
Sh. Shakibazadeh
Department of Aquaculture, Faculty of Agriculture, Universiti Putra Malaysia, 43400 Serdang, Selangor Darul Ehsan, Malaysia
A. Christianus
Department of Aquaculture, Faculty of Agriculture, Universiti Putra Malaysia, 43400 Serdang, Selangor Darul Ehsan, Malaysia
P. Hajeb
Centre of Excellence for Food Safety Research, Faculty of Food Science and Technology, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
Asian Journal of Animal and Veterinary Advances
Year: 2013 | Volume: 8 | Issue: 2 | Page No.: 275-283







ABSTRACT
Amoebocytes are the main carrier of innate immune system in horseshoe crabs. Cytoplasm of these cells are packed with granules which consist of all coagulation factors and anti-bacterial compounds. Light microscopy (using May-Grunwald and Giemsa staining technique) of 46 blood specimen for both sexes of Tachypleus gigas and observation under Transmission Electron Microscopy (TEM) were performed. Light microscopy revealed that these are ovoid cells containing blue nucleus surrounded by a cytoplasm packed with pink granules. Large and small axes measured 18.8±0.5 and 12.8±0.4 μm, respectively. Total blood cell count (33000 cell mm-3) does not show any significant difference between sexes. TEM of normal and degranulated blood cells revealed that T. gigas hemocytes consist of two type of granules (large and small). Blood cells have all the necessary organelles of normal cells. During the granule formation, fusion of different intermediate granules were observed. Degranulated hemocytes under light microscope and TEM showed cells containing large nucleus and cytoplasmic granules were replaced with a large cytoplasmic vacuole. In light and electron microscopy only one type of hemocyte was observed, the granular cells. These granular cells consist of large and small granules.
PDF Abstract XML References Citation
Received: July 02, 2012;
Accepted: July 05, 2012;
Published: February 11, 2013
How to cite this article
Sh. Shakibazadeh, A. Christianus and P. Hajeb, 2013. Blood Cell Histology of Horseshoe Crab, Tachypleus gigas. Asian Journal of Animal and Veterinary Advances, 8: 275-283.
DOI: 10.3923/ajava.2013.275.283
URL: https://scialert.net/abstract/?doi=ajava.2013.275.283
DOI: 10.3923/ajava.2013.275.283
URL: https://scialert.net/abstract/?doi=ajava.2013.275.283
INTRODUCTION
Invertebrates are thought to rely solely on their innate immune system (Kurtz, 2005). The circulating hemolymph in marine invertebrate contains biologically active substances such as lectins, anti-coagulant factors and anti-bacterial compounds. All of these factors contribute to the innate immune system in marine invertebrates against invading microorganisms which can reach to 106 bacteria mL-1 and 109 virus mL-1 in the sea water. The survival of marine invertebrates in this environment suggests a highly effective innate immune system (Tincu and Taylor, 2004).
In the horseshoe crab this innate immune system carried mainly in the hemolymph (Miyata et al., 1989) contained hemocytes or amoebocytes. Cytoplasm of these cells are packed with granules including all the factors required for blood coagulation which comprised of clottable protein and antibacterial compounds (Murer et al., 1975).
The blood of Limulus polyphemus contains only one type of cell, amoebocyte (Conrad et al., 2004). Under the transmission and scanning electron microscopy, the cytoplasm of the Limulus polyphemus amoebocytes are seen packed with only one type of granule (Murer et al., 1975). In Tachypleus tridentatus there are two types of hemocytes, non-granular and granular hemocytes. The latter contained numerous dense granules which are classed into two major types, large and small granules. The large granules contain three clotting factors and one anti-microbial compound and the small granules exclusively contained the other anti-microbial compounds (Toh et al., 1991). The intermediate granules were reported by Copeland and Levin (1985) in L. polyphemus, suggested that these are the intermediate stages for large granules. While Toh et al. (1991) described four types of Golgi vacuoles in Tachypleus tridentatus with various sizes and contents.
There are only four species of extant horseshoe crab. One; Limulus polyphemus in North and Central America (Botton and Rops, 1987), and the other three; Tachypleus tridentatus, Tachypleus gigas and Carcinoscorpius rotundicauda in South East Asia (Chiu and Morton, 1999).
In this study the blood cells of Tachypleus gigas, one of the species found in Peninsular Malaysia, were investigated through Light microscope and Transmission electron microscopy (TEM).
MATERIALS AND METHODS
In blood cell count and smear preparation, a total of 46 horseshoe crab specimens (22 females and 24 males) used were bought from fishermen in Pulau Lumut, Selangor, Malaysia during November 2004 to April 2005.
The blood was extracted by cardiac puncture. For blood extraction animals were fixed onto a rack and bent with dorsal surface towards front. The muscles above the heart were exposed and swabbed with 70% ethanol. Disposable pyrogen free 1 mL syringes and 25G needles were used to draw 0.2 mL hemolymph and then diluted to 1 mL with pre-chilled 10% formalin (in autoclaved seawater). The diluted hemolymph was stirred evenly by inversion and kept chilled for 1-2 h. The diluted hemolymph was used for blood cell count as well as smear preparation.
Blood cell count (repeated three times) of the diluted specimen was carried out using hemocytometer.
Blood smear was prepared by dropping the diluted blood onto 70% ethanol pre-cleaned slides and a clean slide used at 30-40° angle to prepare blood smear (Miy and Sparks, 1980). Each specimen has three replications. The slides were fixed for 1-2 min by absolute methanol, followed by May-Grunwald stain for 5-7 min, followed by Giemsa for 20-30 min.
Blood cell measurement was carried out using a total of seven specimens (each containing three replications). Both axes of the oval shaped granulocyte of 50 cells per slides were measured randomly. The measurements of red blood cells and photography were carried out using Image analyzer, PC_image (vga 24) software, Nikon microscope (Eclipse E 200) and a Nikon camera (Cool Pix 995).
TEM of fresh extracted blood was investigated (under extremely sterilized condition) for normal blood cell before and after degranulation. For normal blood cell before degranulation, a 0.5 mL of freshly extracted blood was immediately added into 10 mL volume of 3% glutaraldehyde in 0.1 M cacodylate buffer solution containing 0.1 M glucose for osmoregulation at 4°C. The cells were pre-fixed for 2 h then centrifuged at 500 rpm for 30 min, washed three times with 0.1 M cacodylate buffer (with 10 min interval). The cells then post-fixed in 1% osmium tetroxide (OsO4) in the same buffer solution for 3 h at 4°C. Same procedure carried out for clotted blood to view the cells after degranulation.
RESULTS
Under light microscope, blood cells of T. gigas were ovoid with light pink cytoplasm packed with red granules. A circular or ovoid blue nucleus was observed in the cytoplasm (Fig. 1). After the discharged of granules, the cytoplasm was empty. Some pseudopodia were observed to form and extended outward. A round and blue nucleus was observed (Fig. 2). Non-granular blood cells were extremely rare in T. gigas hemolymph. This cell consisted of a large purple nucleus which occupied most of the cytoplasm space. A thin crescent strip of light blue or purple cytoplasm was observed. There was no granule distinguished in the cytoplasm of this cell (Fig. 3).
The blood cell count ranged from 10,000-60,000 cell mm-3, with an average of 33,000 cell mm-3 (29,000 for females and 37,000 for males). Blood cell count of the sexes was compared using student t-test. There was no significant difference (p>0.05) between the blood cell counts of the sexes of T. gigas.
Blood cell measurements of T. gigas in terms of large and small axes were 18.8±0.5 and 12.8±0.4 μm, respectively.
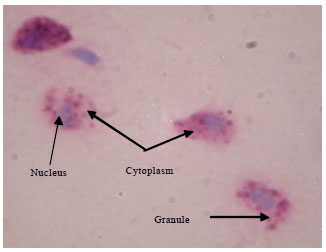 | |
| Fig. 1: | Light-microscopic image of normal granular blood cell of Tachypleus gigas magnified at 1000x |
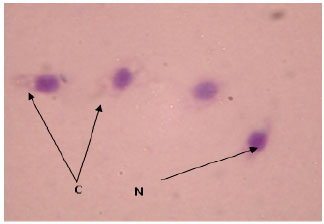 | |
| Fig. 2: | Tachypleus gigas granular blood cells after the discharge of granules, magnified at 1000x, N: Nucleus, C: Cytoplasm |
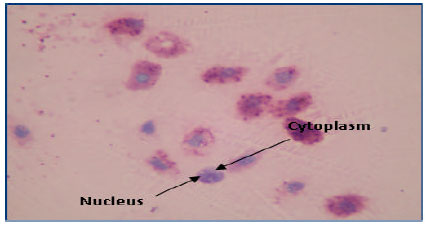 | |
| Fig. 3: | Nongranular blood cell of Tachypleus gigas magnified at 1000x |
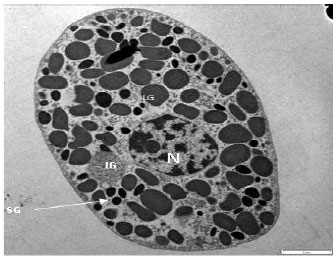 | |
| Fig. 4: | Normal granular blood cell of Tachypleus gigas, under TEM magnified at 1000x, N: Nucleus, IG: Intermediate granule, LG: Large granule, SG: Small granule |
TEM of normal granular cells of T. gigas revealed that the cytoplasm consisted of large (1 μm) and small (0.5 μm) granules. A large nucleus observed in the cytoplasm (Fig. 4). A well developed Golgi apparatus appeared close to the nucleus. The electron density of Golgi cisternae increased from the cis to the trans side with many small Golgi vesicles presented around the cisternae (Fig. 5b-e). Several spherical or oval shaped mitochondria were observed in the cytoplasm (Fig. 5f-h). Different intermediate granules as described by Toh et al. (1991) were tubular (T-granule), condense electron-dense (C-granule) and granules which contained patches of electron-dense material scattered in the lumen (P-granules). Moreover an intermediate granule with amorphous pattern of content was observed (Fig. 5a and b). Fusion of different intermediate granules such as, fusion of C and P-granule, C and T-granule (Fig. 5b, d and e) and fusion of two C-granules (Fig. 5d) were observed during the granules maturation.
Transmission Electron Microscopy of the granular blood cells after the discharged of granules is showed by the empty cytoplasm (no more granules remained in the cytoplasm).
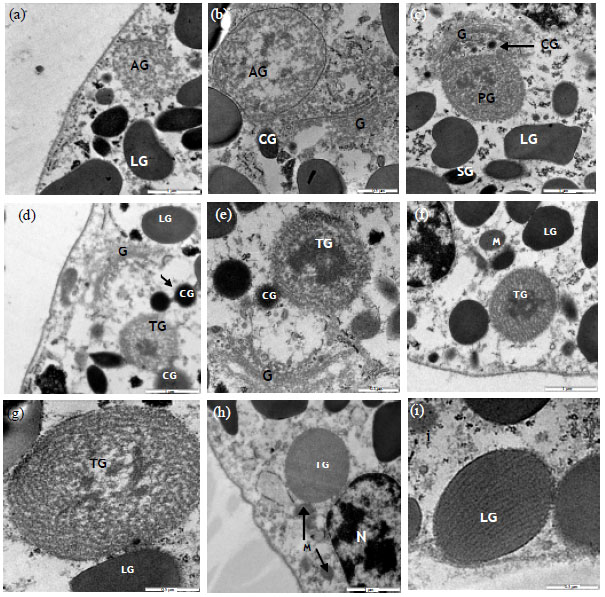 | |
| Fig. 5(a-i): | Transmission of electron microscopy of Tachypleus gigas of granular blood cell, (a) An amorphous intermediate granule (AG) and large granule (LG), 4000x, (b) An amorphous intermediate granule (AG) Containing few electron dense particles evaginated toward C-granules (CG) and Golgi apparatus (G), 6300x, (c) An Intermediate P-granule containing more regular pattern and dense particles (PG), Golgi apparatus (G), C-granule (CG), large (LG) And small granule (SG), 4000x, (d) Fusion of an intermediate T-granule containing an electron dense core (TG) with a C-granule (CG), Golgi apparatus (G), large granule (LG) and fusion of two C-granules (CG and arrow), 4000x, (e) Fusion of an intermediate T-granule containing an electron dense core (TG) in tubular pattern with a C-granule (CG) and Golgi apparatus (G), 4000x, (f) An intermediate T-granule containing an electron dense core (TG), mitochondria (M), Large granule (LG), 4000x, (g) An intermediate T-granule containing electron dense particle in tubular pattern (TG), Large granule (LG), 8000x, (h) An intermediate T-granule containing very regular and dense tubular pattern (TG), Mitochonderia (M) and nucleus (N), 4000x, (i) A large granule (LG) containing tubular pattern, 8000x |
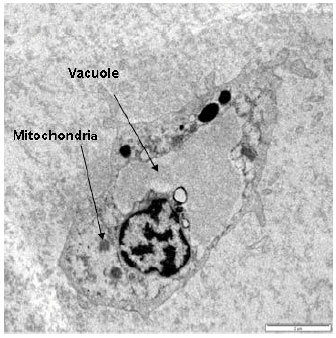 | |
| Fig. 6: | Transmission electron microscopy of Tachypleus gigas granular cell after the discharged of granules, magnified at 1300x |
Numerous vesicles was observed around a large and prominent nucleus. A large vacuole appeared in the cytoplasm of the granular cells after the discharged of their granules (Fig. 6).
DISCUSSION
Almost all the blood cells circulating in the hemolymph of the T. gigas are granulocytes. Non-granular cells. These cells are extremely rare (only one cell was observed during this study). Since, the respiratory pigment, hemocyanin is located extracellularly (Synder and Mangum, 1982) therefore, no mechanisms to transport the respiratory pigment like those of red blood cells in higher animals.
The average blood cells count of T. gigas was 33,000 cell mm-3 (29,000 for females and 37,000 for males). It was almost similar range with blood cell count of L. polyphemus (20,000-60,000 cell mm-3) as reported by Armstrong (1985). There was no significant difference between blood cell counts of sexes of the T. gigas using Student t-test.
The granulocytes were mostly oval or round, consisting of a large nucleus. The average granular cells size in terms of large and small axes were 18.8 μm±0.5 and 12.8 μm±0.4, respectively. The large axes measurements of T. gigas were within similar range (8-25 μm) to that reported by Sekiguchi (1988) and Conrad et al. (2004).
TEM observation revealed that the normal granular blood cells of horseshoe crabs, T. gigas, contained all the organelles of a well equipped cell which includes a nucleus, mitochondria, Golgi apparatus, endoplasmic reticulum, ribosome particles and microtubules. These observations were similar to those studies on L. polyphemus carried out by Dumont et al. (1966), Copeland and Levin (1985), Conrad et al. (2004) and on T. tridentatus by Jakobsen and Suhr-Jessen (1990) and Toh et al. (1991).
The granular cells were large and ovoid to fusiform, containing a large oval-shaped nucleus. The cytoplasm was packed with two types of granules, large and small. The average sizes for large and small granule were 1 and 0.5 μm in diameter, respectively. These measurements were similar to those reported for T. tridentatus, up to 1.5 μm and less than 0.6 μm for large and small granule, respectively (Toh et al., 1991) and L. polyphemus, 1-2 μm (Conrad et al., 2004). Presence of two types of granules in the granulocytes of T. tridentatus was reported by Toh et al. (1991) and Shigenaga et al. (1993) while Copeland and Levin (1985) reported that the small granules were not observed in all of the amoebocytes in L. polyphemus.
Normal granular blood cell observation in T. gigas showed a large well developed Golgi apparatus and its associated structure. A large recently made intermediate granule was usually observed closed to the Golgi complex. These intermediate granules mostly showed tubular (T-granule) and/or amorphous pattern in different densities which consist of electron dense core. The amorphous granules probably are early stage for P-granule, since, fusion with electron dense granule (C-granule) resulted in increased of electron density of the content and electron dense core inside the granule will appeared. Fusion of dense granules (C-granule) to the intermediate amorphous (P-granule) and tubular granules (T-granule) and even fusion of two electron dense granules (C-granule) were observed.
Copeland and Levin (1985) and Toh et al. (1991) suggested that the tubular granules are the intermediate form for large granules in L. polyphemus and T. tridentatus. Fusion of P and C-granule in T. tridentatus leads to the theory of small granule formation by Toh et al. (1991) however, this theory was not suggested in L. polyphemus.
Mitochondria appeared oval or spherical and possessed typical cristae and a relatively dense matrix which is similar to the observation by Dumont et al. (1966).
Rough endoplasmic reticulum appeared in different areas of the cytoplasm, mostly close to the nucleus and Golgi apparatus.
Although, studies on decapods crustacean and scorpion hemocytes in 1990 by Hose et al. (1990) and Ravindranath (1974), respectively, showed that the hemolymph consisted of two types of cells in crustacean and seven classes of cells in scorpion. TEM observation of normal blood cells of T. gigas did not show non-granular blood cells and it was rarely seen under light microscope.
TEM observation of clotted blood cells revealed that almost all two populations of granules were discharged. Cytoplasmic area was filled by different size vacuoles, some cytoplasm evaginations appeared, mitochondria and other organelles were seen in the medullary area of the cell, similar to those reported by Dumont et al. (1966).
Although, the granulocytes did not exhibit morphological differences (Sekiguchi, 1988), some usually discharge all their granules in respond to un-sterile condition while some remain intact. It is most likely that the activity of these granulocytes is due to their different physiological roles in the hemolymph.
CONCLUSION
Granular blood cell is the only cell found in the hemolymph of T. gigas. This cell consists of two types of granules, large and small. Amorphous intermediate granule seems to develop their structures during fusion with electron dense granules to tubular granule which is known as an intermediate granule for large granule. Small granule formation might be due to fusion of electron dense granules.
ACKNOWLEDGMENT
Authors thank the government of Malaysia for funding this project through the Intensified Research on Priority Areas (IRPA) grant No. 01-02-04-0864 EA001.
REFERENCES
- Botton, M.L. and J.W. Rops, 1987. Population of horseshoe crabs, Limulus polyphemus, on the Northwestern Atlantic continental shelf. Fish Bull., 85: 805-812.
Direct Link - Conrad, M., J. Denobile, I. Cheikhoutdinov, D. Escribano, K. Lee and W.D. Cohen, 2004. Cytoskeletal organization of Limulus amebocytes pre- and post-activation: Copmaretive aspects. Biol. Bull., 207: 56-66.
PubMedDirect Link - Copeland, D.E. and J. Levin, 1985. The fine structure of the amoebocyte in the blood of Limulus polyphemus, morphology of the normal cell. Biol. Bull., 169: 449-457.
Direct Link - Dumont, J.N., E. Anderson and G. Winner, 1966. Some cytologic characteristics of the hemocytes of Limulus during clotting. J. Morphol., 119: 181-207.
Direct Link - Hose, J.E., G.G. Martin and A.S. Gerald, 1990. A decapod hemocyte classification scheme integrating morphology, cytochemistry and function. Biol. Bull., 17: 33-45.
Direct Link - Kurtz, J., 2005. Specific memory within innate immune systems. Trends Immunol., 26: 186-192.
CrossRef - Miy, M.C. and A.K. Sparks, 1980. Hemocyte classification and differential counts in the Dungeness crab, Cancer magister. J. Invertebr. Pathol., 33: 134-143.
CrossRef - Miyata, T., F. Tokunaga, T. Yoneya, K. Yoshikawa and S. Iwanaga et al., 1989. Antimicrobial peptides, isolated from horseshoe crab hemocytes, tachyplesin II and polyphemusins I and II: Chemical structures and biological activity. J. Biochem., 106: 663-668.
Direct Link - Murer, E.H., J. Levin and R. Holm, 1975. Isolation and studies of the granules of the amebocytes of Limulus polyphemus, the horseshoe crab. J. Cell Physiol., 86: 533-542.
Direct Link - Ravindranath, M.H., 1974. The hemocyte of a scorpion, palamnaeus swammerdami. J. Morphol., 144: 1-10.
PubMed - Shigenaga, T., Y. Takayenoki, S. Kawasaki, N. Seki and T. Muta et al., 1993. Separation of large and small granules from horseshoe crab (Tachypleus tridentatus) hemocytes and characterization of their components. J. Biochem., 114: 307-316.
PubMedDirect Link - Synder, G.K. and C.P. Mangum, 1982. The Relationship Between the Capacity for Oxygen Transport, Size, Shape and Aggregation State of an Extracellular Oxygen Carrier. In: Physiology and Biology of Horseshoe Crab: Studies on Normal and Environmentally Stressed Animlas, Bonaventura, J., C. Bonaventura and S. Tesh (Eds.). A.R. Liss Inc., New York, USA., ISBN: 9780845100813, pp 173-188.
- Tincu, J.A. and S.W. Taylor, 2004. Antimicrobial peptides from marine invertebrate. Antimicrob. Agents Chemother., 48: 3645-3654.
CrossRef - Toh, Y., F. Amizutani, T. Tokunaga and S.I. Muta, 1991. Morphology of the granular hemocytes of the Japanese horseshoe crab Tachypleus tridentatus and immunocytochemical localization of clotting factors and antimicrobial substances. Biomed. Life Sci., 266: 137-147.
CrossRef