
Ehsan Ramezani-Fard
Department of Aquaculture, Faculty of Agriculture, Universiti Putra Malaysia, 43400, Serdang, Selangor, Malaysia
Mohd Salleh Kamarudin
Department of Aquaculture, Faculty of Agriculture, Universiti Putra Malaysia, 43400, Serdang, Selangor, Malaysia
Sharr Azni Harmin
Center for Land and Aquatic Technology, Faculty of Science and Biotechnology, Universiti Selangor, 45600 Bestari Jaya, Selangor, Malaysia
Asian Journal of Animal and Veterinary Advances
Year: 2013 | Volume: 8 | Issue: 2 | Page No.: 205-215







ABSTRACT
Production of large amounts of fish eggs with high quality is one of the major goals in aquaculture industry. All reproductive processes in fish are regulated by endocrine system. Numerous studies have been carried out in recent years in order to develop or improve egg production protocols. This study provides a summary of some recent advances regarding fish oocyte differentiation, maturation and ovulation stages. The oocyte growth stages including first and second growth stages, veitellogenesis and final growth stage and maturation are briefly explained. Recent advances on the hormonal systems which control the reproductive process and oocyte development are also highlighted.
PDF Abstract XML References Citation
Received: July 02, 2012;
Accepted: July 24, 2012;
Published: February 11, 2013
How to cite this article
Ehsan Ramezani-Fard, Mohd Salleh Kamarudin and Sharr Azni Harmin, 2013. Endocrine Control of Oogenesis in Teleosts. Asian Journal of Animal and Veterinary Advances, 8: 205-215.
DOI: 10.3923/ajava.2013.205.215
URL: https://scialert.net/abstract/?doi=ajava.2013.205.215
DOI: 10.3923/ajava.2013.205.215
URL: https://scialert.net/abstract/?doi=ajava.2013.205.215
INTRODUCTION
Production of a high amount of fish eggs with high quality is one of the major goals in aquaculture industry. All reproductive processes, in fish as in other vertebrates, are regulated by a proper balance and interplay between the hormones of the hypothalamus, anterior pituitary and gonads which is classically referred to as the hypothalamo-hypophyseal-gonadal axis (Bhardwaj et al., 2012). Gonadotropin-releasing Hormones (GnRH) are released by hypothalamus while gonadotropins (Follicle-stimulating Hormone-FSH and Luteinizing Hormone-LH) are secreted by pituitary gland. Major hormones of reproduction, such as LH and FSH, directly control many aspects of gonadal development and function across vertebrates (Levavi-Sivan et al., 2010). Numerous studies have been carried out in recent years in order to develop or improve egg production protocols (Harmin and Crim, 1992; Muntaziana et al., 2011b; Zalina et al., 2011). However, the dynamic processes associated with oogenesis are not fully understood yet. Oogenesis is the process of creation of an egg in the female body and it involves the various stages and complicated regulatory mechanism. Oogenesis always starts with the differentiation of germ-line stem cells to generate a cyst of 16 cells that one of them will become the oocyte and the remaining 15 supply the oocyte with materials (Tavosanis and Gonzalez, 2003). Briefly, stem cells proliferate and undergo changes that turn them into oogonia. It is not clear whether true ooginal stem cells remain in the adult fish or not. Then, meiosis starts but the cell freezes at the diplotene stage of the first meiotic division and the previous oogonia turn into primary oocytes. As the primary oocytes are arrested at the first meiotic division, their growth begins (Babin et al., 2007). After final stage of the growth period, the follicular layers start collapsing and ovulation takes place. At this stage, the female gamete is ready for sperm binding and fertilization. It is clear now that oocyte maturation process and vitellogenesis are completely influenced by the pituitary gonadotropins and sex steroids. This study, therefore, tries to provide a summary of some recent advances regarding fish oocyte differentiation, maturation and ovulation stages.
OOCYTE GROWTH STAGES
At the primary growth stage, the nucleus of oocyte disperses numbers of nucleoli at its periphery. These nucleoli produce large amounts of ribosomal RNA and mRNAs that encode proteins required for subsequent oocytes growth. Primary growth is characterized by a substantial increase of the cell size and by formation of Balbiani bodies. Balbiany bodies are cytoplasmic masses corresponding to various cellular organelles such as Golgi apparatus, endoplasmatic reticulum cisternae, multivesicular bodies and even lipid granule. However the follicular layers are still undifferentiated in this stage (Rocha and Rocha, 2006). Oogenesis growth in teleosts includes more than one stage. The secondary growth period begins with appearance of some vesicles called cortical alveoli. The numbers of these vesicles increase steadily until occupying almost the entire ooplasm (Abdalla and Cruz-Landim, 2003). Cortical alveoli contains of glycoproteins which is synthesized by the oocytes itself. Therefore, these vesicles sometimes have been named yolk vesicles or endogenous yolk (Selman and Wallace, 1989; Gonzalez De Canales et al., 1992). Since the cortical alveoli do not provide nutrients for the developing embryo, using of any terms including “yolk” may not be true. The content of the cortical alveoli is released to the egg surface after the cortical reaction at fertilization. This releasing leads to the restructuring of egg envelop proteins and forming the chorion. At the secondary growth stage, lipid droplet spread all through the cytoplasm (Kimaro, 2011). At the same time, the follicular layers begin to differentiate to granulosa and therca sublayers. Follicle cell layers are usually established by an inner well-defined stratum, named granulosa cell layer and one or two outer sub-layers of theca cells. Follicle cells surround the centered developing oocyte and once the oocyte starts its growing, the follicular layers change in order to support, nourish and regulate its development in a continuous manner (Rocha and Rocha, 2006).
The main bulk of the fully-grown oocyte is due to yolk accumulation from its precursors, vitellogenins, during third growth period. The yolk precursor proteins are synthesized in the liver and freed into blood in response to a hormonal stimulation controlled by endocrine system and will be explained further on. At this stage follicular layers are well developed and granulosa and theca cells are easily observable. Beside the veitellogenesis, the vitelline envelope begins its formation in the early section of this stage (Srijunngam et al., 2005; Gulsoy et al., 2006). At the end of vitellogenesis, the oocyte contains maternal mRNAs, proteins, lipids, carbohydrate, vitamins and hormones that are important for the proper development of the embryo and becomes competent to undergo fertilization.
Throughout the final growth period, the oocytes achieve its maximum dimension, while tends to lose their spherical shape and becomes slightly flattened. In the animal pole, on one of the compressed surfaces, the tunnel-shaped micropyle develops throughout the zona radiate. During this time, the Maturation-inducing Hormones (MIH) are largely produced (Degani et al., 1994). At this stage the first meiotic division resumes and the oocyte divides to two cells differing in size. The small cell (first polar body) degenerates and the large secondary oocyte is formed. Finally, the follicular layers start collapsing and ovulation takes place at the end of maturation process (Rocha and Rocha, 2006). At this stage, the female gamete is known as ovum which is haploid due to occurrence of the second meiotic division and the formation of second polar body. The second polar body also degenerates. The ovum is arrested again at the second meiotic metaphase and fertilization becomes possible at this time. Continuation of the second division and completion of the meiosis needs to the sperm binding. This kind of division into different stages is quite artificial to facilitate explaining of the progression. As oocytes development is a dynamic process, it is difficult to identify the beginning or ending of each event accurately.
ENDOCRINE CONTROL OF REPRODUCTION
Commonly, fish brain controls reproduction via the release of gonadotropin-releasing hormones (GnRHs). These decapeptides, produced in hypothalamus, seems to be essential for reproduction of all vertebrates. Presence of three variants of GnRHs in the brain of many perciform species such as striped bass (Hassin et al., 1998) and tilapia (Weber et al., 1997) has been confirmed. Each of these three forms of GnRHs has a restricted regional pattern in the brain and appears to be regulated by both hormones and environmental cues. The hypothalamus in osteichhyan fish synthesizes products that are transported to the pituitary by direct neuronal innervations. However this structure is not found in chondrichthyan. Therefore, in many (but not all) fish, neuronal processes, originated in the hypothalamus, penetrate into the pituitary and allow direct neural control of the pituitary function (Rocha and Rocha, 2006). The pituitary gland is composed of adenohypophysis which regulates gonadal function in fish and the neurohypophysis. The adenohypophysis in tetrapods is the place of synthesis, storage and releasing of at least eight peptid and protein hormones. Some of them including gonadotropins (GTH-I or FSH and GTH-II or LH) are identified in teleosts (Chyb et al., 1999). GTH-I and -II are dimeric glycoproteins consisting of a common α-subunit and a hormone specific β-subunit that confers biological activity. The αβ dimmer forms after transcription to become the active hormone (Garcia-Campayo et al., 2002). Similar sequences are detected among the α- and β-subunit of each species as well as β-subunit of different species. This finding suggested that all subunit molecules originate evolutionarily from a common ancestral gene (Takei and Loretz, 2006). Gonadotropin (GTH) discharge is stimulated by GnRH and is repressed by dopamine (Azuadi et al., 2011).
GTHs can act directly on the GnRH receptors in the hypothalamus to reduce or modify the GnRH release. Indeed, this mechanism of GTHs in the body can be likened to what a thermostat does for controlling of a room temperature. In addition to GnRHs, there are also other hypothalamic factors which participate in the regulation of gonadotropins. Some of these peptides are including Pituitary Adenylate Cyclase Activating (PACAP), neoropeptide-Y (NPY), galanin, endothelin, oxytocin, Orexin, Vasoactive Intestinal Polypeptide (VIP) and substance-P (Khazali and Behzadfar, 2009; Levavi-Sivan et al., 2010). It has been shown that NPY amplifies GTH-II release in Goldfish (Carassius auratus) by a direct act on the pituitary or by escalating GnRH release (Peng et al., 1993). It has also been reported that leptin acts on the hypothalamic-pituitary-ovarian axis in several farm animals to enhance GnRH secretion and ovarian function (Sejian et al., 2010).
GTH-I released by pituitary stimulates the production of sexual hormones such as testosterone (T) by the theca cells (Fig. 1). Testosterone undergoes aromatization and changes to 17β-estradiol (E2) in the granulosa cells. In a prompt response to the E2 stimulus, the liver produces vitellogenin and secrete into the blood, where they form a complex with Ca2+ (Nagahama, 1994). Vitellogenins are sequestered by the follicles to be incorporated into the oocyte as yolk.
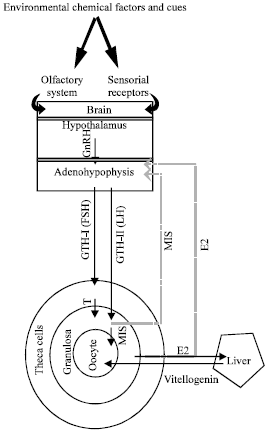 | |
| Fig. 1: | General aspects of endocrine reproduction control in females of fish, external stimuli influence the brain to release neurotransmitters, consequently, hypothalamic secretion induced and released GnRH stimulates the adenohypophysis for GTHs secreting |
There is a possible route for the vitellogenin passage from the blood capillaries, via the extracellular space among the theca cells, across the basal lamina, through the space between adjacent granulosa layer and across the canal in the chorion to make contact with the oolemma (Wallace and Selman, 1990).
Meiotic maturation of fish oocytes is induced by the action of MIH. The 17α,20β-dihydroxy-4-pregnen-3-one (17,20βP) and the 17α,20β,21-trihydroxy-4-pregnen-3-one (17,20β,21-P) are the most important MIH in many teleost species (Nagahama, 1997). Both of ovarian follicle cell layers (theca and granulosa) are involved in MIH synthesize. Theca layer produces 17α-hydroxyprogesterone (17-P) which is converted to MIH in the underneath granulosa cells. The presence of 20β-hydroxy steroid dehydrogenase (20β-HSD) is essential for this conversion. The stimulation of GTH-II leads to rapid expression of 20β-HSD mRNA transcripts in the granulosa cells during oocyte maturation (Nagahama, 1994; Nagahama, 1997; Yaron et al., 2003; Levi et al., 2008).
ENDOCRINE ASPECTS OF OOCYTE DEVELOPMENT
Pituitary injection increases ooginal proliferation and this increase is blocked by the gonadotropin antagonist (Tokarz, 1978; Dadzie and Hyder, 1976). Therefore, it is suggested that GTHs, either directly or indirectly via stimulation of ovarian mediators, increase ooginal proliferation (Lubzens et al., 2010). Recent studies showed that in addition to GTHs, some sex steroids may be effective on ooginal proliferation and inducing the first meiosis. “E2“ and “17,20βP” have been suggested for this purpose as the sexual hormone initiating the first meiotic division (Miura et al., 2007). The onset of meiosis has also been linked to the expression of insulin-like growth factor-I in the somatic cells and oocytes of tilapia (Berishivili et al., 2006).
Primary growth: As mentioned above, the primary growth of oocyte begins with freezing of cells at diploten stage of first meiosis and continues to early cortical alveoli stage. It is still debated whether primary oocyte development in teleosts is GTHs-independent or not. Few studies showed that hypophysectomi does not inhibit the primary growth of oocytes and follicles are able to proceed through development in the absence of GTHs (Pickford and Atz, 1957; Khoo, 1979). However, both FSHβ and LHβ transcripts and proteins have been detected in the primary and secondary oocytes of gilthead seabream, Sparus auratus (Wong and Zohar, 2004). Therefore it may be more accurate to use pituitary-independent term instead of GTHs-independent. Although, based on current very little information, it is not necessary to regard primary growth as pituitary-independent stage (Lubzens et al., 2010).
Secondary growth: Unlike the primary growth, transition of primary oocyte into secondary growth stage which corresponds to the accumulation of cortical alveoli is dependent on the presence of pituitary. Plasma levels of GTH-I increase during this stage and the hormonal level of circulation as well as expression of endocrine-related genes change. Consequently, stimulation of perinucleolar ovarian follicles by GTHs leads to the increase of E2 production (Swanson et al., 1989; Breton et al., 1998; Santos et al., 2001).
Campbell et al. (2006) showed that the degree of oocyte development in coho salmon is strongly affected by growth rate of broodstock before spawning season. Larger fish possess more advanced oocyte than smaller and slower growing fish. These authors demonstrated that synthesize of cortical alveoli is associated with the increase of plasma GTH-I, E2 and expression of transcripts encoding ovarian steroidogenic acute regulatory protein (StAR). They also showed that transcription rate of growth hormone receptors and somatolactin receptors in the ovary are reduced during that time. Moreover, accumulation of lipid in the oocyte of this species is related to the component of FSH-ovary axis such as plasma FSH, E2 and ovarian mRNA for GTHs receptors (Campbell et al., 2006).
Lubzens et al. (2010) reviewed available literature and pointed out to a notable increase of gene expression during secondary growth of oocyte. The expression of GTH-I receptor genes and the genes for some hormones made in granulose cells such as anti-mullerian hormone (AMH) highly increases during cortical alveoli stage and then decreases again during vitellogenesis. High potential of steroids production is another characteristic of cortical alveoli stage. This characteristic leads to the lipid accumulation and increase of oocyte diameter. Recently, the accumulation of cortical alveoli in zebrafish has also been associated with increased aromatase mRNA expression and E2 production (Kwok et al., 2005).
Veitellogenesis: From a hormonal point of view, vitellogenesis may be defined as the E2 induced hepatic synthesize of egg yolk precursors proteins, their secretion and transport via the circulation to the ovary and their uptake into developing oocyte. Early vitellogenesis is identified by increase of GTH-I and E2 and increased expression of ovarian GTH receptors (Breton et al., 1998; Kobayashi et al., 2008). E2 is considered as the most important steroids in inducing hepatic vitellogenin synthesis. It is difficult to measure vitellogenin level in normal male fish but the administration of E2 to juvenile and male teleost fish induced vitellogenin accumulation in their blood (Mommsen and Walsh, 1988; Muntaziana et al., 2011a). The detection of vitellogenin in male fish, therefore, is used as a marker to indicate exposure to oestrogenic substances and has been proposed to be a bio indicator for endocrine disruption (Ebrahimi, 2007; Ngamniyom and Panyarachun, 2011). In addition to E2, cortisol induces a rapid and transient transcription of vitellogenin mRNA in fish. However, injection of cortisol alone was not able to induce the expression of this messenger in rainbow trout (Babin et al., 2007). The involvement of androgens in vitellogenin synthesis has also been reported (Peyon et al., 1997; Kim et al., 2003). In vivo experiments showed that 17α-methyltestosterone and 5α-dihydrotestosterone treatment also induces vitellogenin in goldfish (Hori et al., 1979) and Gobius niger (Le Menn et al., 1980). While rainbow trout androgens or testosterone induce the expression of vitellogenin transcript, their oral administration in Oreochromis niloticus inhibited both the hepatic meesanger expression and the appearance of vitellogenin in the blood (Lazier et al., 1996). Despite the main role of E2 in inducing of hepatic vitellogenin synthesis, some other hormones such as growth hormone are also necessary for the induction (Kwon and Mugiya, 1994). In addition, it has been shown that recombinant sea bream parathyroid hormone related protein (PTHrP) has a potentiating effect on E2 stimulation of vitellogenin production by sea bream hepatocytes (Bevelander et al., 2006).
Vitellogenin level in blood is considered as a good biomarker to evaluate the presence of endocrine disrupting compounds (Kime et al., 1999). These compounds are able to conflict with the endocrine system and intend to prevent proper action of E2 and thus messing the reproduction of aquatic animals up. Therefore measuring of vitellogenin is suggested as a good indicator for monitoring antagonistic effect of endocrine disrupting compound.
To date, many studies confirmed that endocrine systems control the beginning of vitellogenesis in fish (Specker and Sullivan, 1994; Ohkubo et al., 2004) but the information regarding vitellogenesis termination is still insufficient. This lack is more prominent when it has been observed that during vitellogenesis termination, high peripheral E2 levels are still present (Babin et al., 2007). However, some studies have been expressed that acute increase in plasma GTH-II (LH) level, accompanied with increased expression of the LH receptor are the major endocrine events related with termination of vitellogenesis (Nagahama and Yamashita, 2008). Through this condition, LH turns the ovarian follicle steroidogenic pathway from the production of E2 during vitellogenesis to the production of MIH, although presence of high level of E2, even after vitellogenesis may be contrary with this assumption.
Maturation: Many studies showed an acute increase in plasma LH levels during the final growth stage (Nagahama and Yamashita, 2008). Binding of LH to its receptor on granulosa cells initiates the process of maturation of ovarian follicle which is marked by:
| • | Acquisition of oocyte maturational competence |
| • | Production of MIH |
| • | Production of Maturation-promoting Factor (MPF) and resumption of meiosis |
| • | Cytoplasmic maturation involving changes in the yolk proteins and lipids in order to provide diffusible nutrients for early embryogenesis |
Stimulation of meiosis resumption by MIH requires a high sensitivity of the oocyte to respond to the MIH. This is achieved by increasing the MIH receptors on the oocyte cell membrane as well as an increase in communication among the granulose cells and the oocyte through gap junctions. This process is known as maturational competence and may be induced by GTHs (Lubzens et al., 2010).
The process of producing MIH has been explained above. The MIH (17,20βP in most of the fish or 17,20β,21-P in some fish such as Atlantic croaker) induces synthesis of cyclin B in the ooplasm at the final growth satge of development. Cyclin B together with cdc2 kinase, form MPF (Yaron, 1995). Subsequently, MPF triggers the dissolution of the germinal vesicles and reinitiates meiosis. Upon egg activation, MPF is inactivated by degradation of cyclin B.
After the final growth stage and resumption of meiosis, the metaphase II oocyte is released from the follicle as a result of the ovulatory process. Arachidonic acid and its metabolites including prostaglandins F2α (PGF) has been known for years to induce ovulation in all fishes studied including carp, goldfish, yellow perch and Atlantic croaker (Yaron and Sivan, 2006). However, in some species, stimulation by MIH leads to both oocyte maturation and ovulation in vitro (Lubzens et al., 2010). Ovulation is associated with degradation of the follicular wall and formation of a rupture space throughout which the egg emerges. PGF and MIH enhance the activity of certain enzymes produced by the follicles for this purpose (Hsu and Goetz, 1992; Garczynski and Goetz, 1997; Patino et al., 2003). Although PGF stimulates the contraction of follicular wall, some of the other types of prostaglandins inhibit these contractions (Yaron and Sivan, 2006).
Atresia: Vitellogenic oocytes failed to undergo maturation and ovulation start a degenerative process called atresia. Atretic oocytes may be observed in the ovaries amongst the normal oocytes at any stage of development (Ramadan and EL-Halfawy, 2007). Different factors have been suggested as causing atresia including overproduction of oocytes due to environmental stress, hypophisectomy, starvation and inadequate hormone treatment (Guraya, 1986; Leino and Mccormik, 1997). Expression of GnRH and its receptors during gonadal regression resulted from lack of the sufficient level of GTHs might be involved in the induction of atresia (Habibi and Andreu-Vieyra, 2007). It is also proposed that atresia is related to increased activity of some of the protease enzyme (Lubzens et al., 2010). However, the mechanisms that initiate and regulate oocyte atresia in teleost fish are poorly known, especially at the molecular level.
REFERENCES
- Abdalla, F.C. and C.D. Cruz-Landim, 2003. Some histological and ultrastructural aspects of oogenesis in Piaractus mesopotamicus holmberg, 1887 (teleostei). Braz. J. Morphol. Sci., 20: 3-10.
Direct Link - Azuadi, N.M., S.S. Siraj, S.K. Daud, A. Christianus, S.A. Harmin, S. Sungan and R. Britin, 2011. Enhancing ovulation of Malaysian mahseer (Tor tambroides) in captivity by removal of dopaminergic inhibition. J. Fish. Aquatic Sci., 6: 740-750.
CrossRefDirect Link - Berishivili, G., H. D'Cotta, J.F. Baroiller, H. Segner and M. Reinecke, 2006. Differential expression of IGF-I mRNA and peptide in the male and female gonad during early development of a bony fish, the tilapia Oreochromis niloticus. Gen. Comp. Endocrinol., 146: 204-210.
CrossRef - Bevelander, G.S., X. Hang, W. Abbink, T. Spanings, A.V.M. Canario and G. Flik, 2006. PTHrP potentiating estradiol-induced vitellogenesis in sea bream (Sparus auratus, L.). Gen. Comp. Endocrinol., 149: 159-165.
CrossRef - Bhardwaj, A., V. Nayan, Parvati, Mamta and A.K. Gupta, 2012. Inhibin: A role for fecundity augmentation in farm animals. Asian J. Anim. Vet. Adv., 7: 771-789.
CrossRefDirect Link - Breton, B., M. Govoroun and T. Mikolajczyk, 1998. GTH I and GTH II secretion profiles during the reproductive cycle in female rainbow trout, relationship with pituitary responsiveness to GnRH-A stimulation. Gen. Comp. Endocrinol., 111: 38-50.
CrossRef - Campbell, B., J. Dickey, B. Beckman, G. Young, A. Pierce, H. Fukada and P. Swanson, 2006. Previtellogenic oocyte growth in salmon: Relationships among body growth, plasma insulin-like growth factor-1, estradiol-17beta, follicle-stimulating hormone and expression of ovarian genes for insulin-like growth factors, steroidogenic-acute regulatory protein and receptors for gonadotropins, growth hormone and somatolactin. Biol. Reprod., 75: 34-44.
CrossRefDirect Link - Chyb, J., T. Mikolajczyk and B. Breton, 1999. Post-ovulatory secretion of pituitary gonadotropins GtH I and GtH II in the rainbow trout (Oncorhynchus mykiss): Regulation by steroids and possible role of non-steroidal gonadal factors. J. Endocrinol., 163: 87-97.
CrossRef - Dadzie, S. and M. Hyder, 1976. Compensatory hypertrophy of the remaining ovary and the effects of methallibure in the unilaterally ovariectomized Tilapia aurea. Gen. Comp. Endocrinol., 29: 433-440.
CrossRef - Degani, G., E. Gal and J. Vaya, 1994. In vitro biosynthesis of steroids in ovary of asynchronic Trichogaster trichopterus (Pallus, 1770). Comp. Biochem. Physiol., 109: 715-723.
CrossRef - Ebrahimi, M., 2007. Vitellogenin assay by enzyme-linked immunosorbant assay as a biomarker of endocrine disruptor chemicals pollution. Pak. J. Biol. Sci., 10: 3109-3114.
CrossRefPubMedDirect Link - Garcia-Campayo, V., T.R. Kumar and I. Boime, 2002. Thyrotropin, follitropin, and chorionic gonadotropin expressed as a single multifunctional unit reveal remarkable permissiveness in receptor-ligand interactions. Endocrinology, 143: 3773-3778.
CrossRef - Garczynski, M.A. and F.W. Goetz, 1997. Molecular characterization of a ribonucleic acid transcript that is highly up-regulated at the time of ovulation in the brook trout (Salvelinus fontinalis) ovary. Biol. Reprod., 57: 856-864.
PubMed - Gonzalez De Canales, M.L., M. Blanco and M.C. Sarasquete, 1992. Carbohydrate and protein histochemistry during oogenesis in Halobatrachus didactylus (Schneider, 1801) from the bay of cadiz (Spain). Histochem. J., 24: 337-344.
CrossRef - Gulsoy, N., Y. Aytekin and R. Yuce, 2006. Changing of follicular epithelium during oogenesis in rainbow trout (Oncorhynchus mykiss, W.), studied by light and electron microscopy. Pak. J. Biol. Sci., 9: 935-939.
CrossRefDirect Link - Guraya, S.S., 1986. The cell and molecular biology of fish oogenesis. Monogr. Dev. Biol., 18: 1-223.
PubMed - Harmin, S.A. and L.W. Crim, 1992. Gonadotropic hormone-releasing hormone analog (GnRH-A) induced ovulation and spawning in female winter flounder, Pseudopleuronectes americanus (Walbaum). Aquaculture, 104: 375-390.
CrossRefDirect Link - Hori, S.H., T. Kodama and K. Tanahashi, 1979. Induction of vitellogenin synthesis in goldfish by massive doses of androgens. Gen. Comp. Endocrinol., 37: 306-320.
CrossRef - Hsu, S.Y. and F.W. Goetz, 1992. Angiotensins stimulate In vitro ovulation and contraction of brook trout (Salvelinus fontinalis) follicles. Fish Physiol. Biochem., 10: 277-282.
CrossRef - Khazali, H. and M. Behzadfar, 2009. Effect of orexin infusion into third ventricle on the GnRH and LH secretions in the prepubertal rat. J. Applied Sci., 10: 1936-1942.
CrossRef - Kim, B.H., A. Takemura, S.J. Kim and Y.D. Lee, 2003. Vitellogenin synthesis via androgens in primary cultures of tilapia hepatocytes. Gen. Comp. Endocrinol., 132: 248-255.
CrossRef - Kimaro, W.H., 2011. A histological and ultrastructural study of gland cells in the ovary of the sexually immature ostrich (Struthio camelus). Asian J. Biol. Sci., 4: 182-188.
CrossRefDirect Link - Kime, D.E., J.P. Nash and A.P. Scott, 1999. Vitellogenesis as a biomarker of reproductive disruption by xenobiotics. Aquaculture, 177: 345-352.
CrossRef - Kobayashi, T., P. Pakarinen, J. Torgersen, I. Huhtaniemi and O. Andersen, 2008. The gonadotropin receptors FSH-R and LH-R of Atlantic halibut (Hippoglossus hippoglossus)-2: Differential follicle expression and asynchronous oogenesis. Gen. Comp. Endocrinol., 156: 595-602.
CrossRef - Kwok, H.F., W.K. So, Y. Wang and W. Ge, 2005. Zebrafish gonadotropins and their receptors: I. cloning and characterization of zebrafish follicle-stimulating hormone and luteinizing hormone receptors-evidence for their distinct functions in follicle development. Biol. Reprod., 72: 1370-1381.
CrossRef - Kwon, H.C. and Y. Mugiya, 1994. Involvement of growth hormone and prolactin in the induction of vitellogenin synthesis in primary hepatocytes culture in the eel, Anguilla japonica. Gen. Comp. Endocrinol., 93: 51-60.
CrossRef - Lazier, C.B., S. Langley, N.B. Ramsey and J.M. Write, 1996. Androgen inhibition of vitellogenin gene expression in tilapia (Oreochromis niloticus). Gen. Comp. Endocrinol., 104: 321-329.
CrossRef - Le Menn, F., H. Rochefort and M. Garcia, 1980. Effect of androgen mediated by the estrogen receptor of fish liver: Vitellogenin accumulation. Steroids, 35: 315-328.
CrossRef - Levavi-Sivan, B., J. Bogerd, E.L. Mananos, A. Gomez and J.J. Lareyre, 2010. Perspectives on fish gonadotropins and their receptors. Gen. Comp. Endocr., 165: 412-437.
CrossRef - Levi, L., B. Levavi-Sivan and E. Lubzens, 2008. Expression of genes associated with retinoid metabolism in the trout ovarian follicle. Biol. Reprod., 79: 570-577.
CrossRef - Lubzens, E., G. Young, J. Bobe and J. Cerda, 2010. Oogenesis in teleosts: How fish eggs are formed. Gen. Comp. Endocrinol., 165: 367-389.
CrossRefDirect Link - Miura, C., T. Higashino and T. Miura, 2007. A progestin and an estrogen regulate early stages of ogenesis in fish. Biol. Reprod., 77: 822-828.
PubMed - Muntaziana, M.P.A., A.A. Rahim, S.A. Harmin and S.M.N. Amin, 2011. Estrogen-induced vitellogenin in tor tambroides (Bleeker, 1854): Purification, characterization and ELISA development. J. Fish. Aquatic Sci., 6: 700-714.
CrossRefDirect Link - Muntaziana, M.P.A., A. Abd Rahim and S.A. Harmin, 2011. Effect of broodfish sex ratio on seed production of red Tilapia in Suspended Hapa. J. Fish. Aquatic Sci., 6: 862-866.
CrossRefDirect Link - Nagahama, Y., 1994. Endocrine regulation of gametogenesis in fish. Int. J. Dev. Biol., 38: 217-229.
PubMedDirect Link - Nagahama, Y., 1997. 17α, 20β -Dihydroxy-4-pregnen-3-one, a maturation-inducing hormone in fish oocytes: Mechanisms of synthesis and action. Steroids, 62: 190-196.
CrossRef - Nagahama, Y. and M. Yamashita, 2008. Regulation of oocyte maturation in fish. Dev. Growth Differ., 50: S195-S219.
CrossRef - Ngamniyom, A. and B. Panyarachun, 2011. Expression levels of hormone receptor and Vitellogenin mRNAs in livers of thai medaka, Oryzias minutillus, inhabiting the suburbs of Bangkok, Thailand. J. Fish. Aquat. Sci., 6: 438-446.
CrossRefDirect Link - Ohkubo, N., T. Andoh, K. Mochida, S. Adachi, A. Hara and T. Matsubara, 2004. Deduced primary structure of two forms of vitellogenin in Japanese common goby (Acanthogobius flavimanus). Gen. Comp. Endocrinol., 137: 19-28.
CrossRef - Patino, R., P. Thomas and G. Yoshizaki, 2003. Ovarian follicle maturation and ovulation: An integrated perspective. Fish Phys. Biochem., 28: 305-308.
CrossRef - Peng, C., S. Humphries, R.E. Peter, J.E. Rivier, A.G. Blomqvist and D. Larhammar, 1993. Actions of goldfish neuropeptide Y on the secretion of growth hormone and gonadotropin-II in female goldfish. Gen. Comp. Endocrinol., 90: 306-317.
CrossRef - Peyon, P., S. Baloche and E. Burzawa-Gerard, 1997. Investigation into the possible role of androgens in the induction of hepatic vitellogenesis in the European eel: In vivo and in vitro studies. Fish Physiol. Biochem., 16: 107-118.
CrossRef - Ramadan, A.M. and M.M. El-Halfawy, 2007. Common forms of Atresia in the ovary of some red sea fishes during reproductive cycle. Pak. J. Biol. Sci., 10: 3120-3125.
CrossRefPubMedDirect Link - Santos, E.M., M. Rand-Weaver and C.R. Tyler, 2001. Follicle-stimulating hormone and its a and b subunits in rainbow trout (Oncorhynchus mykiss): Purification, characterization, development of specific radioimmunoassay and their seasonal plasma and pituitary concentrations in females. Biol. Reprod., 65: 288-294.
CrossRef - Sejian, V., T.V. Meenambigai, M. Chandirasegaran and S.M.K. Naqvi, 2010. Reproductive technology in farm animals: New facets and findings: A review. J. Biol. Sci., 10: 686-700.
CrossRefDirect Link - Selman, K. and R.A. Wallace, 1989. Cellular aspects of oocyte growth in teleost. Zool. Sci., 6: 211-231.
Direct Link - Srijunngam, J., N. Kitana, I.P. Callard and K. Wattanasirmkit, 2005. Ultrastructural changes in the ovarian follicular wall during oocyte growth in the Nile tilapia, Oreochromis niloticus linn. Nat. Hist. J. Chulalongkorn Univ., 5: 21-30.
Direct Link - Swanson, P., M. Bernard, M. Nozaki, K. Suzuki, H. Kawauchi and W.W. Dickhoff, 1989. Gonadotropins I and II in juvenile coho salmon. Fish Phys. Biochem., 7: 169-176.
CrossRef - Tavosanis, G. and C. Gonzalez, 2003. γ-Tubulin function during female germ-cell development and oogenesis in Drosophila. Cell. Biol., 100: 10263-10268.
CrossRef - Yaron, Z., 1995. Endocrine control of gametogenesis and spawning induction in the carp. Aquaculture, 129: 49-73.
CrossRef - Yaron, Z., G. Gur, P. Melamed, H. Rosenfeld, A. Elizur and B. Levavi-Sivan, 2003. Regulation of fish gonadotropins. Int. Rev. Cytol., 225: 131-185.
CrossRefDirect Link - Wallace, R.A. and K. Selman, 1990. Ultrastructural aspects of oogenesis and oocyte growth in fish and amphibians. J. Electron. Micr. Tech., 16: 175-201.
CrossRef - Weber, G.M., J.F. Powell, M. Park, W.H. Fischer and A.G. Craig et al., 1997. Evidence that gonadotropin-releasing hormone (GnRH) functions as a prolactin-releasing factor in a teleost fish (Oreochromis mossambicus) and primary structures for three native GnRH molecules. J. Endocrinol., 155: 121-132.
PubMedDirect Link - Wong, T. and Y. Zohar, 2004. Novel expression of gonadotropin subunit genes in oocytes of the gilthead seabream (Sparus aurata). Endocrinology, 145: 5210-5220.
CrossRef - Zalina, I., C.R. Saad, A.A. Rahim, A. Christianus and S.A. Harmin, 2011. Breeding performance and the effect of stocking density on the growth and survival of climbing perch, Anabas testudineus. J. Fish. Aquatic Sci., 6: 834-839.
CrossRefDirect Link