
X.R. Wang
College of Animal Science and Technology, Gansu Agricultural University, Lanzhou 730070, China
Y. Liu
College of Animal Veterinary, Gansu Agricultural University, Lanzhou 730070, China
L.P. Zhang
College of Animal Science and Technology, Gansu Agricultural University, Lanzhou 730070, China
X.J. Wang
College of Animal Science and Technology, Gansu Agricultural University, Lanzhou 730070, China
J.P. Wu
College of Animal Science and Technology, Gansu Agricultural University, Lanzhou 730070, China
Asian Journal of Animal and Veterinary Advances
Year: 2012 | Volume: 7 | Issue: 9 | Page No.: 884-890







ABSTRACT
The epidural retia mirabile is an arterial network formed by many of anastomotic branches from adjacent arteries for supplying blood to the brain in artiodactyla and plays a vital role in regulating cerebral blood flow. The purpose of this work was to compare anatomical differences of the epidural retia mirabile between the yak and cattle and to analyse their blood supply characteristics and provide a morphological basis for further research on the plateau adaptability of the yak. The vascular casts of the epidural retia mirabile were yielded on head specimens of 12 yaks (obtained from Qinghai China) and 10 cattle (obtained in Gansu China) by the corrosion casting technique. Their morphological features and arterial diameters were compared and analyzed using the methods of comparative anatomy. The general arrangements of the epidural retia mirabile in the yak were similar to that seen in cattle and mainly composed of the anterior portion, posterior portion, basi-occipital arterial plexus and anterior V-shaped extension. Middle width of the anterior epidural retia mirabile was more extensive and the posterior joint section was more developed and there were more abundant anterior anastomotic branches and communicating rami in the epidural retia mirabile of the yak. Moreover, the distribution ratios of blood flow from the maxillary artery entering the anterior epidural retia mirabile were 33.12% and 30.42% in the yak and in cattle, respectively. Our results indicated that the yak has more developed epidural retia mirabile. Its anatomical peculiarity perhaps is an adaptive characteristic of supplying blood system to the brain and which would help to buffer and regulate cerebral blood flow better so as to meet the cerebral oxygen demand of the yak in plateau areas.
PDF Abstract XML References Citation
Received: January 06, 2012;
Accepted: March 06, 2012;
Published: May 10, 2012
How to cite this article
X.R. Wang, Y. Liu, L.P. Zhang, X.J. Wang and J.P. Wu, 2012. Comparative Anatomical Study of the Epidural Retia Mirabile in the Yak and Cattle. Asian Journal of Animal and Veterinary Advances, 7: 884-890.
DOI: 10.3923/ajava.2012.884.890
URL: https://scialert.net/abstract/?doi=ajava.2012.884.890
DOI: 10.3923/ajava.2012.884.890
URL: https://scialert.net/abstract/?doi=ajava.2012.884.890
INTRODUCTION
The Epidural Retia Mirabile (ERM) has been also called the rete mirabile epidurale, arterial retia mirabile, carotid retia or rete which was found commonly within the cranium of artiodactyla (Uehara et al., 1978; Ocal and Aslan, 1994; Khamas and Ghoshal, 1985; Ocal et al., 1998). So far, abundant studies have been performed about the structures and functions of the ERM in many animals including bovine. Previous reports showed that the carotid rete arises from the external carotid artery as hundreds of small arterioles in artiodactyla (Mitchell and Lust, 2008). The carotid retia carries arterial blood supplying the brain, replacing the absent internal carotid artery (Fukuta et al., 2010) and it acts as a flow-facilitating system in a situation in which smooth muscle of brain vessels contracts in response to stretch (Dieguez et al., 1988) and regulate blood temperature (Jessen, 2001). Hence, animals that have a rete have an additional weapon in their thermoregulatory arsenal denied to non-rete animals (Mitchell and Lust, 2008). Presently a study have referred to morphological description of the ERM in the yak (Ding et al., 2007), but no reports have been published on comparison of anatomical features of the ERM between the plateau yak and low-altitude cattle.
The yak (Bos grunniens) is regarded as one of the world’s most remarkable domestic animals as it thrives in conditions of extreme harshness and deprivation while providing a livelihood for people (Gerald et al., 2003). At present, more and more reports have focused on the plateau adaptive characteristics of the yak. In this study, we observed the anatomical peculiarity of the ERM in the yak and cattle by the vascular corrosion casting technique and examined the arterial diameters and anastomosis of the vessels meshwork by comparative anatomy methods. The present study has the objectives to analyse the characteristics of the yak plateau adaptability on blood supply to the ERM and provide some morphological basis on cerebral arteries for further research in plateau yak.
MATERIALS AND METHODS
Experimental materials: Twelve heads from healthy and adult yaks that living in Haibei (altitude is 3,800 m), Qinghai, were obtained immediately after slaughter in Lejiawan slaughterhouse of Xining, China and ten heads from cattle were obtained in Guanghe (altitude is 1,800 m), Gansu, China. Yaks were approximately 8-10 years old and body weight seemed to be 220-250 kg, cattle were 3-4 years old and body weight seemed to be 450-500 kg. The animals were killed by exsanguination for food and specimens were demanded fresh, intact and pollution-free.
Experimental procedures: The vascular corrosion casting procedures were carried out due to the reference (He, 2007) and made some improvement in present study. The ABS resin (Toray Industries. Inc., Japan) solution was injected into the cerebral arteries via bilateral common carotid arteries, then the specimens were kept for 48 hours for polymerization at room temperature. At last they were soaked in concentrated hydrochloric acid (Hangzhou Linglong Chemical. Co., Ltd., China) for 7-10 days for maceration. After having finished corrosion, remnants of the brain tissue were carefully flushed with pipe water, until vascular casts were emerged totally.
Data obtaining and analysis: Specimens’ figures were taken with digital camera and diameters and sizes were measured by vernier caliper. The internal diameters of arteries were measured at the origin of the vessels. In addition, specimens’ morphological features were observed with naked eyes, their differences were described and documented.
RESULTS
General architectural features of the ERM in the yak: General architectural features of the ERM in the yak were similar to those seen in cattle. The ERM was consisted of the anterior portion, posterior portion, basi-occipital arterial plexus and anterior V-shaped extension. The anterior ERM jointed with 5-7 anterior anastomotic branches and one posterior anastomotic branch from the maxillary and external ophthalmic arteries. Owing to the ERM completely surrounded the pituitary gland, a distinct hole was seen in the middle of casts. The left and right sides of the anterior ERM showed two bilateral lobes and adjacent small vessels in it were anastomotic each other. The posterior ERM jointed with the basi-occipital arterial plexus formed by the occipital, vertebral and condylar arteries. The anterior and posterior ERM were jointed by several communicating rami.
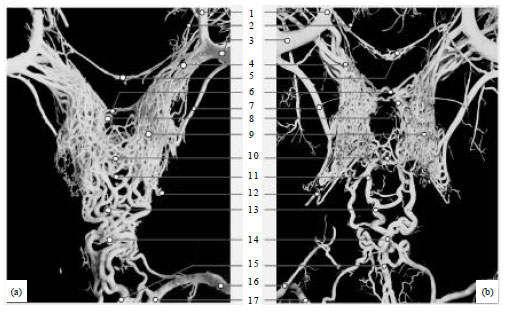 | |
| Fig. 1(a-b): | Corrosion casts from the ERM in yak and cattle, (a) Yak (b) Cattle, (1) External ophthalmic artery, (2) Internal ophthalmic artery, (3) Maxillary artery, (4) Anterior anastomotic branches, (5) V-shaped extension, (6) Anterior joint section, (7) Posterior anastomotic branch, (8) Cerebral carotid artery, (9) Right lob of the ERM, (10) Posterior joint section, (11) Communicating rami, (12) Internal carotid artery, (13) Posterior ERM, (14) Basi-occipital arterial plexus, (15) Vertebral artery, (16) Occipital artery and (17) Condylar artery |
The anterior V-shaped extension of the ERM lied at the bottom of optic chiasm and its two sides jointed with the maxillary and internal ophthalmic arteries (Fig. 1).
The anterior ERM of the yak: The partial features of the anterior ERM between the yak and cattle showed significant differences. The anterior ERM in the yak looked like U-shaped and its middle width was 33.98 mm meanwhile the anterior ERM in cattle looked like H-shaped and its middle width was 30.23 mm. The number of the anterior anastomotic branches from the maxillary and external ophthalmic arteries was 6.73 and 5.22 on average in the yak and cattle, respectively. In present study, however, we found that the diameter of the posterior anastomotic branch of the anterior ERM in the yak was thinner (1.49 mm) than those in cattle (1.96 mm) and diameters of communicating rami of the posterior ERM in the two species were similar.
The joint sections communicating with two lobes of the anterior ERM were composed of some small vessels laying at the anterior or posterior portion of the pituitary gland. The anterior joint section in the yak was formed by 2-3 fine arteries while the posterior joint section was formed by two vessels network layers. The anterior joint section in cattle was formed by several middle arteries while the posterior joint section was formed mainly by a larger artery whose diameter was beyond 1.0 mm (Fig. 1).
In addition, we also found that the internal carotid artery whose diameter was about 0.50 mm at the retral end of the anterior ERM (Fig. 2a). It arose from the middle section of the occipital artery and entered into the cranial cavity through the jugular foramen, formed a loop bending ventrally, at last entered the anterior ERM. The course of the internal carotid artery was similar in the two animal species.
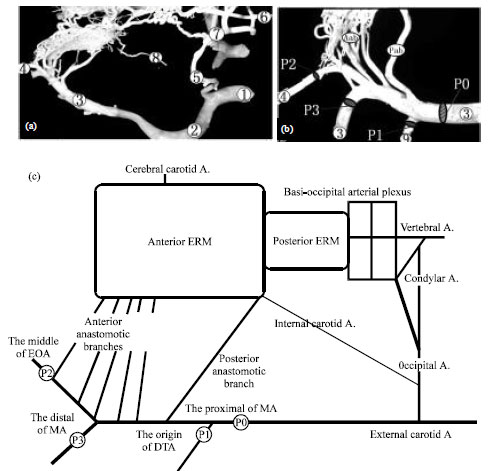 | |
| Fig. 2(a-c): | Main arteries of supplying blood to the ERM and the diagram, (a) Main arteries of supplying blood to the ERM, (b) Main branches from the maxillary artery and (c) Supplying blood diagram, (1) Common carotid artery, (2) External carotid artery, (3) Maxillary artery, (4) External ophthalmic artery, (5) Occipital artery, (6) Vertebral artery, (7) Condylar artery, (8) Internal carotid artery and (9) Deep temporal artery, Pab: Posterior anastomotic branch, Aab: Anterior anastomotic branches, P0: Proximal of MA, P1: Origin of DTA, P2: Middle of EOA, P3: Distal of MA, MA: Maxillary artery, DTA: Deep temporal artery and EOA: External ophthalmic artery |
The posterior ERM of the yak: Anatomical features of the posterior ERM between the yak and cattle also showed definite differences. The rostral section of the posterior ERM of the yak was wider and the retral section was narrower than that of cattle, so it looked like inverted ladder-shaped while the rostral and retral sections of cattle were homogeneous (Fig. 1). Furthermore, the number of communicating rami in the posterior ERM was 5.43 and 3.21 in the yak and cattle, respectively.
The basi-occipital arterial plexus and the anterior V-shaped extension: The basi-occipital arterial plexus located at distal portion of the posterior ERM and it was a vessels network formed by the occipital, vertebral and condylar arteries. Its vascular diameters were thick and its communicating rami jointed with all adjacent arteries. The anterior V-shaped extension was also found in the two animal species and their constituting vessels were too thin to see intact structures in casting specimens (Fig. 1). In present study, we found that the two structural elements in the yak and cattle had no distinct differences.
| Table 1: | Distribution ratio of blood flow from the maxillary artery entering the ERM |
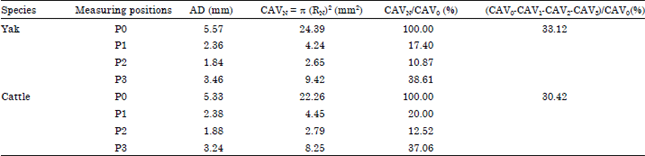 | |
| AD: Average diameter, R = D/2, N: P0, P1, P2 or P3, CAVN: Cross area of vessels, P0: Proximal of MA, P1: Origin of DTA, P2: Middle of EOA, P3: Distal of MA, MA: Maxillary artery, DTA: Deep temporal artery, EOA: External ophthalmic artery | |
Characteristics of blood supply to the ERM in the yak: The anterior anastomotic branches arose from the maxillary and external ophthalmic arteries and the posterior anastomotic branch arose from the maxillary artery carried partial blood flow from the maxillary artery entering the ERM (Fig. 2b). The blood flow of the ERM was collected by the cerebral carotid artery and carried to the cerebral arterial circle and then blood flow was carried to the brain tissues. Casts showed the left and right cerebral carotid arteries were the main exits of the blood flow of the ERM toward the brain. Moreover, the basi-occipital arterial plexus was also a main channel of supplying blood to the posterior ERM and it received blood supply from the vertebral, occipital and condylar arteries (Fig. 2b). Present study displayed that the main arteries supplying blood to the ERM in the yak and cattle were the maxillary, occipital, vertebral, external ophthalmic, condylar and internal carotid arteries, meanwhile these arteries were also the indirect source of supplying blood to the brain tissues (Fig. 2).
In present study, we measured the average diameters of four arteries at certain positions including the proximal of the maxillary artery (P0), the origin of the deep temporal artery (P1), the middle of the external ophthalmic artery (P2) and the distal of the maxillary artery (P3) and calculated the cross area of each artery. The results showed, in the course of supplying blood to the ERM via the maxillary artery, except for supplying blood to external region of the ERM, the gross cross area of vessels carrying the blood of entering the ERM was 33.12% of the cross area of the proximal of the maxillary artery in the yak and 30.42% in cattle (Fig. 2, Table 1).
DISCUSSION
General morphological features of the ERM in the yak: The general arrangements of the ERM in the yak were similar to that seen in cattle, but it implied several anatomical peculiarities in some aspects. The macroscopic shape of the anterior ERM in the yak showed like letter “U” but like letter “H” in cattle. The anterior ERM of the yak was more extensive and the anterior anastomotic branches from the maxillary and external ophthalmic arteries were more abundant than those of cattle. In present study, we generally agreed with the correlative reports from references (Uehara et al., 1978; Ding et al., 2007). In the yak, however, we still found that there were more abundant communicating rami in the posterior ERM and the posterior joint section and posterior ERM were also more developed than that of cattle. Consequently, our results illustrated that the anatomical architecture of the ERM in the yak were evidently different to those seen in cattle, it perhaps be an evolving characteristic of supplying blood system to the brain in the yak during adapting the plateau environments.
In addition, we observed the anterior V-shaped extension presenting at the rostral portion of the ERM and their structures were similar between the two species. Though, the diameters of its main vessels were a little different, we considered it was caused by different individual development. Some studies showed that the anterior V-shaped extension of the ERM was only present in the calf (Uehara et al., 1978), but buffalo not (HRG, 1984). From our results, it was deduced that the anterior V-shaped extension of the ERM might be the common feature in most bovine animal.
Furthermore, some reports demonstrated that the posterior ERM of the ox communicated directly with the vertebral artery through the form of the basi-occipital arterial plexus (Baldwin, 1964). We found that the basi-occipital arterial plexus was present in the ERM of the two species and their rostral and retral portions were jointed with the posterior ERM and the vertebral artery, respectively, meanwhile the left and right rami were jointed with the occipital and condylar arteries.
Distribution ratio of blood flow from the maxillary artery to the ERM in the yak: Changes of vascular calibers become the main factor determining blood flow if the differential pressure of artery and vein would be similar. Some studies pointed out that the ratio of the blood supply to the ERM can be presumed from the diameter of each vessel cast of the afferent arteries (Uehara et al., 1978; Ocal et al., 1998). In the course of the maxillary artery carrying the blood flow, the partial blood flow was distributed to the infraorbital, deep temporal and external ophthalmic arteries, but remaining blood flow was mainly distributed to the ERM via the anastomotic branches. Our results illustrated that the distribution ratios of blood flow from the maxillary artery entering the ERM in the yak was significantly higher than those in cattle, resulting the blood flow of the ERM receiving from the maxillary artery in the yak would be more sufficient than those in cattle. We considered it was caused mainly by the anatomical peculiarity of the ERM of the yak having been formed in the high-altitude environments.
Arterial supply to the ERM in the yak: Previous studies showed that the major contribution of blood reached the carotid arterial rete through the arterial anastomosis and ramus anastomosis, the branches of the internal maxillary artery in the buffalo (Bamel et al., 1975). In the ox, the main source of blood supply to the ERM was the external carotid artery via branches of the internal maxillary (Baldwin, 1964) and the ERM received additional branches at its caudal end from the vertebral and occipital arteries (Gillian, 1974). In present study, we considered that the basi-occipital arterial plexus in the yak and cattle was a vascular hinge of connecting several supplying blood vessels to the ERM. Some arteries including the vertebral, occipital, condylar arteries and communicating rami of the posterior ERM were jointed by it each other, so it should be regarded as the main structural basis of supplying blood to the posterior ERM. From above all, we concluded that the ERM of the yak received the blood supply mainly from the maxillary, occipital, vertebral, condylar, external ophthalmic and internal carotid arteries and the maxillary artery was the most important supplying blood source among these. Except for the anastomotic branches from the maxillary artery itself, still there were the anastomotic branches from the external ophthalmic artery, to form common blood supply channel to the anterior ERM becoming main source of arterial supply to the ERM.
CONCLUSION
Our studies demonstrated that the general arrangements of the ERM and their main sources of supplying blood in the yak were similar to that seen in cattle. There were the V-shaped extension and basi-occipital arterial plexus in the ERM of the yak which could be regarded as the common structural feature in most bovine animal. The arterial supply to the ERM in the yak mainly came from the maxillary, occipital, vertebral, condylar, external ophthalmic and internal carotid arteries and was similar to those in cattle. Remarkably, relative to cattle living in the low-altitude areas, the ERM in the yak was more developed and there were more abundant anterior anastomotic branches and communicating rami in it. The distribution ratios of blood flow from the maxillary artery entering the ERM in the yak was significantly higher than that in cattle. The results of the present study indicated that the anatomical peculiarity of the ERM in the yak perhaps be a adaptive characteristic of supplying blood system to the brain which would help to buffer and regulate cerebral blood flow better so as to meet the cerebral oxygen demand of the yak in plateau areas.
ACKNOWLEDGMENTS
This study was partially funded by the National Natural Science Foundation (30960164). In addition, we thank Binyun Ma from the Lanzhou University and Xin Cai from the Southwest University of Science and Technology for all their support.
REFERENCES
- Baldwin, B.A., 1964. The anatomy of the arterial supply to the cranial regions of the sheep and ox. Am. J. Anat., 115: 101-117.
CrossRefDirect Link - Dieguez, G., A.L. Garcia-Villalon, B. Gomez and S. Lluch, 1988. Hemodynamic significance of the carotid rete during changes in arterial blood pressure. Am. J. Physiol., 5: 770-775.
PubMed - Ding, Y., B. Shao and J. Wang, 2007. The arterial supply to the brain of the yak (Bos grunniensi). Ann. Anat., 189: 31-38.
PubMed - Jessen, C., 2001. Selective brain cooling in mammals and birds. Jpn. J. Physiol., 51: 291-301.
PubMed - Fukuta, K., H. Kudo, M. Sasaki, J. Kimua, Dahlan bin Ismil and H. Endo, 2010. Absence of carotid rete mirabile in small tropical ruminants: Implications for the evolution of the arterial system in artiodactyls. J. Anatomy, 210: 112-116.
Direct Link - Khamas, W.A. and N.G. Ghoshal, 1985. Gross and scanning electron microscopy of the carotid rete-cavernous sinus complex of the sheep (Ovis aries). Anat. Anz., 159: 173-179.
PubMed - Mitchell, G. and A. Lust, 2008. The carotid rete and artiodactyl success. Biol. Lett., 4: 415-418.
CrossRef - Ocal, M.K., H. Erden, I. Ogut and M.E. Kara, 1998. A quantitative study on the retial arteries in one-humped camels. Ann. Anat., 180: 369-371.
CrossRefDirect Link - Ocal, M.K. and K. Aslan, 1994. A putative study on the retinal arteries in the bovine fetus. Ann. Anat., 176: 151-153.
PubMed - Bamel, S.S., L.D. Dhingra and D.N. Sharma, 1975. Anatomical studies on the arteries of the brain of buffalo (Bubalus bubalis). I. The rete mirabile cerebri. Anat. Anz., 137: 440-446.
PubMed