genetic distance (Nei, 1978; below diagonal) and genetic differentiation FST (Weir and Cockerham, 1984; upper diagonal) among three populations of T. gigas |
 | |
| ns: Not significant | |
| Table 5: | Hierarchical analysis of molecular variance (AMOVA) in T. gigas |
 | |
| *Significant at p<0.01 | |
In conclusion, the data presented in this study revealed restricted migratory pattern of horseshoe crab (T. gigas) along the east coast of Malaysia. However extensive study need to carried out with more sampling size to conserve their genetic population subdivisions in natural habitat.
ACKNOWLEDGMENT
Author wish to extend his sincere gratitude to ministry of Higher Education Malaysia who funded this project under Fundamental Research Grant Scheme (FRGS) (Project Reference Number: 02-10-07-307FR).
REFERENCES
- John, B.A., K.C.A. Jalal, K. Zaleha, P. Armstrong and B.Y. Kamaruzzaman, 2011. Effects of blood extraction on the mortality of Malaysian horseshoe crabs (Tachypleus gigas). Marine Freshwater Behaviour Physiol., 44: 321-327.
Direct Link - Akbar John, B., S. Sihin, B.Y. Kamaruzzaman, K.C.A. Jalal and K. Zaleha, 2012. Sediment profiling of the nesting grounds of horseshoe crabs at East Peninsular Malaysia. Int. J. Biol., 4: 159-165.
CrossRefDirect Link - Chen, C.P., H.Y. Yeh and P.F. Lin, 2004. Conservation of horseshoe crabs in Kinmen, Taiwan: strategies and practices. Biodiversity Conservation, 13: 1889-1904.
Direct Link - Goldstein, D.B., G.W. Roemer, D.A. Smith, D.E. Reich, A. Bergman and R.K. Wayne, 1999. The use of microsatellite variation to infer population structure and demographic history in a natural model system. Genetics, 151: 797-801.
PubMedDirect Link - Faurby, S., T.L. King, M. Obst, E.M. Hallerman, C. Pertoldi and P. Funch, 2010. Population dynamics of American horseshoe crabs-historic climatic events and recent anthropogenic pressures. Molecular Ecol., 19: 3088-3100.
Direct Link - Goudet, J., 1995. FSTAT (version 1.2): A computer program to calculate F-statistics. J. Heredity, 86: 485-486.
Direct Link - John, B.A., K.C.A. Jalal, Y.B. Kamaruzzaman and K. Zaleha, 2010. Mechanism in the clot formation of horseshoe crab blood during bacterial endotoxin invasion. J. Applied Sci., 10: 1930-1936.
CrossRefDirect Link - Kamaruzzaman, B.Y., B.A. John, K. Zaleha and K.C.A. Jalal, 2011. Molecular phylogeny of horseshoe crab. Asian J. Biotechnol., 3: 302-309.
CrossRefDirect Link - King, T.L. and M.S. Eackles, 2004. Microsatellite DNA markers for the study of horseshoe crab (Limulus polyphemus) population structure. Mol. Ecol. Notes,, 4: 394-396.
CrossRef - Leberg, P.L., 2002. Estimating allelic richness: Effects of sample size and bottlenecks. Molecular Ecol., 11: 2445-2449.
Direct Link - Li, Q.Z., Q. Li, J. Liu, L.H. Ni and L.F. Kong, 2009. Isolation and characterization of microsatellite loci in the horseshoe crab (Tachypleus tridentatus). Conservation Genetics, 10: 1879-1881.
Direct Link - Nei, M., 1978. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics, 89: 583-590.
PubMedDirect Link - Raymond, M. and F. Rousset, 1995. An exact test for population differentiation. Evolution, 49: 1280-1283.
Direct Link - Rozihan, M. and E. Ismail, 2011. Genetic structure and haplotype diversity of Tachypleus gigas population along the West Coast of peninsular Malaysia-inferred through mtDNA AT rich region sequence analysis. Biotechnology.
CrossRefDirect Link - Rozihan, M. and E. Ismail, 2012. Impact of malaysian continental drift on the genetic diversity of horseshoe crab inferred through mtDNA sequence analysis. Int. J. Biol., 4: 104-110.
Direct Link - Aliabadi, M.A.S., S.R. Gilkolaei, A. Savari, H. Zolgharnein and S.M.B. Nabavi, 2008. Microsatellite polymorphism in Iranian populations of cobia (Rachycentron canadum G.). Biotechnology, 7: 775-780.
CrossRefDirect Link - Van Oosterhout, C., W.F. Hutchinson, D.P.M. Wills and P. Shipley, 2004. Micro-Checker: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes, 4: 535-538.
CrossRefDirect Link - Weir, B.S. and C.C. Cockerham, 1984. Estimating F-statistics for the analysis of population structure. Evolution, 38: 1358-1370.
CrossRefDirect Link - Wirgin, I., J. Waldman, J. Stabile, B. Lubinski and T. King, 2002. Comparison of mitochondrial DNA control region sequence and microsatellite DNA analysis in estimating population structure and gene flow rates in Atlanic sturgeon (Acipenser oxyrinchus). J. Applied Ichthyol., 18: 313-319.
Direct Link - Xia, X., 2000. Phylogenetic relationship among horseshoe crab species: Effect of substitution models on phylogenetic analyses. Syst. Biol., 49: 87-100.
Direct Link - Orti, G., D.E. Pearse and J.C. Avise, 1997. Phylogenetic assessment of length variation at a microsatellite locus. Proc. Natl. Acad. Sci. USA., 94: 10745-10749.
Direct Link