
R. Ara
Department of Aquaculture, Faculty of Agriculture, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
S.M.N. Amin
Department of Aquaculture, Faculty of Agriculture, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
A.G. Mazlan
School of Environmental and Natural Resource Sciences, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor, Malaysia
A. Arshad
Department of Aquaculture, Faculty of Agriculture, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
Asian Journal of Animal and Veterinary Advances
Year: 2013 | Volume: 8 | Issue: 2 | Page No.: 247-256







ABSTRACT
Morphometric data of six dominant families of fish larvae were examined to observe the degree of similarity among the six families. The experiment was conducted in the seagrass-mangrove ecosystems of Gelang Patah, Johor Strait, Peninsular Malaysia. Samples of fish larvae were collected by using bongo net through 30 min subsurface tow. The six dominant families (Terapontidae, Gobiidae, Sillaginidae, Nemipteridae, Blenniidae and Leiognathidae) were selected for the morphometric analysis using one-way ANOVA by SPSS and PRIMER 5 software for cluster analysis. All the morphometric characters, total length, standard length, body depth, snout length, head length, eye diameter and pre anal length of the six dominant families of the order Perciformes were found to be significantly different (p<0.05). The study is formed a basis for further extensive research to be carried out on larval fishes in Malaysian waters.
PDF Abstract XML References Citation
Received: July 02, 2012;
Accepted: August 04, 2012;
Published: February 11, 2013
How to cite this article
R. Ara, S.M.N. Amin, A.G. Mazlan and A. Arshad, 2013. Morphometric Variation Among Six Families of Larval Fishes in the Seagrass-Mangrove Ecosystem of Gelang Patah, Johor, Malaysia. Asian Journal of Animal and Veterinary Advances, 8: 247-256.
DOI: 10.3923/ajava.2013.247.256
URL: https://scialert.net/abstract/?doi=ajava.2013.247.256
DOI: 10.3923/ajava.2013.247.256
URL: https://scialert.net/abstract/?doi=ajava.2013.247.256
INTRODUCTION
In majority of species with pelagic eggs occurring around the British Isles, the newly hatched larvae is less than 4.0 mm long (Russell, 1976). The larval stage after the yolk-sac absorption is conveniently divided into preflexion, flexion and postflexion stages. Ecologically, larvae and adult are often entirely different and can be considered as different ecospecies (Leis and Carson-Ewart, 2000). Various fish species especially their larval and juvenile stage inhabit the seagrass beds for their reproduction and feeding. Merambong Shoal seagrass beds are among the most dense seagrass ecosystem in Peninsular Malaysia (Ara et al., 2011c). Seagrass beds and mangrove are the place, where species are known to use the habitats during their juvenile life stage (Nagelkerken et al., 2000). They stay there for at least a few months, as these habitats provide high abundance of foods and low predation pressure (Huijibers et al., 2008). The fish and its habitats should be safe in national balances. For this perspective, P. anatolicus is common and rare fish on the global scale (Sasi, 2011).
Larvae are often amazing morphologically change from the adult (Leis and Rennis, 1983). In measuring the discreteness and relationship among various taxa, morphological characteristics are commonly used and have long been used to identify various stocks of fish. As a first step in investigating the structure of species, the morphometric analyses can be used (Daud et al., 2005). It is often the best method for obtaining the desired information by means of statistical analysis (Gouch, 1984).
Measuring discreteness and relationships among stocks for morphometric characters are powerful implements (Ihssen et al., 1981; Melvin et al., 1992). Although, the feeding biology of major adult fishes in the seagrass have been studied in the Peninsular Malaysia (Jimmy, 2007), there is no report concerning morphometric analysis of fish larvae in Malaysia so far. Therefore, the focus on morphometric variation among the families of fish larvae is a new attempt in this direction.
MATERIALS AND METHODS
Study site and sampling: The study was conducted in seagrass, mangrove and estuarine ecosystem of Gelang Patah, Johor Strait Peninsular Malaysia (Fig. 1). Five sampling stations were selected along the axis of Pendas river estuary and Johor Strait. Among them, two stations were established within the river estuary (S1 and S2), two were outside the river mouth along the Strait (S3 and S4) and the other one was in open sea (S5). The specific location of the sampling stations were as S1 (N 01°23.345’; E 103°36.741’), upper estuary; S (N 01°22.79’; E 103°38.140’), middle estuary; S3 (N 01°21.597’; E 103°37.491’), lower estuary; S4 (N 01°19.414’; E 103°35.628’), Merambong seagrass beds and S5 (N 01°18.799’; E 103°35.246’), outside seagrass areas (open sea).
 | |
| Fig. 1: | Geographical location of the sampling sites in the seagrass-mangrove ecosystem of Gelang Patah, Johor Strait, Peninsular Malaysia |
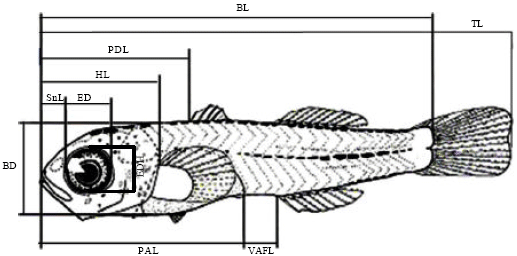 | |
| Fig. 2: | Diagrammatic representation of measurements total length (TL), body length (BL), body depth (BD), snout length (SnL), head length (HL), eye diameter (at vertical axis) (ED), eye diameter (at horizontal axis) (EDH), pre anal length (PAL) and vent to anal-fin length (VAFL) taken on specimens (Leis and Carson-Ewart, 2000) |
Sampling stations were approximately 1 km apart from each other (Fig. 1).
Sample processing: Samples of fish larvae were collected by using Bongo net through 30 min subsurface tow from each station. Samples were immediately fixed in 5% formalin and transported to the laboratory. In the laboratory, fish larvae were sorted from the rest of the zooplankton and were preserved in 75% alcohol. Individuals of fish larvae were identified to the family level using the appropriate literatures (Leis and Carson-Ewart, 2000; Russell, 1976; Okiyama, 1988; Ghaffar et al., 2010).
Data measurement: In total of 252 specimens in various sizes ranging from 1.08-6.05 mm were used for morphometric measurements. Eight selected morphometric characteristics, as shown in Fig. 2 were measured using the KEYENCE Digital microscope (VHX-500) for each sample. The morphometric characters measured include the Total Length (TL), Standard Length (SL), Body Depth (BD), Snout Length (SnL), Head Length (HL), eye diameter (at vertical axis) (EDV), eye diameter (at horizontal axis) (EDH) and pre anal length (P anal L).
Data analysis: The morphometric data were analyzed using one-way ANOVA by SPSS software and PRIMER 5 software for cluster analysis.
RESULTS
Morphometric description
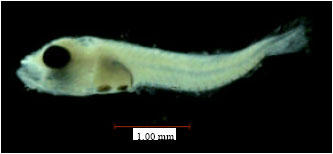
Gobiidae: The body was moderate to elongated with the total length that ranged from 1.08 to 5.58 mm with an average of 2.35 (±0.63) mm. The gut was straight or gently curved but not compact and some samples show ‘S’- shaped of gut (Fig. 3). These larvae were lightly pigmented. Some samples show the pigmented at ventral of the gut or the ventral midline of the tail or both.
 | |
| Fig. 3: | Photograph of a Gobiidae larva collected from the seagrass-mangrove ecosystem of Gelang Patah, Johor, Peninsular Malaysia |
 | |
| Fig. 4: | Photograph of a Terapontidae larva collected from the seagrass-mangrove ecosystem of Gelang Patah, Johor, Peninsular Malaysia |
 | |
| Fig. 5: | Photograph of a Sillaginidae larva collected from the seagrass-mangrove ecosystem of Gelang Patah, Johor, Peninsular Malaysia |
Terapontidae: The body of Terapontidae larva was elongated. The total length of the larvae was 1.47-4.78 mm with the average standard length of 2.76 mm (±0.50). The gut was triangularly coiled and compact and the snout was rounded. The head length was moderate with the range from 0.12 to 1.52 mm and the average was 0.78 mm. Body was lightly pigmented or sometimes no pigment was observed (Fig. 4). The pigment was observed at ventral of gut.
Sillaginidae: The body was elongated which the range of TL between 2.00 to 6.05 mm with an average of 3.61 mm. The gut was coiled. The snout was pointed and slightly concave in profile (Fig. 5). The head length was 0.15-0.50 mm. These larvae were lightly pigmented and characterized by a single row of melanophores along the ventral midline of the trunk and tail (Fig. 5).
Blenniidae: The total length of the family was moderate, 1.80-3.43 mm with an average of 2.44 mm. The gut was coiled, relatively short to body length and do not reach the mid body (Fig. 6).
 | |
| Fig. 6: | Photograph of a Blenniidae larva collected from the seagrass-mangrove ecosystem of Gelang Patah, Johor, Peninsular Malaysia |
 | |
| Fig. 7: | Photograph of a Nemipteridae larva collected from the seagrass-mangrove ecosystem of Gelang Patah, Johor, Peninsular Malaysia |
The snout was rounded and relatively short to head length (0.47-0.97 mm). The mouth was reaches of mid eye. No dorsal or anal fin was observed among the sample during the observation but the pectoral fin can be observed. The larvae were heavily pigmented. The pigment was observed on dorsal surface of the gut or at the pectoral fin (Fig. 6).
Nemipteridae: Larvae were deep and compressed. Body depth of this family was 0.52-0.94 mm with on average of 0.70 mm and the standard length measured range from 1.75 to 3.45 mm with the average standard length of 2.38 mm. The gut was tightly coiled, triangular coiled and relatively short to body length. Lightly pigmented or sometimes no pigment was observed. The pigment was observed on nearly dorsal surface of the gut or ventrally on the gut (Fig. 7).
Leiognathidae: Larvae were deep and compressed. Total length was 1.55 to 2.21 mm. The body depth of this family was 0.54-0.74 mm with an average was 0.64 mm and the standard length measured range from 1.16 to 2.01 mm with the average standard length was 1.63 mm. The gut was compactly coiled. The head was large in size with spine. The larvae were lightly pigmented. The pigment was observed on head region. The special features of this larva include the pre-opercula spine and serrate supra-occipital (Fig. 8).
Morphometric variation: The range and Mean±Standard Deviation (SD) values of the morphometric characters for the six dominant families from the same order Perciformes are presented in Table 1. The ANOVA showed that the mean differences of all characteristics are highly significant (p<0.05).
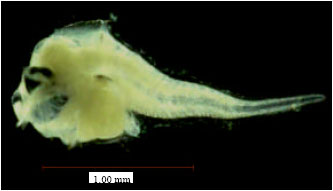 | |
| Fig. 8: | Photograph of a Leiognathidae larva collected from the seagrass-mangrove ecosystem of Gelang Patah, Johor, Peninsular Malaysia |
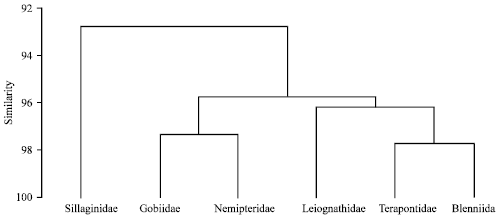 | |
| Fig. 9: | Dendrogram of six dominant families of fish larvae based on morphometric characters |
| Table 1: | Analysis of each morphometric characters (mm) of six dominant families |
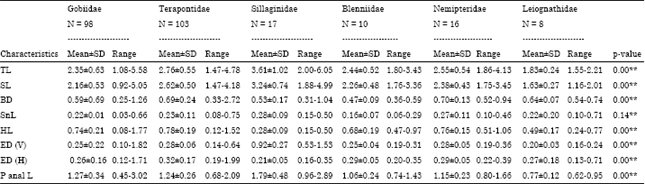 | |
| TL: Total length, SL: Standard length, BD: Body depth, SnL: Snout length, HL: Head length, ED (V): Eye diameter (at vertical axis), ED (H): Eye diameter (at horizontal axis), P anal L: Pre anal length, **Highly significant at 5% level (p<0.05) | |
Dendrogram based on morphometric characters showed five clusters amongst the populations (Fig. 9). The similarity matrix indicated that there were about 92-94% similarity between Sillaginidae and from other families. Second cluster, it contains families of Gobiidae, Leiognathidae, Nemipteridae, Terapontidae and Blenniidae. Third clusters contains family of Leiognathidae, Terapontidae and Blenniidae. Blenniidae and Sparidae fall under the same group, same as Gobiidae and Nemipteridae. The highest morphometric resemblance was observed between Blenniidae and Terapontidae with 97.73% of similarity (Fig. 9). While, the lowest morphometric resembles was observed between Sillaginidae and Gobiidae with 91.36% of similarity.
DISCUSSION
To describe the morphological features of fish larvae are often difficult. Firstly, some characteristics that are vital in describing the morphology of the larvae, whether they are not fully developed or broken such as fin ray and head spines. For further description, the shape and length of the gut are considered. Other characteristics that can be seen and described during the research works are size of mouth, shape of snout and head. Moreover, the characteristic such as fin ray and teeth cannot be fully ascertained because of the limitation such us broken appendages, unconfirmed taxonomic characteristic or not fully developed. These were encountered in most families except Blenniidae. As Blenniidae, the pectoral fins are well formed and this is the distinguished characteristic of this family.
Family Blenniidae are characterized by their short coiled gut and heavily pigmented, rounded head, moderate to large preopercular spine, moderately sized teeth and early development of the pectoral fin (Watson, 1983; Leis and Carson-Ewart, 2000). This is the most abundant family in the seagrass-mangrove ecosystem of Gelang Patah, Johor Strait. It is found in all the stations during the study period. It is previously reported from Sarawak and Sabah (Blaber et al., 1997), Indonesia (Soewito and Schalk, 1990), Thailand (Janekarn and Boonruang, 1986) and Taiwan (Tzeng and Wang, 1992).
Fish larvae of family Gobiidae are characterized by elongated and depth body, triangular gut, rounded and large eye, melanophores are present on the gut, ventral and dorsal of the tail. All characteristics are well agreed with the (Leis and Carson-Ewart, 2000). It is one of the dominant families in the study area. Gobiidae larvae are available in almost all estuaries in the Sarawak and Sabah (Blaber et al., 1997). It is also reported in studies from Thailand (Janekarn and Boonruang, 1986), Indonesia (Soewito and Schalk, 1990) and Taiwan (Tzeng and Wang, 1992).
Fish larvae family Leiognathidae are previously reported from Taiwan (Tzeng and Wang, 1992), Indonesia (Soewito and Schalk, 1990), Thailand (Janekarn and Boonruang, 1986) and Australia (Leis and Carson-Ewart, 2000). Larvae were easily distinguished by their deep and compressed body. The head is large, with spine and the pre operculum spines are present (Janekarn and Boonruang, 1986). It is found S3 to S5 from the study area. However, this family adult is previously reported (Jimmy, 2007) from the same region and (Yaakup et al., 2006) from Terengganu, Peninsular Malaysia.
Fish larvae of Nemipteridae are identified by their compressed body, triangular gut, large head with a round snout and melanophores are on dorsal surface of the gut. All these characteristics are similar with Leis and Carson-Ewart (2000). It is found in all the investigated areas during the study period.
Larvae Sillaginidae is recognized by an elongated body, straight gut, head is moderate to elongate and snout pointed which are similar with the description of Leis and Carson-Ewart (2000). This family is observed in all the stations (S1 to S5) of seagrass-mangrove ecosystem of Gelang Patah, Johor Strait, Peninsular Malaysia.
Family Terapontidae is one of the dominant fishes in the seagrass-mangrove ecosystem of Gelang Patah, Johor Strait. It is observed in the upper estuary (S1), lower estuary (S3), seagrass beds (S4) and outside of seagrass beds (S5) during the study period.
There is no report regarding morphometric variation among the families of fish larvae. There are some report on feeding (Ara et al., 2009, 2010 and 2011a) and some on composition (Ara et al., 2011b) from the study areas. However, most previous works on the morphometric and meristic variation were among species level of fish (Daud et al., 2005; Quilang et al., 2007) and shrimp (Amin et al., 2010). The one way ANOVA test is used by the most researchers to investigate the morphometric characteristic among the species whether it differs significantly or not. According to the ANOVA test, among of the six families, all the characteristics of the fish larvae are significantly different (p<0.05). This revealed that morphometric characteristic of fish larvae, amongst the six families is not similar and they are differing. Therefore, it could be concluded that all the families belongs o order Perciformes are different from each other.
CONCLUSIONS
All the morphometric characters, total length, standard length, body depth, snout length, head length, eye diameter, eye diameter and pre anal length of the six dominant families of the order Perciformes were found to be significantly different (p<0.05). Further details study on systematic ecology and biology of the fish larvae are needed from other geographical locations in Malaysia to gather more information for sustainable management of fishery resources.
ACKNOWLEDGMENTS
The research was supported by the research grant provided by the Ministry of Science, Technology and Innovation (MOSTI), Malaysia (Grant No. 5450247). Technical assistance and logistics were provided by the Universiti Putra Malaysia. The authors also acknowledge all the local fishermen for their assistance during field sampling.
REFERENCES
- Amin, S.M.N., A. Arshad, N.H. Ismail, M.H. Idris, J.S. Bujang and S.S. Siraj, 2010. Morphometric variation among the three species of genus Acetes (Decapoda: Sergestidae) in the coastal waters of Malacca, Peninsular Malaysia. Pertanika J. Trop. Agric. Sci., 33: 341-347.
Direct Link - Ara, R., A. Arshad, S.M.N. Amin, S.K. Daud and M.A. Ghaffar, 2011. Environment and diversity of ichthyoplankton in the seagrass beds of sungai pulai estuary, Johor, Peninsular Malaysia. J. Food Agric. Environ., 9: 733-738.
Direct Link - Ara, R., A. Arshad, L. Musa, S.M.N. Amin and P. Kuppan, 2011. Feeding habits of larval fishes of the family Clupeidae (Actinopterygii: Clupeiformes) in the Estuary of River Pendas, Johor, Malaysia. J. Fish. Aquatic Sci., 6: 816-821.
CrossRefDirect Link - Ara, R., A. Arshad, S.M.N. Amin, S.K. Daud, J.S. Bujang and M.A. Ghaffar, 2010. Feeding habits of larval fishes of the family Gobiidae (Actinopterygii: Perciformes) in seagrass beds of Sungai Pulai estuary, Johor Strait, Malaysia. Coastal Mar. Sci., 34: 123-128.
Direct Link - Ara, R., A. Arshad, N. Amrullah, S.M.N. Amin, S.K. Daud, A.A. Nor Azwady and A.G. Mazlan, 2009. Feeding habits and temporal variation of diet composition of fish larvae (Osteichthyes: Sparidae) in the Sungai Pulai seagrass bed, Johore, Peninsular Malaysia. J. Biol. Sci., 9: 445-451.
CrossRefDirect Link - Ara, R., A. Arshad, S.M.N. Amin, S.K. Daud and A.G. Mazlan, 2011. Comparison of larval fish density between seagrass beds and outside seagrass beds of the Southwestern Johor, Peninsular Malaysia. J. Fish. Aquatic Sci., 6: 795-801.
CrossRefDirect Link - Blaber, S.J.M., M.J. Farmer, D.A. Milton, J. Pang, O. Boon-Teck and P. Wong, 1997. The ichthyoplankton of selected estuaries in Sarawak and Sabah: Composition, distribution and habitat affinities. Estuarine Coastal Shelf Sci., 45: 197-208.
CrossRefDirect Link - Huijibers, C.M., E.M. Molle and I. Nagelkerken, 2008. Post-larval French grunts (Haemulon flavolineatum) distinguish between seagrass, mangrove and coral reef water: Implications for recognition of potential nursery habitats. Fish. Res., 357: 134-139.
CrossRef - Melvin, G.D., M.J. Dadswell and J.A. McKenzie, 1992. Usefulness of meristic and morphometric characters in discriminating populations of American shad (Alosa sapidissima) (Ostreichthyes:Clupeidae) inhabiting a marine environment. Can. J. Fish. Aquat. Sci., 46: 266-280.
CrossRefDirect Link - Nagelkerken, I., G. Van Der Velde, M.W. Gorissen, G.J. Meijer, T. van't Hof and C. Den Hartog, 2000. Importance of mangroves, seagrass beds and the shallow coral reef as a nursery for important coral reef fishes, using a visual census technique. Estuarine Coastal Shelf Sci., 51: 31-44.
CrossRef - Quilang, J.P., Z.U. Basiao, R.C. Pagulayan, R.R. Roderos and E.B. Barrios, 2007. Meristic and morphometric variation in the silver perch, Leiopotherapon plumbeus (Kner, 1864), from three lakes in the Philippines. J. Appl. Ichthyol., 23: 561-567.
CrossRef - Tzeng, W.N. and Y.T. Wang, 1992. Structure, composition and seasonal dynamics of the larvae and juvenile fish community in the mangrove estuary of Tanshui River, Taiwan. Mar. Biol., 113: 481-490.
CrossRef