
Roushon Ara
Department of Aquaculture, Faculty of Agriculture, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
Aziz Arshad
Department of Aquaculture, Faculty of Agriculture, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
S.M.N. Amin
Department of Aquaculture, Faculty of Agriculture, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
A.G. Mazlan
School of Environmental and Natural Resource Sciences, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor, Malaysia
M. Aminur Rahman
Laboratory of Marine Science Biotechnology, Institute of Bioscience, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
Asian Journal of Animal and Veterinary Advances
Year: 2013 | Volume: 8 | Issue: 2 | Page No.: 266-274







ABSTRACT
Larval stage of marine fishes, habitat characteristics, nursery and feeding ecology has been discussed in this review. Identification of fish larvae is difficult and that is why limited research is available on biology of fish larvae in the Indo-Pacific coastal region. Estuaries play a vital role in energy transfer between a river and a sea, which is especially important for many commercial coastal fishes whose larvae and juveniles are dependent on the estuary as a nursery and feeding grounds. Many coral reef fish species use mangrove and seagrass beds as nursery habitats. Diet composition of the family Clupeidae were made up of seven major categories to include phytoplankton, plant-like matter, debris, fragment of copepod, algae and unidentified food items. The most predominant food item in the gut of clupeids was phytoplankton (82.53%).
PDF Abstract XML References Citation
Received: July 02, 2012;
Accepted: July 31, 2012;
Published: February 11, 2013
How to cite this article
Roushon Ara, Aziz Arshad, S.M.N. Amin, A.G. Mazlan and M. Aminur Rahman, 2013. Feeding Ecology and Nursery of Marine Larval Fishes. Asian Journal of Animal and Veterinary Advances, 8: 266-274.
DOI: 10.3923/ajava.2013.266.274
URL: https://scialert.net/abstract/?doi=ajava.2013.266.274
DOI: 10.3923/ajava.2013.266.274
URL: https://scialert.net/abstract/?doi=ajava.2013.266.274
INTRODUCTION
Fish larval studies are often the best way to give information of great value to fishery biologists and managers of fisheries. These comprise: spawning grounds in space and time, determination of habitats used by fish during their larval phase. Due to the difficulty of identification, there is a limited knowledge on the biology of larvae of the Indo-Pacific coastal fishes (Leis, 1991).
Different fishes consume different types of food items and feeding habit of fishes varies with time also. Dietary preferences for the larval stages are therefore an important factor in the assessment of feeding conditions and larval chances of meetings food requirements (Robichaud-LeBlanc et al., 1997). Prey preferences of larval fishes are determined mainly based on the gut contents analysis. There is a limitation of empirical data on the diets of fish larvae in the wild (Humphries et al., 1999). The information about the specification of diet composition both qualitatively and quantitatively are described here can be applied in aquaculture development. Various studies have been carried out in terms of food and feeding habits of the adult fishes (Chrisfi et al., 2007; Dadzie et al., 2000; Jardas et al., 2007) but there have been very little information regarding the feeding habits of larval fishes in the wild (Grabowska and Grabowski, 2005; Kakareko et al., 2005). The present report summarizes the information of feeding ecology and nursery of marine fish larvae on the basis of present collected literatures.
THE FISH LARVAE
In majority of species with pelagic eggs occurring around the British Isles, the newly hatched larva is less than 4.0 mm long (Russell, 1976). The exceptions are mostly species with elongated larvae such as those of the clupeids. Many of the larvae of shore fish are however, considerably larger when they hatch. In the early larva (Fig. 1), the yolk sac on the anterior ventral side of the body is a prominent feature and it often occupies about half of the total body length. In those with a short incubation period, the eyes are not pigmented, the mouth is not functional and the anus is not open.
The larval stage after the yolk-sac absorption is conveniently divided into preflexion, flexion and postflexion stages (Fig. 2). The flexion stage is usually accompanied by rapid development of fin rays, changes in body shape, abilities in both swimming and feeding behaviors. During larval development, the eyes become fully pigmented, the mouth and the anus open.
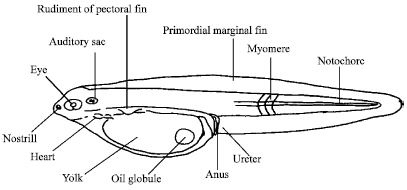 | |
| Fig. 1: | Diagrammatic representation of a newly hatched larva from a pelagic egg (Russell, 1976) |
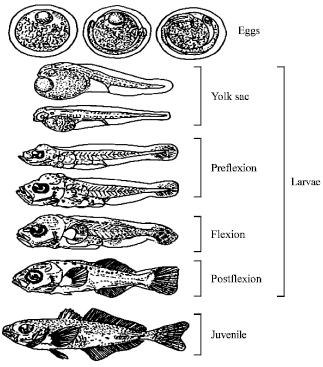 | |
| Fig. 2: | Early life stages of Trachurus symmetricus (Ahlstrom and Ball, 1954) |
The position of the anus may be a useful character for identification. It may be closed to the posterior end of the yolk sac or slightly far behind it. At the end of the larval stage the arrangement of the pigmentation has usually been changed, the pigment cells being moved into positions characteristic of the post larvae by which most species can be identified.
THE EGG STAGE
Identification of fish eggs is more complex than that of larvae because eggs have fewer characters than larvae (Fig. 2). For egg identification, the following characteristics are useful (Moser et al., 1984):
| • | Egg size | |
| • | Shape | |
| • | Size of perivitelline space | |
| • | Yolk diameter and size | |
| • | Pigment on the yolk and embryo | |
| • | Characters of the developing embryo: | |
| • | Relative rate of development of various body arts | |
| • | Body shape | |
| • | Number of somites | |
THE YOLK- SAC STAGE
The yolk-sac stage is a transitional phase between the egg and larval stage (Fig. 2). The following characteristics are used for identification of yolk-sac stage:
| • | Shapes of dorsal, caudal and anal fin folds and larval pectoral fins |
| • | Melanophore patterns moving from the dorsal surface to the lateral or ventral parts |
| • | Presence/absence and position of oil globules of yolk-sac larvae |
| • | Shape and relative size of the yolk-sac |
HABITAT OF FISH LARVAE
Estuarine habitat: An estuary is a region containing a volume of water of mixed origin derived partly from a discharging river system and partly from the adjacent sea; the region usually being partially enclosed by a land mass. Estuaries play an important role in energy transfer between the river and the sea, which is especially important for many commercial coastal fishes whose larvae and juveniles are dependent on the estuary as a nursery and feeding ground (Tzeng and Wang, 1992, 1993). The macro-fauna comprises two ecologically distinct grouping: those (mainly invertebrate) species permanently resident within an estuary and those (mainly vertebrate) species entering estuaries at high or low tide principally to feed (e.g., fish). The resident species of temperate regions are drawn mainly from within the polychaeta, the Mollusca (especially lamellibranches) and the Crustacea (especially isopods and amphipods). In tropical and subtropical areas, the dominant animals are gastropod molluscs and crabs of the family Ocypodidae and Grapsidae. The majority of the migratory fauna uses estuaries as feeding grounds. Many families of fish have representatives which are usually feed in estuaries during some of their life cycle stages, for example the Clupeidae (Barnes, 1984).
All estuaries, which drain incompletely as the tide ebbs, have characteristic plankton population. If this condition is not fulfilled, populations cannot be sustained and the benthic invertebrate populations which have planktonic larvae depend upon external water movements to bring recruits from outside colonies. Even when this condition is satisfied, a more variable salinity and temperature, varying degrees of turbidity and negative estuaries apart, a net loss of water to the open sea result in a special challenge to the inhabitants of an estuary (Perkins, 1974). Flagellates, diatoms and blue-green algae may all are abundant in and on estuarine mud. The plankton of estuaries is usually sparse in numbers of species, although, frequently rich in numbers of individuals. Diatoms (especially Skeletonema, Asterionella, Chaetoceros, Nitzschia, Thalassionema and Melosira) frequently dominate the phytoplankton but small dinoflagellates (including Gymnodinium and Gonyaulax), often show irregular and localized bloom throughout the year and in many estuaries Gymnodinium group of dinoflagellates is dominant (Barnes, 1984). Estuaries usually composed of relatively few species and usually dominated by one or two species of calanoid copepod (Bagnall, 1976).
Mangrove habitat: Mangrove habitat can be the best defining as the prop-root system and adjacent muddy bottoms areas such as lagoons, creeks and passages. Mangroves are the characteristics intertidal plant formations of sheltered tropical and subtropical coastlines (Saenger, 2002). The most restrictive and possible best definition of the mangrove habitat is the mangrove prop-root system and adjacent muddy bottom areas, such as lagoons, creeks and passages, that are derived from mangrove-induced deposition processes. The variability of physical conditions within mangroves-salinity, tidal exposure, turbidity and a complex prop root system-means they can support unique assemblages of flora and fauna (Nisbet, 2007). Heald and Odum (1970) stated it as widely accepted view that mangroves are primary nursery areas for commercially important fishes. Many coral reef fish species use mangrove and seagrass beds as nursery habitats (Mumby et al., 2004). Nisbet (2007) reported that while the importance of mangrove as a nursery area for fish and prawn species, has become a focus for research. The zooplankton of mangroves are a major component of the trophic structure and should be equally valued within the research (Beck et al., 2001). The role of zooplankton in mangroves is virtually unknown, despite their pivotal trophic role as intermediaries between alternative carbon sources (detritus) and higher consumers such as fish (McKinnon and Klumpp, 1998).
Seagrass habitat: A seagrass meadow is a highly productive and dynamic ecosystem; it ranks among the most productive in the ocean. Seagrass beds, macroalgal beds and sandy bottom areas are adjacent to the mangroves in most areas with interaction among these habitats being an important ecological feature of near shore tropical ecosystem (Ogden and Gladfelter, 1983). Among the forms spent only part of their life in the seagrass beds are several economically important fishes and shrimps. The seagrasses usually occur in enormous quantities and form dense beds which cover immense areas in the coastal waters of both temperate and tropical seas and in fact one of the most conspicuous communities of the shore. Besides, it is known that the seagrass beds indeed fulfill several functions such as stabilizing the bottom, providing a substrate for epiphytes and epizoa on bottoms otherwise unsuitable for these species and providing shelter and nursery ground for fishes, shrimps and other invertebrates (Den Hartog, 1977).
Nursery area of fish larvae: For demersal fishes, a nursery area should be an area of optimum habitat where larval fishes can settle and continue to grow and develop with a low mortality rate due to low risk of predation or starvation. The optimality of any habitat should be determined by a balance between predation and growth rate. For rapid growth in larval and juvenile fishes, there must be an adequate food supply. Larger larval fishes feed directly on the zooplankton assemblage. When larval demersal fishes settle (usually at the beginning of the juvenile stage), generally there is a switch to more benthic food sources. Due to high detritus levels in the mangroves, there is an array of benthic invertebrates available as food for these juvenile fishes (Austin and Austin, 1971).
Within the tropical reef ecosystem, the species diversity declines from coral reefs to mangroves to seagrass beds in that order (Abele, 1974; Kimmel, 1985). One explanation for this decrease in diversity is the decrease in spatial heterogeneity (i.e., shelter). Though direct evidence is lacking, the paucity of fishes on seagrass beds during the day when compared to those at night suggests strong selective pressures against inhabiting seagrass beds, during the day (Robblee and Zieman, 1984). Diurnal predators, such as jacks, snappers and sharks, feeding in seagrass beds can take advantage of the low relief to obtain prey fishes, which once detected have few means of escaping other to flee. The higher dimensionality of the mangrove prop-root system provides more opportunity for predator avoidance. Yet, there is less protection afforded by the mangrove prop-roots than the coral reef habitat due to the relative open nature of the roots.
Many coral reef fishes exhibit a separation of habits between the juvenile and adult stages. A benefit of habitat separation could be a reduced predation rate in juvenile habitats. Whereas settlement is in nearby habitats, such as seagrass beds and mangroves could provide an opportunity to develop to a size that might improve survival on the coral reef. These suggest that there could be an advantage for larval and juvenile coral reef fishes to use seagrass beds and mangroves as nursery grounds. Of the near shore tropical habitats, mangroves potentially provide both a good food source and a low predation, thus making them ideal nursery areas.
FOOD AND FEEDING HABITS
Feeding habits of larval fishes of the family Clupeidae (Actinopterygii: Clupeiformes) was studied from October 2007 to September 2008 in the estuary of river Pendas, Johor, Peninsular Malaysia (Ara et al., 2010). The diet composition were grouped into seven major categories of phytoplankton, plant like matter, debris, fragment of copepod, algae and unidentified food items. The most predominant food item in the gut of clupeids was phytoplankton (82.53%).
Diet composition and feeding habits of fish larvae from the family Gobiidae were investigated in seagrass beds of Sungai Pulai estuary, Johor Strait, Malaysia (Ara et al., 2011). Samples were collected monthly from December 2007 to May 2008. Stomach contents were analyzed in 88 larval specimens with size range between 2.0-4.0 mm in total length. Prey analyses of stomach contents identified 24 important items belonging to seven major diet groups: phytoplankton, zooplankton, algae, insects, plant-like matter, debris and unidentified matters. In term of percentage, dominant preys were phytoplankton (56.33%) and this was followed by plant matters (15.13), zooplankton (9.84%), algae (9.73%), debris (4.85%), decapods appendages (3.19%), unidentified matters (2.39%) and insects in that order (1.73%). The results of the study could be used to conclude that gobiid larvae are mainly herbivorous.
Diet composition of most common fish larvae of the family Sparidae was studied in the Sungai Pulai Seagrass bed of Gelang Patah, the South Western part of Johor, Peninsular Malaysia from December 2007 to May 2008 (Ara et al., 2009). Larvae were collected by subsurface towing of a bongo net. In situ hydrographic parameters were recorded during the sampling cruises. Stomachs were removed from a total of 80 sparidae specimens during the study period. Analysis of prey in the stomach showed 24 important food items belonging to 8 major groups: phytoplankton, zoology, algae, insect, plant like matter, decapoda, debris, fragment of copepod and unidentified food items. The predominant food item is found in the stomach were phytoplankton (60.85%). This was followed by algae (11.73%), zooplankton (9.35%), plant like matter (7.84%), debris (4.60%), insect (2.84%), unidentified food items (1.96%) and decapod appendages (0.82%). Among the food items examined, phytoplankton was ranked first by simple resultant index (60.85%). Therefore it could be concluded that the fish larvae of family Sparidae are mainly herbivorous.
It is believed that early life history stages of fishes are highly susceptible to starvation, which may enhance predation and both the starvation and predation together may be the overpowering causes of mortality (Hunter, 1981). These are important reasons for studying the food and feeding ecology of larval fishes (Brewer and Kleppe, 1986). One of the most important influences on the survival of fish larvae is the availability of suitable food, thus knowledge of their feeding behavior is necessary to understand the factors affecting the mortality of the larvae in the wild (Last, 1980; Sanchez-Velascoa, 1998).
Feeding strategies of fish larvae are complex. Moreover, they depend on the food availability in areas defined by mesoscale hydrographic structures (Sanchez-Velascoa, 1998). Some factors, such as the physiological and morphological features of the larvae and their habitat preferences, play important roles (Sanchez-Velascoa, 1998). Perception, recognition, capture and digestibility of prey influence the apparent selection of food items (Govoni et al., 1986). Some selected studies on diet composition and feeding habits of fish larvae are given below:
Contrasting feeding patterns among species of fish larvae from the tropical Andaman Sea were reported by Ostergaard et al. (2005). Feeding habits of tropical fish larvae were analyzed during a comparative study among the 4 species (Scorpaenodes sp., Carangoides sp., Acanthocepola sp. and Cynoglossus sp.). Gut contents of a total of 300 larvae were examined and compared with local zooplankton composition. They investigated morphological characteristics and their potential influence on larval feeding and prey preference.
Strong dietary relationships were identified between fishes and copepods in the same community of Chikugo estuary, Japan (Islam et al., 2006). They reported that Estuarine Turbidity Maximum (ETM) appear to have significant influence on the distribution and abundance of the oligohaline copepod, S. sinensis and this prey copepod appears to have strong influence on the fishes in the oligohaline regions. Most of the fishes were dispersed in the low saline upper estuary where they foraged on the single dominant copepod S. sinensis, which contributes to the majority of the copepod biomass of the estuary and thus appear to maintain nursery ground for fish. They concluded that the ETM-based copepod, S. sinensis plays a key role in survival and distribution of larvae and juvenile fishes in Chikugo estuary, Japan. Feeding ecology of Morone americana larvae were studied by Shoji et al. (2005) in the Chesapeake Bay estuarine turbidity maximum. They reported that Copepodites and adults of Eurytemora affinis were the predominant prey in guts of all length classes of larvae (3.2-9.8 mm standard length). Multiple regression analyses were conducted to evaluate the influence of several factors on feeding success by determining whether physical conditions (temperature, current velocity, salinity, turbidity and light) and prey concentrations explained variability in mean gut fullness. They accomplished those high concentrations of E. affinis, which enhanced encounter rates of white perch larvae with prey, may have been the most important factor determining feeding success in the estuarine turbidity maximum region. Govoni et al. (1986) studied the diet vertical distribution of fish larvae and their prey in near shore waters of southern California. They used bongo net for collecting ichthyoplankton during a winter cruise in Santa Monica Bay. Many large northern anchovy (≥12 mm in length) apparently migrated to the sea surface at night but large white croaker (≥6 mm in length) did not. Only 9% of northern anchovy larvae contained at least 1 food particle (i.e., 9% feeding incidence), compared to 76% for white croaker larvae. Early life stages of copepods were important foods for both fish species. Bivalve veligers and tintinnids observed frequently in the guts of white croaker but not in the northern anchovy. They concluded that little relationship was maintained between daytime feeding distribution of fish larvae and esculent microzooplankton, except the distribution of white croaker and bivalve veligers which were significantly correlated (p<0.05). Diet composition and feeding habits of fish larvae of two co-occurring species (Pisces: Callionymidae and Bothidae) were studied by Sanchez-Velascoa (1998) in the North-western Mediterranean. Sanchez-Velascoa (1998) mentioned that the dominant prey were Oithona sp. and Oncaea sp. Followed by less frequency, Paracalanus sp. copepodites. Copepod nauplii were not significant in the diet of these two species, although they were the most plentiful zooplankter in that area. As larvae of both species grew, the size and number of prey increased. This tendency was most obvious in Callionymus sp. larvae, which reflected greater prey diversity during all of their early ontogeny. An analysis of covariance showed that prey size increased with larval size in both species but the prey consumed by Callionymus sp. larvae was larger than that of A. laterna, corresponding to the larger mouth size. It is concluded that these co-occurring species have different feeding strategies.
CONCLUSION
Since the study on natural habitat, feeding ecology and diet composition of larval fishes have enormous importance in fishery biology the information gathered from this comprehensive review, would immensely be helpful towards the understanding of feed and feeding habit and nursery of many commercially important fishes, which will ultimately be useful for successful aquaculture management and biodiversity conservation.
ACKNOWLEDGMENTS
The research was supported by the research grant provided by the Ministry of Science, Technology and Innovation (MOSTI), Malaysia (Grant No. 5450247). Technical assistance and logistics were provided by the Universiti Putra Malaysia. The authors also acknowledge all the local fishermen for their assistance during field sampling.
REFERENCES
- Abele, L.G., 1974. Species diversity of decapods crustaceans in marine habitats. Ecology, 55: 156-161.
Direct Link - Ara, R., A. Arshad, N. Amrullah, S.M.N. Amin, S.K. Daud, A.A. Nor Azwady and A.G. Mazlan, 2009. Feeding habits and temporal variation of diet composition of fish larvae (Osteichthyes: Sparidae) in the Sungai Pulai seagrass bed, Johore, Peninsular Malaysia. J. Biol. Sci., 9: 445-451.
CrossRefDirect Link - Ara, R., A. Arshad, S.M.N. Amin, S.K. Daud, J.S. Bujang and M.A. Ghaffar, 2010. Feeding habits of larval fishes of the family Gobiidae (Actinopterygii: Perciformes) in seagrass beds of Sungai Pulai estuary, Johor Strait, Malaysia. Coastal Mar. Sci., 34: 123-128.
Direct Link - Ara, R., A. Arshad, L. Musa, S.M.N. Amin and P. Kuppan, 2011. Feeding habits of larval fishes of the family Clupeidae (Actinopterygii: Clupeiformes) in the Estuary of River Pendas, Johor, Malaysia. J. Fish. Aquatic Sci., 6: 816-821.
CrossRefDirect Link - Beck, M.W., K.L. Heck Jr., K.W. Able, D.L. Childers and D.B. Eggleston et al., 2001. The identification, conservation and management of estuarine and marine nurseries for fish and invertebrates. BioScience, 51: 633-642.
CrossRefDirect Link - Brewer, G.D. and G.S. Kleppe, 1986. Diel vertical distribution of fish larvae and their prey in nearshore waters of southern California. Mar. Ecol. Prog. Ser., 27: 217-226.
Direct Link - Chrisafi, P., P. Kaspiris and G. Katselis, 2007. Feeding habits of sand smelt (Atherina boyeri, Risso 1810) in Tichonis Lake (Western Greece). J. Applied Ichthyol., 23: 209-214.
CrossRef - Dadzie, S., F. Abou-Seedo and E. Al-Qattar, 2000. The food and feeding habits of the silver pomfret, Pampus argenteus (Euphrasen), in Kuwait waters. J. Applied Ichthyol., 16: 61-67.
CrossRefDirect Link - Humphries, P., A.J. King and J.D. Koehn, 1999. Fish, flows and floodplains: Links between freshwater fish and their environment in the Murray-Darling Rive system, Australia. Environ. Biol. Fish., 56: 129-151.
CrossRef - Islam, S., M. Hibino and M. Tanaka, 2006. Distribution and the diet of larval and juvenile fishes: Influences of salinity gradient and the turbidity maximum in a temperate estuary in upper Ariake Bay, Japan. Estuarine Costal Shelf Sci., 68: 62-74.
CrossRefDirect Link - Kakareko, T., J. Zbikowski and J. Zytkowicz, 2005. Diet partitioningin summer of two syntopic neogobiids from two differenthabitats of the lower Vistula River, Poland. J. Applied Ichthyol., 21: 292-295.
CrossRef - McKinnon, A.D. and D.W. Klumpp, 1998. Mangrove zooplankton in North Queensland, Australia. Hydrobiologia, 362: 127-143.
CrossRef - Mumby, P., A. Edwads, E.A. Gonzalez, K. Lindeman and P. Blacwell et al., 2004. Mangroves enhance the biomass of coral reef fish communities in the Caribbean. Nature, 427: 533-536.
Direct Link - Ostergaard, P., P. Munk and V. Janekarn, 2005. Contrasting feeding patterns among species of fish larvae from the tropical Andaman Sea. Mar. Biol., 146: 595-606.
Direct Link - Robblee, M.B. and J.C. Zieman, 1984. Diel variation in the fish fauna of a tropical seagrass feeding ground. Bull. Mar. Sci., 34: 335-345.
Direct Link - Robichaud-LeBlanc, K.A., S.C. Courtenay and J.M. Hanson, 1997. Ontogenic diet shifts in age of striped bass, Morone saxatilis, from the Miramichi River estuary, Gulf of St. Lawrence. Can. J. Zool., 75: 1300-1309.
Direct Link - Sanchez-Velascoa, L., 1998. Diet composition and feeding habits of fish larvae of two co-occurring species (Pisces: Callionymidae and Bothidae) in the north-western Mediterranean. ICES J. Mar. Sci., 55: 299-308.
CrossRefDirect Link - Shoji, J., E.W. North and E.D. Houde, 2005. The feeding ecology of Morone americana larvae in the Chesapeake Bay estuarine turbidity maximum: The influence of physical conditions and prey concentrations. J. Fish Biol., 66: 1328-1341.
CrossRef - Tzeng, W.N. and Y.T. Wang, 1992. Structure, composition and seasonal dynamics of the larvae and juvenile fish community in the mangrove estuary of Tanshui River, Taiwan. Mar. Biol., 113: 481-490.
CrossRef - Tzeng, W.N. and Y.T. Wang, 1993. Hydrography and distribution dynamics of larval and juvenile fishes in the coastal waters of the Tanshui River estuary, Taiwan, with reference to estuarine larval transport. Mar. Biol., 116: 205-217.
CrossRef