
Zi-Jun Zhang
College of Animal Science and Technology, Anhui Agricultural University, Hefei 230036, Peoples Republic of China
Ying-Hui Ling
College of Animal Science and Technology, Anhui Agricultural University, Hefei 230036, Peoples Republic of China
Chun-Huan Ren
College of Animal Science and Technology, Anhui Agricultural University, Hefei 230036, Peoples Republic of China
Xiao Cheng
College of Animal Science and Technology, Anhui Agricultural University, Hefei 230036, Peoples Republic of China
Hong-Guo Cao
College of Animal Science and Technology, Anhui Agricultural University, Hefei 230036, Peoples Republic of China
Xiao-Fei Guo
College of Animal Science and Technology, Anhui Agricultural University, Hefei 230036, Peoples Republic of China
Ya-Feng Huang
College of Animal Science and Technology, Anhui Agricultural University, Hefei 230036, Peoples Republic of China
Xiao-Rong Zhang
College of Animal Science and Technology, Anhui Agricultural University, Hefei 230036, Peoples Republic of China
Asian Journal of Animal and Veterinary Advances
Year: 2013 | Volume: 8 | Issue: 4 | Page No.: 571-581







ABSTRACT
Myostatin (MSTN) was a major gene of skeletal muscle growth regulation and a member of the transforming growth factor-β (TGF-β) super family. It acts as a negative regulator of skeletal muscle growth. To knock out goat MSTN gene of nuclear donor cells correctly, a gene targeting vector should be constructed. The homologous arm of goat MSTN was amplified by PCR and restriction enzyme digested. Intermediate vector pMD18-SA and pMD18-LA were constructed by conventional molecular cloning method. The final target vector pLOXP-MSTN was constructed by step-by-step cloning. The replacement type knock-out vector was transfected into goat fetal fibroblast by electrotransfection. The homologous arm sequences of goat MSTN including the 4.6 kb homologous long arm (LA) and the 1.9 kb homologous Short Arm (SA) were successfully amplified using LA-PCR technology. The LA and SA were inserted into vector pLOXP, generating MSTN gene targeting vector pLOXP-MSTN which contained neo and tk positive-negative-selection markers. The pLOXP-MSTN was linearized and electroporated into the nuclear donor cells. After G418 and GANC selection, fifty-eight drug resistant cell clones were screened and from which totally 3 positive clones were confirmed by PCR and DNA sequencing. The constructed gene targeting vector could efficiently target of MSTN locus and the gene knockout cell clones will be used to produce MSTN knockout goat by means of somatic cell nuclear transplantation.
PDF Abstract XML References Citation
Received: January 15, 2013;
Accepted: March 06, 2013;
Published: June 27, 2013
How to cite this article
Zi-Jun Zhang, Ying-Hui Ling, Chun-Huan Ren, Xiao Cheng, Hong-Guo Cao, Xiao-Fei Guo, Ya-Feng Huang and Xiao-Rong Zhang, 2013. Construction of Myostatin Gene Knockout Vector in Goat and Transfection Of Nuclear Donor Cells. Asian Journal of Animal and Veterinary Advances, 8: 571-581.
DOI: 10.3923/ajava.2013.571.581
URL: https://scialert.net/abstract/?doi=ajava.2013.571.581
DOI: 10.3923/ajava.2013.571.581
URL: https://scialert.net/abstract/?doi=ajava.2013.571.581
INTRODUCTION
Myostatin (MSTN), a growth and differentiation factor-β (GDF-β), is a member of the transforming growth factor-β (TGF-β) superfamily and acts as a negative regulator of skeletal muscle growth (Lu et al., 2012; Liu and Tong, 2012; Stinckens et al., 2011). MSTN is related to the control of skeletal muscle mass and the function’s lack will result in abnormal hypertrophy of skeletal muscle (Zhang et al., 2012; Jin, 2011). A mutation in the conservative function area of MSTN gene is closely related to meat yield, whose homozygote and heterozygote beef cattle show the advantage of strong muscle, increased birth weight and fast growth (Casas et al., 1999). The myostatin-deficient mice have developed by gene targeting, whose skeletal muscle weight are twice more than normal mice (Hamrick et al., 2002), but the fat tissue is not increasing. It showed that deletion or mutation of MSTN gene would result in obviously increasing in skeletal muscle growth (Chen et al., 2012). Therefore, producing MSTN gene knockout goat will increase goat’s carcass muscling, increase the efficiency of feed utilization and decrease raising cost.
Gene targeting is also called site-specific homologous recombination” or “gene knocking out”. It’s a genetic manipulation technique based on gene homologous recombination and embryonic stem cell (Zhan et al., 2011). This technique make fixed-point modified to the cell genome by homologous recombine exogenous DNA with cell genome DNA homologous sequence (Yamanouchi et al., 2000), achieving exogenous gene’s fixed-point integration, making the expectations that human beings transform biological genetic information directly into reality. At present, rodent such as mice, their gene-targeting techniques have been mature comparatively. Many scholars studied mammalian gene functions by the gene fixed-point knockout mice (Lin et al., 2001; Liou et al., 2005; Chen et al., 2002). The pig, cattle, sheep and other large mammals’ gene targeting of embryonic stem cells have been unable to achieve because of lacking mature embryonic stem cell lines. In recent years, the development of somatic cell cloning technology has provided effective technical support for the production of gene knockout large animals. Gene knock out of somatic cells, with target cells act as nuclear donor on somatic cell cloning, can obtain gene knockout animals, the gene knockout pig, cattle, sheep produced by this technology have successively been born (Lai et al., 2002; Kuroiwa et al., 2004; Zhu et al., 2009). It has been found that the main factors of influence the efficiency of the homologous recombination are the homology and length of the homologous sequence.
Before the homologous sequence was obtained, the length and homology of the sequence should be ensured. Li et al. (2000) construct myostatin gene target vector in pigs by gap repair and obtain gene knockout cells by transfection of Yorkshire pig fetal fibroblast cells. In this study, goat myostatin gene homologous arm sequences were successfully amplified using LA-PCR technology. Than the myostatin gene was replaced by neomycin phosphotransferase gene after the homologous arms cloned into pLOXP vector, for inactivating their function and achieving the knock-out purpose. The replacement type knock-out vector, pLOXP-MSTN, was identified by the enzyme digestion and sequence analysis and then transfected into goat fetal fibroblast by electrotransfection. That laid to the foundation for obtaining the myostatin gene deletion goat somatic cell line.
MATERIALS AND METHODS
Materials: Anhui white goat fetal fibroblast cells were kindly provided by Mr. Cao Hongguo in lab; Target vector pLOXP (Amp resistance) was kindly provided by Pan Dengke, who is a researcher in the institute of Animal Sciences, Chinese Academy of Agricultural Sciences; pMD18-T vector, Taq polymerase of highly-fidelity long fragment and DNA Marker, restriction enzymes (Xho I, SaL I, Not I and BamH I), T4 DNA ligase were bought from TaKaRa company in Dalian; Plasmid extraction kit was purchased from QIAGEN; Escherichia coli DH5a, cell genomic DNA extraction kits, gel purified recovery kits were purchased from TIANGEN Biotech (Beijing) CO.,LTD. Non-endotoxin plasmid DNA extraction kits were purchased from QIAGEN; Transfection reagent Basic Nucleofector® kit was purchased from LONZA company.
Primer design and synthesis: At present, goat MSTN gene sequence has not been reported. The goat MSTN gene primers used for amplification were designed according to sheep MSTN gene sequence (serial number: DQ530260.1). The primers used in this study were listed in Table 1.
| Table 1: | Primers used for goat MSTN gene amplification |
 | |
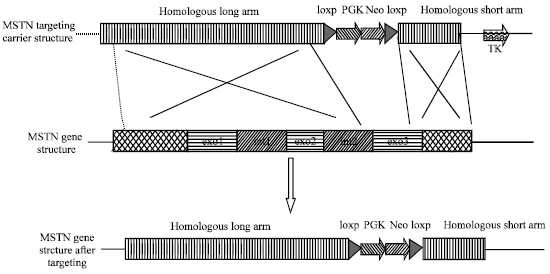 | |
| Fig. 1: | Sketch map of homologous recombination, Homologous long arm including the MSTN gene Exon 1 and 2, intron 1 and 2 and the gene sequences upstream 4605bp. Homologous short arm is MSTN gene exon 3 sequences downstream of 1856 bp |
The underlined sequences were 5’-end designed enzyme cutting sites. LAF and LAR were used for amplifying MSTN gene homologous long arms from goat fetal fibroblast genomic DNA. SAF and SAR were used for amplifying MSTN gene homologous short arms from goat fetal fibroblast genomic DNA. SAF1 and SAR1, NF and NR were used for identifying if the pLOXP-MSTN was integrated into goat fetal fibroblast cells to get resistant clone cells. SAF1 and SAR1 were used for amplifying homologous short arms from the pLOXP-MSTN integrated into goat fetal fibroblast cells. NF and NR were used for amplifying part of the neo gene sequence from the pLOXP-MSTN integrated into goat fetal fibroblast cells. Primers were synthesized by Shanghai Biotech CO., LTD.
PCR amplification of homologous arm: The genomic DNA from Anhui white goat fetal fibroblast cells was extracted according to the cell genomic DNA extraction kits. The quality of the genomic DNA was detected by agarose gel electrophoresis detection and ultraviolet spectrophotometer. Than the DNA was diluted into 50 ng μL-1 and saved at -20°C. A homologous short arm (SA, 1.9 kb), located in the exon 3 area of MSTN gene of goat, was obtained by PCR with primers SAF and SAR. A 4.6 kb homologous long arm (LA), covered the intron 1, exon 2 and intron 3 areas, was obtained by PCR with primers LAF and LAR (Fig. 1). PCR system contains 10xbuffer 2 μL, 2.5 mmol L-1 dNTP 0.8 μL, 0.4 μL of each primer, 5U μL-1 Taq enzyme 0.2 μL, ddH2O 15.2 μL, template 1 μL. For the long arm, the amplification conditions are the following parameters: an initial 5 min 94°C denaturing step, 30 cycles of the following temperature series -94°C for 1 min denaturing, 66°C for 6 min annealing, 72°C for 10 min extending, final 6 min extension at 72°C. For the short arm, the amplification conditions are the following parameters: an initial 5 min 94°C denaturing step, 30 cycles of the following temperature series-94°C for 55 sec denaturing, 55°C for 55s annealing, 72°C for 2 min extending, final 10 min extension at 72°C. PCR products were detected by electrophoresis and the purpose fragments were recovered by gel purified recovery kits.
Construction of intermediate vector pMD18-SA and pMD18-LA: SA and LA were cloned into pMD18-T vector (TaKaRa) respectively and transformed into Escherichia coli-DH5a (TIANGEN Biotech (Beijing) CO., LTD) competent cells by general molecule clone methods. The recombinant vector pMD18-T-SA, carried SA and pMD18-T-LA, carried LA, were obtained after the sequencing.
Step-by-step cloning of the final target vector pLOXP-MSTN: The final target vector was constructed by stepwise cloning method. Vector pLOXP (Amp resistance), provided by Pan Dengke, who is a researcher in the institute of Animal Sciences, Chinese Academy of Agricultural Sciences, included express components of positive selector gene neo, express components of negative selector gene HSV-tk and plasmid skeleton. The pLOXP and pMD18-T-SA were digested using Sal I and BamH I. Than the fragments after digestion of pLOXP and pMD18-T-SA which were 8.0kb and 1.9 kb respectively, were recovered by gel electrophoresis. Both fragments were connected using T4 DNA ligase and transferred to E.coli-DH5a. Restructuring carrier pLOXP-SA was obtained after identification. The pLOXP-SA and pMD18-T-LA were digested by Not I and Xho I. Corresponding fragments of pLOXP-SA and pMD18-T-LA which were 9.9 and 4.6 kb, respectively, were recovered by gel electrophoresis. Both fragments were connected using T4 DNA ligase and transferred to E.coli-DH5a. The MSTN gene targeting vector pLOXP-MSTN in Anhui white goat fetal fibroblast cells was obtained after screening recon, plasmid extraction and restriction enzyme identification.
The linearization of target vector: The plasmid pLOXP-MSTN was disposed using restriction enzyme digestion Not I at 37°C, 3-5 h. The plasmid was dissolved in TE after gel electrophoresis confirmation and alcohol precipitation. Linear plasmid DNA concentration was detected by UV spectrophotometer. The concentration was adjusted r 1.0 μg μL-1 for reserve.
The toxic sensitivity test of goat fetal fibroblast cells.
Goat fetal fibroblast cell lines constructed by the lab were thawed out to 24-hole boards according to the 2x104 of each hole and were divided into 8 groups of three repeated. Adding G418 whose different concentrations were 0, 100, 200, 400, 600, 800, 900, 1000 μg mL-1 in cells when 80~90% confluence was reached. Every 2 days changes one culture medium and replenish G418, record cell death conditions after culturing 2 weeks.
Cell electrotransfection, screening and appraisal: Cells were transfected with transfection instrument and transfection reagent Amaxa Nucleofector II (LONZA Company). Cells were inoculated to 100 mm culture dish before transfection. Cells were collected by digestion and counted accurately when 80~90% confluence was reached. Every 1x106 cells were resuspended by 100 μL transfection reagent, adding 8 μg linearizing pLOXP-MSTN vector to cell suspension, transferring to shock cup after adequate mixing, shocking it according to Nucleofector II program T016. Each shock cup’s cells were inoculated to single 60 mm culture dish after electric shock and keep on being cultured 48 h by DMEM which contains 20% FBS. Then cells were collected by digestion. Cells in every 60 mm culture dish were diluted and inoculated to six 96-hole boards, added culture solution that contained 400 μg mL-1 G418 after 24 h and transfection cells were screened for 5 to 10 days. Then using culture solution that contained 400 μg mL-1 G418 and 2 μM GANCD screen until cell confluence was reached, to get resistant cell clone which was inoculated to 24-hole boards and amplified, each hole’s cells were inoculated to 24-hole boards according to the proportion of 1 and 2 when cells fulled. Resistant cell clone DNA was extracted and further identified by PCR identification and gene sequence analysis.
RESULTS AND ANALYSIS
PCR amplification of goat MSTN gene LA and SA: Based on the published Myostatin gene sequence homology in sheep, two pairs of primers were designed and synthesized to amplify the 5’-end (4605 bp) 4.6 kb fragment (LA) and 3’-end (1866bp) 1.9 kb fragment (SA) of MSTN gene sequence. The PCR amplification results were showed in Fig. 2. The specificity of the bands was good and the size of fragment was consistent with expectation.
Identification of the transitional vector pMD18-T-SA and pMD18-T-LA: Plasmid pMD18-T-SA was digested by Sal I and BamH I to obtain 1.9kb SA fragment and 2.7 kb fragment of vector pMD-T (Fig. 3). Plasmid pMD18-T-LA was digested by Not I and XhoI to obtain 4.6 kb LA fragment and 2.7 kb fragment of vector pMD-T (Fig. 3).
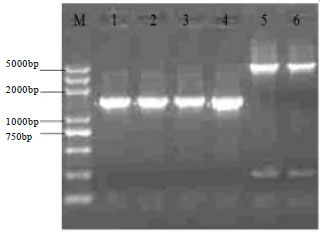 | |
| Fig. 2: | Electrophoresis of the two goat MNST fragments by PCR, 1, 2, 3, 4: 1.9 kb fragment, 5,6: 4.6 kb fragment; M: DNA Marker DL 5000 |
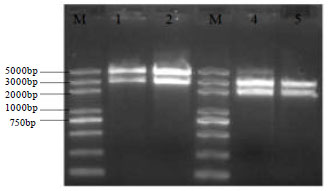 | |
| Fig. 3: | Restriction analysis of the pMD18-T-LA: pMD18-T-SA, 1,2: pMD18-T-LA digested by Notl+XhoI; 4,5: pMD18-T-SA digested by Sal 1+BamHI; M :DNA Marker DL5000 |
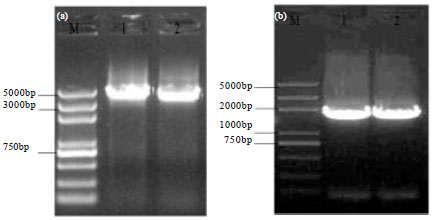 | |
| Fig. 4(a-b): | Identification of the pLOXP-MNST by PCR, 1.2: The PLOXP-MNST fragments by PCR; M: DNA Marker DL5000, a: PCR amplified by LAF+LAR Primers, b: PCR amplified by SAF+SAR Primers |
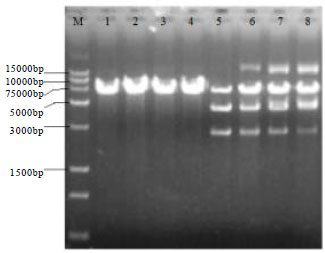 | |
| Fig. 5: | Identification of PLOXP-MNST by PCR and restriction enzyme digested, 1, 2, 3, 4: PLOXP-MNST; 5, 6, 7, 8: 5, 6, 7, 8/Not I+XhoI+Sal I+BamH I; M: DNA Marker DL15000 |
The restriction analysis of two transitional vectors was consistent with expected results. Sequencing indicated that insertion direction in exogenous DNA was correct and the inserted sequence was really MSTN gene fragment.
Evaluation of target vector PLOXP-MNST: PLOXP-MNST plasmid bacterial liquid was amplified by PCR using two pairs, LAF, LAR, SAF, SAR, respectively. The PCR products were detected by 1% agarose electrophoresis (Fig. 4a). The image of two amplified bands was clear and bright. The amplification fragment length was 4605 bp and 1866 bp, respectively. It’s consistent with expectation and both of the results were shown in Fig. 4.
Identification of PLOXP-MNST by PCR and restriction enzyme digested: PLOXP-MNST was digested by Sal I, BmH I, Not I, Xho I to obtain four bands of 12.3 kb, 1.4 kb, 9.4 kb and 4.3 bkb, respectively (Fig. 5). The identification of enzyme digestion showed that the construction of the PLOXP-MNST plasmid could be confirmed.
Confirmation of optimum concentration of G418: G418 (100~1000 μg mL-1) was confirmed toxic to goat fetal fibroblast cells using G418 treatment to normal cells (Table 2). The optimum filter concentration was confirmed as cells die both in 10 days and finally confirmed to be 400 μg mL-1.
Gene target efficiency: About 0.8x107 cells were transfected in a 60 mm culture dish and 10 culture dishes were used. In 11-14 days after transfection, cell colony appeared and 280 cell clones were got which were plated into the twenty four-well cluster dishes and screened. Then cell clones were plated into the six-well cluster dishes at a density of 90%. Finally one target cell line was obtained. After measurement, the absolute efficiency and relative efficiency of cell transfection were 1.25x10-8 and 3.6%, respectively.
The positive cells clone selection: Anhui white goat fetal fibroblast cells were electrotransfected with AmaxaNucleofector II program T016. After 10-15 days of pressure selection of the transfected cells in 96 orifice plate, the positive cells clone sites appeared as shown in Fig. 6. And 58 resistant cell lines were obtained.
Identification of positive cells by PCR: Drug-resistant cell clones of transfected linearizing plasmid PLOXP-MNST were amplified by PCR using primers SAF1 and SAR1 (Fig. 7a). A specific band, about 1.4 kb, was amplified from three in fifty-eight resistant clone cell lines; A target fragment of 409 bp was amplified according to the pair of NF and NR primers in neo gene region of empty targeting vector and is consistent with the size of the theory (Fig. 7b).
| Table 2: | Goat fetal fibroblast cells cultured in different concentration of G418 |
 | |
 | |
| Fig. 6: | Positive cells clone after selection x40 |
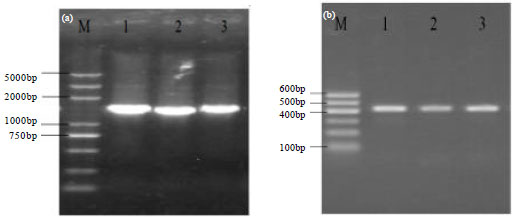 | |
| Fig. 7(a-b): | Identification of positive cells by PCR, 1, 2, 3: The positive cells fragments by PCR; M: DNA Marker, a: PCR amplified by SAF1+SAR1 Primers, b: PCR amplified by NF+NR Primers |
DISCUSSION
The construction of target vector was the key to the process of preparing gene knockout animals using somatic cells clone technology. In order to screen target cells more convenient, the structure of homologous arms fragment of carrier always be designed as one long and one short. However, the short arm couldn’t be shorter than 1 kb, or the efficiency of being targeted would reduce (Thomas et al., 1992). The length of the homologous arm had an effect on homologous recombination efficiency of transgenic structure in cells Li et al. (2000). Therefore, longer homologous arm sequences were amplificated to increase transfection effect (Thomas et al., 1992). Two pairs of primers were designed according to sequence of genetic base-pairs and obtained a 4.6 kb homologous long arm (LA) and a 1.9 kb homologous short arm (SA). In the two arms designed process, the restriction site of pLOXP directional connection and non-overlapping of two arms’ restriction site were taken account to design suitable amplified primer and make the mature peptide coding sequence swept away by middle knockout part, destroy MSTN gene activity of the original coding protein. The reaction condition of Long-PCR was so strict that Taq enzyme of highly-fidelity long fragment was needed. Moreover, when the longer fragment was amplified, the space resistance was bigger, amplification efficiency was lower and amplification products were connected harder. So the reaction must be connected at a lower temperature, increase mole ratio of inserted fragment relatively and extend connection time to increase connection efficiency (Dan et al., 2011). The enzyme sites were added into amplified products 5’-end of inserted fragment to reach directional connection destination when constructing vectors. After connection, the double digestion was used to connect with vector to ensure directionality.
In this study, the MNST gene-targeting vector pLOXP-MSTN constructed was 14.5 kb long. Among it, the homologous long arm (LA), 4.6 kb, covered the exon 1, exon 2 and intron 1, intron 2 areas of MSTN gene in goat. And the homologous short arm (SA), 1.9kb, was located in coding regions and 3’-untranslated regions of the exon 3 area. The partial exon 3 was replaced by Neo gene on carrier skeletons and it resulted that the gene function was deactivated, then to reach the goal of gene knockout Because the promoter upstream and polyA downstream of Neo gene were located in fit locus, respectively, it was convenient to reach the goal of gene knockout (Cox, 1989; Morris et al., 1991; Jayaram et al., 1988). It’s a replacement vector which directed at this area especially to design and knockout. pLOXPII-0VIS-MSTN was constructed as a deleted replacement vector which was aiming at MSTN exon 3 area in sheep (Dan et al., 2011). Site-directed mutagenesis of MSTN gene by gene targeting in qinchuan cattle was researched and one allele of myostatin was successfully mutagenized in bovine fetal fibroblasts (Liu et al., 2010).
At present, there were many kinds of nuclear donor cells in the research of mammalian somatic cell cloning. This study results were confirmed that the undifferentiated cell could support nuclear epigenetic reprogramming more effectively (Reik et al., 2001). The different effects on nuclear transplantation were researched according to the design when porcine adult and fetal fibroblasts, granular cells and oviductal epithelial cells were transfected individually and the effect of porcine fetal fibroblasts was the best (Hyun et al., 2003). Anhui white goat separated fetal fibroblast cells of 37 days old, had the advantages of easy separation, easy cultivation, more passage times, long time cultured in vitro and more steady nuclear type to satisfy the demand of long time cultured in vitro when screening, were selected in this study. Based on the theory of homologous recombination, replacement gene knockout vectors which had been constructed were transfected into primary cultured goat fetal fibroblasts by liposome-mediated method. Fixed-point knockouting goat MSTN gene, after screening for 5-10 days by G418 and positive cells were obtained after screening for 7-10 days by G418+GANC. Gene target cells obtained successfully by PCR identification will be used to produce MSTN gene knockout goat by means of somatic cell nuclear transplantation.
CONCLUSION
The MSTN gene was replaced by neomycin phosphotransferase gene after the homologous arms cloned into pLOXP vector, for inactivating their function and achieving the knock-out purpose. The replacement type knock-out vector, pLOXP-MSTN, was identified by the enzyme digestion and sequence analysis. And it was transfected into goat fetal fibroblast by electrotransfection. That laid to the foundation for obtaining the MSTN gene deletion goat somatic cell line.
ACKNOWLEDGMENTS
This study was supported by the Special Fund of China Agriculture Research System (11004986) and the fund of National Genetically Modified Organisms Breeding Major Projects of China, No. 2009 ZX08006-007B and Anhui Provincial Natural Science Foundation (1308085QC54).
REFERENCES
- Casas, E., J.W. Keele, S.C. Fahrenkrug, T.P. Smith, L.V. Cundiff and R.T. Stone, 1999. Quantitative analysis of birth, weaning and yearling weights and calving difficulty in Piedmontese crossbreds segregating an inactive myostatin allele. J. Anim. Sci., 77: 1686-1692.
PubMedDirect Link - Chen, H.Q., J. Qin, Y.J. Zhu, Z.T. Pan and Y.N. Xie et al., 2012. The polymorphisms of goat THRSP gene associated with ecological factors in Chinese indigenous goat breeds with different lipogenesis ability. Asian J. Aaim. Vet. Adv., 7: 802-811.
CrossRefDirect Link - Hyun, S., G. Lee, D. Kim, H. Kim and S. Lee et al., 2003. Production of nuclear transfer-derived piglets using porcine fetal fibroblasts transfected with the enhanced green fluorescent protein. Biol. Reprod., 69: 1060-1068.
CrossRefDirect Link - Chen, J.Y., B. Levy-Wilson, S. Goodart and A.D. Cooper, 2002. Mice expressing the human CYP7A1 gene in the mouse CYP7A1 knock-out background lack induction of CYP7A1 expression by cholesterol feeding and have increased hypercholesterolemia when fed a high fat diet. J. Biol. Chem., 277: 42588-42595.
CrossRef - Dan, S., J. Mei, D. Lixin, W. Caihong, Z. Li et al., 2011. Construction of sheep myostatin gene targeting vector. Biotechnol. Bull., 5: 93-100.
Direct Link - Kuroiwa, Y., P. Kasinathan, H. Matsushita, J. Sathiyaselan and E.J. Sullivan et al., 2004. Sequential targeting of the genes encoding immunoglobulin-mu and prion protein in cattle. Nat. Genet, 36: 775-780.
PubMedDirect Link - Lai, L., D. Kolber-Simonds, K.W. Park, H.T. Cheong and J.L. Greenstein et al., 2002. Production of alpha-13-gala-ctosyltransferase knockout pigs by nuclear transfer cloning. Science, 295: 1089-1092.
CrossRef - Jayaram, M., K.L. Crain, R.L. Parsons and R.M. Harshey, 1988. Holliday junctions in FLP recombination: Resolution by step-arrest mutants of FLP protein. Proc. Natl. Acad. Sci. USA., 85: 7902-7906.
Direct Link - Jin, X.Y., 2011. Cloning and bioformatics analysis on Myostain (MSTN) gene of goat. China Anim. Husbandry Vet. Med., 38: 111-114.
Direct Link - Lin, F., Y. Fukuoka, A. Spicer, R. Ohta and N. Okada et al., 2001. Tissue distribution of products of the mouse decay-accelerating factor (DAF) genes. Exploitation of a Daf1 knock-out mouse and site-specific monoclonal antibodies. Immunology, 104: 215-225.
PubMedDirect Link - Liou, Y.M., L.Y. Chang , J.M. Liaw and M.J. Jiang, 2005. Osteopontin gene expression in the aorta and the heartof propylthiouracil-induced hypothyroid mice. J. Biomedical Sci., 12: 869-880.
CrossRefDirect Link - Liu, L., X. Yu and J. Tong, 2012. Molecular characterization of myostatin (MSTN) gene and association analysis with growth traits in the bighead carp (Aristichthys nobilis). Mol. Biol. Rep., 39: 9211-9221.
CrossRefPubMedDirect Link - Liu, Y., S. Hua, J. Lan, Y. Song, Y. He, F. Quan and Y. Zhang, 2010. [Site-directed mutagenesis of MSTN gene by gene targeting in Qinchuan cattle]. Sheng Wu Gong Cheng Xue Bao, 26: 410-416.
PubMedDirect Link - Lu, J., D. Sun, L. Xu, G. Lu and F. Zhao et al., 2012. Selection of an effective small interference RNA to silence myostatin gene expression in sheep fibroblast cells. Biochem. Genet, 50: 838-847.
CrossRefDirect Link - Morris, A.C., T.L. Schaub and A.A. James, 1991. FLP-mediated recombination in the vector mosquite, Aedes aegypti. Nucleic Acids Res., 19: 5895-5900.
CrossRef - Reik, W., W. Dean and J. Walter, 2001. Epigenetic reprogramming in mammalian development. Science, 293: 1089-1093.
CrossRef - Stinckens, A., M. Georges and N. Buys, 2011. Mutations in the myostatin gene leading to hypermuscularity in mammals: Indications for a similar mechanism in fish? Anim. Genet., 42: 229-234.
CrossRef - Yamanouchi, K., C. Soeta, K. Naito and H. Tojo, 2000. Expression of myostatin gene in regenerating skeletal muscle of the rat and its localization. Biochem. Biophys. Res. Communi., 270: 510-516.
CrossRef - Zhang, C., Y. Liu, D. Xu, Q. Wen, X. Li, W. Zhang and L. Yang, 2012. Polymorphisms of Myostatin gene (MSTN) in four goat breeds and their effects on Boer goat growth performance. Mol. Biol. Rep., 39: 3081-3087.
CrossRefDirect Link - Zhu, C., B. Li, G. Yu, J. Chen and H. Yu et al., 2009. Production of Prnp-/- goats by gene targeting in adult fibroblasts. Transgenic Res., 18: 163-171.
CrossRef