
Hong-Quan Chen
School of Animal Science and Technology, Anhui Agricultural University, Hefei 230036, The People`s Republic of China
Jie Qin
School of Animal Science and Technology, Anhui Agricultural University, Hefei 230036, The People`s Republic of China
Yin-Jian Zhu
School of Animal Science and Technology, Anhui Agricultural University, Hefei 230036, The People`s Republic of China
Zhong-Ting Pan
School of Animal Science and Technology, Anhui Agricultural University, Hefei 230036, The People`s Republic of China
Ya-Nan Xie
School of Animal Science and Technology, Anhui Agricultural University, Hefei 230036, The People`s Republic of China
Ming-Hui Jiao
School of Animal Science and Technology, Anhui Agricultural University, Hefei 230036, The People`s Republic of China
Gong-Wei Chen
School of Animal Science and Technology, Anhui Agricultural University, Hefei 230036, The People`s Republic of China
Hua Chen
School of Animal Science and Technology, Anhui Agricultural University, Hefei 230036, The People`s Republic of China
Ming-Xing Chu
Key Laboratory of Farm Animal Genetic Resources and Utilization of Ministry of Agriculture, Institute of Animal Science, Chinese Academy of Agricultural Sciences, Beijing 100193, The People`s Republic of China
Asian Journal of Animal and Veterinary Advances
Year: 2012 | Volume: 7 | Issue: 9 | Page No.: 802-811







ABSTRACT
THRSP gene has been regarded as a candidate gene for lipogenesis in domestic animals. To understand the association of THRSP gene with habitats and fatty deposits in goat breeds, the polymorphisms of goat THRSP gene, the fat content of goat carcass and their association with the ecological factors were investigated. The Chinese indigenous goat breeds selected were Guishan, Yunling, Chuandong, Banjiao, Xiangdong, Matou and Anhui white goat. The polymorphisms in the 5’ regulatory region and exon 1 of THRSP gene were analyzed using PCR-RFLP. The longissimus Muscle Fat Grade (MFG) and Carcass Fat Grade (CFG) of all goat breeds were estimated. The observed data of ecological factors were provided by the meteorological observatory in the goat habitats which included Average Daily Rainfall (ADR), Elevation (E), average Daily Sunshine Duration (DSD), Latitude (LAT) and longitude. E/LAT, ADRxDSD and SCxEC were regarded as Space Coefficient (SC), Energy Coefficient (EC) and Ecological Index (EI), respectively. The results showed that three mutational sites, C-233T, G113A and A138G, were detected. Nine genotypes in all goat breeds were only found, of which the frequencies of TCAAGG, TTGAAG and TTAAGG genotype were lower than 0.05. The EC significantly related to CAG haplotype frequency (0.8859, p<0.01) and the associations of EI with TGA haplotype frequency was significant (-0.8377, p<0.025). In the four haplotype frequencies, the relationship between CAG haplotype frequency and MFG was significant (-0.8633, p<0.025). The results suggest that the polymorphism of goat THRSP gene may be associated with the ecological factors and affect the goat lipogenesis ability.
PDF Abstract XML References Citation
Received: February 26, 2012;
Accepted: April 09, 2012;
Published: May 10, 2012
How to cite this article
Hong-Quan Chen, Jie Qin, Yin-Jian Zhu, Zhong-Ting Pan, Ya-Nan Xie, Ming-Hui Jiao, Gong-Wei Chen, Hua Chen and Ming-Xing Chu, 2012. The Polymorphisms of Goat THRSP Gene Associated with Ecological Factors in Chinese Indigenous Goat Breeds with Different Lipogenesis Ability. Asian Journal of Animal and Veterinary Advances, 7: 802-811.
DOI: 10.3923/ajava.2012.802.811
URL: https://scialert.net/abstract/?doi=ajava.2012.802.811
DOI: 10.3923/ajava.2012.802.811
URL: https://scialert.net/abstract/?doi=ajava.2012.802.811
INTRODUCTION
The domestic goats originated in the wild goat ancestor inhabiting in mountainous area and being constantly domesticated (Zeder and Hesse, 2000). There are more than 150 goat breeds in the world which can be divided into the milk, fiber, skins, meat and dual-purpose categories depending on their use (Coffey et al., 2004; Pal et al., 2011; Fang et al., 2010). China has vast land with complex terrain and climate from north to south across the frigid, temperate and torrid zones and the Chinese indigenous goat breeds nearly 50, according to Tu et al., (1989). With the need of the consumers to mutton in China, the proportion of goat meat rosed up from 40% in 2000 to 45% in 2010. The consumption of goat meat is changing from the quantity to the quality. However, the goat breeds from different habitats differ significantly in the meat quality, business grades, taste, flavor and juiciness which result in the problems in the prices of goat meat and the economic profits of the producers (Sebsibe, 2008). Seven Chinese indigenous goat populations were investigated with different meat quality in this study: these goat breeds distributed among the Central China, East China and Southwest region (Zhang, 1994), of which some goat breeds have a strong adaptability to the habitats with high mountains, forest and abundant rainfall while others prefer the hilly land on the lower slopes of mountains and the habitats with cool and relatively dry climate (Liu and Feng, 1993).
The key factors affecting the meat quality are environmental factors and genetic factors (Hardini and Djunaidi, 2010; Askari-Hemmat, 2006; Orhan and Kaygisiz, 2009). In the investigation of the genetic mechanism of animal fat metabolism, the genes of some enzymes (Wakil and Abu-Elheiga, 2009) and hormones (Etherton, 2000; Paracchini et al., 2005) in fat metabolism and physiological regulation were analyzed and some molecular markers associated with meat fat content were found which provided the scientific basis in the improvement of important economic characters of goat breeds (Esmaeelkhanian et al., 2007). Recently, the role of thyroid hormone responsive spot 14 (THRSP) in lipogenesis caused the attention of researchers (Zhou et al., 2011). THRSP gene is localized to human chromosome 11 q13.5 region implicated in obesity. The mutation of THRSP gene can cause the obstacles of fat metabolism and disorders of lipogenesis in the body which affect meat quality of animals. At present, the research of THRSP gene was mainly on people, rats and mice and changed to chicken, duck, pig and cattle (Chou et al., 2007; Chen, 2010) but the reports were very few in goat. In ecological environment, some reports showed that the diversity of animal breeds was influenced by the space factors such as elevation and latitude and the energy factors, for example, sunshine duration, average rainfall and temperature (Whittaker, 1977; Rohde, 1992). The ecological factors play an important role in the cultivated process of animal breed via affecting the metabolic and physical regulation of the body. However, the association of ecological factors with goat THRSP gene yet remains unclear.
Based on goat breeds with special habitats, this paper analyzed the polymorphisms of goat THRSP gene and their distribution patterns with ecological factors and investigated the association of the THRSP gene polymorphism with goat fatty deposits and ecological factors.
MATERIALS AND METHODS
Animal sample collection: Seven native goat breeds selected derived from their habitats in China, respectively which are Guishan goat (GS) from Wanjiahuan goat breeding farm of Shilin Yi Nationality Autonomous County (Yunnan, n = 47), Yunling black goat (YL) from Yunnan goat breeding farm of Xundian County (Yunnan, n = 47), Chuandong white goat (CD) from Yikouxian goat breeding farm of Yunyang County (Chongqing, n = 48), Banjiao goat (BJ) from Banjiao goat breeding farm of Wuxi County (Chongqing, n = 44), Xiangdong black goat (XD) from Xiangdong black goat sheep stud of Changsha (Hunan, n = 51), Matou goat (MT) from Matou goat breed conservation field of Shimen County (Hunan, n = 93) and Anhui white goat (AW) from Boda Feidong sheep stud (Anhui, n = 122). Genomic DNA samples from the ear’s tissues of all goats were extracted using DNA extraction reagent box (Beijing Tiangen Biotechnology, China) and stored at 4°C.
Data collection of ecological factors: The observed data of ecological factors were provided by the meteorological observatory in the habitats of seven native goat breeds in 2001 to 2011 which include Average Daily Rainfall (ADR), Elevation (E), average Daily Sunshine Duration (DSD), Latitude (LAT) and Longitude (LON). LAT and E are the space factors in animal diversity. ADR and DSD are the energy factors. Given: E/LAT was regarded as Space Coefficient (SC), ADRxDSD was regarded as Energy Coefficient (EC) and SCxEC was regarded as Ecological Index (EI).
Assessment of goat meat grade: The Muscle Fat Grade (MFG) of goat breed was estimated according to the fat content of goat longissimus muscle and less than 1.0% in the muscle as level 1, then each additional 0.5% to improve a grade, respectively. The Carcass Fat Grade (CFG) was assessed based on the fat content of goat carcass and less than 10.0% in the carcass as level 1, then each additional 1.0% to improve a grade, respectively (Jin et al., 2012).
Detection of polymorphisms
Primers: Based on the sequence of cattle THRSP gene (GenBank AY 656814), two pairs of primers, S1 and S2, were designed using the Primer Premier 5.0 program which were employed for amplifying the fragments containing the 5’ partial regulatory region and the exon 1 of goat THRSP gene, respectively. Then two pairs of primers, TP1 and TP2, were designed for analyzing the Restriction Fragment Length Polymorphism (RFLP) of PCR products of S1 and S2, respectively. All primers were synthesized by Shanghai Bioengineering Inc. (Shanghai, China). The sequences of four pairs of primers were as follows:
| • | S 1: | Sense 5’- GGGTCAATGGTGAAGACGAAGCT-3’, Anti-sense 5’- GCCAAGTACCGGTCCATGACG-3 |
| • | S 2: | Sense 5’- ATTGGCCTAAAAGAGGGCTATGT -3’, Anti-sense 5’- CTCAATACTCAAAAGCGAGGGTG -3 |
| • | TP 1: | Sense 5’- CTCTAAATGGCTCTGCAAGCTGATG-3’, Anti-sense 5’- TAGCCCTCTTTTAGGCCAATCCTAG-3 |
| • | TP 2: | Sense 5’- CTCTAAATGGCTCTGCAAGCTGATG-3’, Anti-sense 5’- GCCCCAATACCCTCTCTTCCT-3 |
PCR-RFLP: PCR was carried out using 25 μL, PCR amplification kits: 1.0 μL (10 μM) of each primer, 2.5 μL of 10xbuffer with Mg2+, 2.0 μL dNTP (2.5 mM), 0.5 μL (5 units μL-1) Taq DNA polymerase (Beijing TransGen Biotech Co., Ltd. China), 1.0 μL of genomic DNA (50 ng μL-1) and 17.0 μL double-distilled H2O. Amplification conditions were: initial denaturation of 5 min at 94°C; 35 cycles of 94°C for 30 sec., 30 sec. at 58°C for TP1 (at 57°C for TP2) and 72°C for 50 sec., followed by 72°C extension for 10 min. The PCR products were detected using 1.5% agarose gel electrophoresis. The PCR products of mixed samples of goat DNA were submitted to the Shanghai Invitrogen Biological Company for sequencing. According to sequencing results, three restriction enzymes, NIa III, Pst I and Kpn I (MBI), were selected for RFLP analysis.
Statistical analysis: A pair of allele frequencies were calculated by p = D+0.5 H and q = 1-p in which D is the homozygous frequency and H is the heterozygous frequency (Chen et al., 2011a; Zhou et al., 2011). The linkage disequilibrium between two loci, the coefficients of linkage disequilibrium (D and r2) and the haplotype frequencies were analyzed via http://www.oege.org/software/cubex/. The sequence alignment was analyzed via http://www.ncbi.nlm.nih.gov/. The promoter and transcription factors binding loci were predicted using Signal scan (http://www-bimas.cit.nih.gov/molbio/signal/). The correlation coefficients between the gene frequencies and the MFG, CFG and ecological factors were calculated and evaluated for p = 0.05 significant relationship using student’s t-test.
RESULTS
Sequence analysis: The fragment lengths of the products with S1 and S2 primers were 1105 bp and 715 bp, respectively and the sequences were published in GenBank (No. JN684754). Comparing the nucleotide sequences in the 5' regulatory region of goat THRSP gene with cattle and human, the homologies were 97% and 80%, respectively and 88% and 95% in the exon 1 with pig and cattle. The potential promoter region predicted located in -81~-32 bp upstream of the Start Codon (USC) of the THRSP gene in Fig. 1.
 | |
| Fig. 1: | Sequence characters of promoter region and exon 1 of goat THRSP gene. The primers with same names in the arrow on the right (upstream primer) or left side (downstream primer) are a pair of primers and the region between upstream and down stream primers is the DNA sequence. The sequence with capital letters is the promoter predicted. The AGGCAGTCAC sequence was binding motif for NF-κB transcription factors in which the GTCAC was R-box for bHLH-H-lcucine (bHLH-Zip) transcription factors binding site. The CDS length of THRSP gene is 453 bp, The C/T (C-233T), G/A (G113A) and A/G (A138G) are the polymorphic loci. C to T at C-233T results in the binding site disappeared and G° A at G113A leads to the arginine replaced by glutamine at 38th of THRSP protein but there was no amino acid change at A138G |
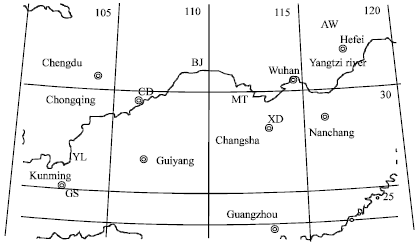 | |
| Fig. 2: | The distribution of seven native goat breeds in China. |
| Table 1: | The frequencies of genotype and haplotype and the coefficients of linkage disequilibrium of C-233T, G113A and A138G sites and their distribution in seven goat breeds |
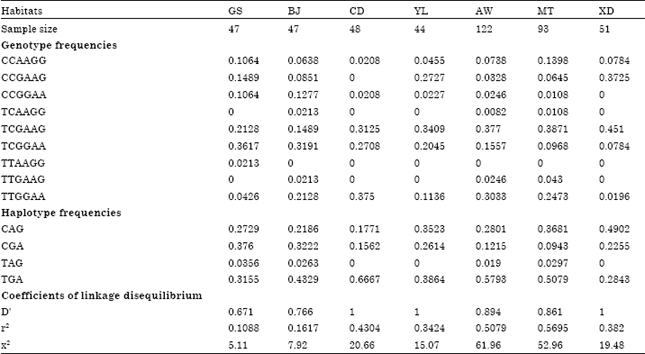 | |
| GS: Guishan goat, YL: Yunling black goat, CD: Chuandong white goat, BJ: Banjiao goat, XD: Xiangdong black goat, MT: Matou goat and AW: Anhui white goat | |
In the 5' regulatory region, there are nine E-boxes for basic Helix-loop-helix (bHLH) transcription factor binding site, one CCAAT box and two ATTGG boxes. AGGCAGTCAC sequence located in -242 bp~-233 bp USC was NF-κB transcription factors binding site and contained one R-box (GTCAC) for bHLH-leucine zipper (bHLH-Zip) transcription factor binding site that regulated lipid synthesis in Eukaryotic cells. In the seven goat breeds, a mutation site was detected in the 5’ regulatory region located at -233 bp USC (C-233T).
| Table 2: | The correlation coefficients between the ecological factors and the haplotype frequencies |
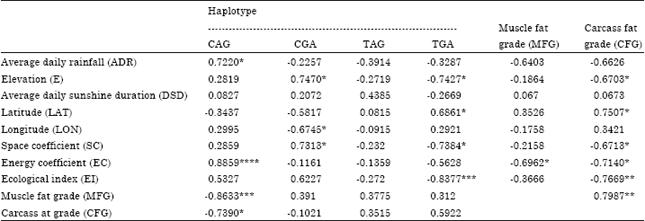 | |
| SC: LOG/LAT, EC: ADRxDSD, EI: SCxEC, *0.05<p<0.1, **0.025<p<0.05, ***0.01<p<0.025, ****p<0.01 | |
According to the sequence of exon 1, goat THRSP protein was composed of 150 amino acids and contained three protein kinase C phosphorylation sites and one casein kinase II phosphorylation site. Two mutations were founded in coding region. One was located at 113 bp of the CDS of THRSP gene (G113A) and another was located at 138 bp (A138 G). The former, nucleotide changed from G to A, led to amino acid substitute at 38th of THRSP protein from arginine to glutamine but there was no amino acid change in the later. Theory isoelectric point and unstability coefficient of THRSP protein with arginine were 5.88 and 32.45, respectively while those with glutamine were 5.72 and 34.62.
Polymorphism distribution: After the PCR products digested with NIa III, Pst I and Kpn I for C-233T, G113A and A138G sites, respectively, the linkage genotype frequencies of all goat breeds were calculated in Table 1. Nine genotypes were only found in seven goat breeds, of which the frequencies of TCAAGG, TTGAAG and TTAAGG genotype were lower than 0.05. The genotypic frequency of TCGAAG in XD was higher, 0.4510 and gradually reduced to 0.1489 from MT, AW, YL, CD, GS to BJ. However, the frequency distribution of TCGGAA genotype in seven goat breeds was contrary to that of TCGAAG genotype. The genotypic frequencies, moreover, of TTGGAA in CD and AW were higher than 0.30. The TAG haplotype frequencies were lower than 0.036 in all goat breeds while the TGA haplotype frequencies in CD, AW and MT were 0.6667, 0.5793 and 0.5079, respectively. From the coefficients of linkage disequilibrium among C-233T, G113A and A138G sites, CD, YL and XD were in complete linkage disequilibrium and AW, MT, BJ and GS in the some degree linkage disequilibrium.
Association of haplotype frequencies with ecological factors: Seven goat breeds were distributed between 24 to 36°0' north latitude and 103 to 117°50' longitude east (Fig. 2). The correlation coefficients between the ecological factors and the haplotype frequencies were listed in Table 2. The CAG haplotype frequency rose with the increase of ADR and EC and the relationship (0.8859) between EC and CAG haplotype frequency achieved at the significant level of 0.01. In the negative effects of E, SC and EI on TGA haplotype frequency, the association of EI with TGA haplotype frequency was significant (-0.8377, p<0.025).
From Table 2, the association of the ecological factors with CFG was stronger than that with MFG in which the increase of CFG was with the increase of LAT and the decrease of EI (-0.7669, p<0.05), EC, SC and E. In the association of four haplotype frequencies with CFG and MFG, the association of CAG haplotype frequency with MFG (-0.8633) was significant (p<0.025) while that with CFG (-0.7390) at p<0.1.
DISCUSSION
The differences of meat quality among goat breeds are the results of the long-term natural and artificial selection, the essence of which is the differences of the gene frequencies among the breeds (Chen et al., 2011b). The adaptability of fat metabolism to ecological environment in different goat breeds may be due to the compliant changes of THRSP gene under the double selection pressures whose expression is limited to liver and adipose tissue and is controlled by nutritional and hormonal factors. These conclusions were in agreement with the THRSP gene frequency distribution in this paper. The THRSP interacting with Mid1-interacting protein 1 (Kim et al., 2010a) interferes with Acetyl-coenzyme A carboxylase alpha binding (Brownsey et al., 1997) for the regulation of lipogenesis. Obviously, THRSP was suggested as a transcription factor to regulate gene expression of rate-limiting enzymes in lipogenesis (Decuypere et al., 2005). The polymorphisms of THRSP gene result in the diversity of fatty acid synthesis and lipogenesis in animal tissue. For example, the marbling of longissimus dorsi (Wang et al., 2005) and the muscle fat content were proportional to the expression level of THRSP gene in cattle (Wang et al., 2009; Graugnard et al., 2009; Zhou et al., 2011; Chen, 2010) found the association of pig’s fatty deposit with THRSP gene polymorphism. The fat traits of chicken were related to the polymorphisms of chicken THRSP α gene (D’Andre Hirwa et al., 2009).
The three polymorphism sites of goat THRSP gene in this paper, C-233T, G113A and A138G, might result from the meaningful response of goat to ecological factors and directional selection in body's physiological and biochemical function. The AGGCAGTCAC sequence of C-233T containing an R-box is the binding site of the nuclear factor-kappaB (NF-κB) transcription factor that is found in almost all animal cell types and is involved in cellular responses to stimuli such as stress, ultraviolet irradiation (Wu et al., 2004; Kim et al., 2010b), free radicals (Xu et al., 1999; Baranisrinivasan et al., 2009; Kriete et al., 2010) and cytokines (Salari and Abdollahi, 2011). When C→T at C-233T site, the binding motif of NF-κB changed to AGGCAGTCAT and the R-box disappeared. It might imply that the expresses of THRSP gene relate to the resistance to the stress from the ecological factors. From the haplotype distribution in different goat breeds, the goat breeds with higher EC had higher frequency of CAG haplotype (r = 0.8859, p<0.01) containing NF-κB binding site for the responses to stimuli in habitat such as the increase of ultraviolet irradiation duration while the goat breeds with higher EI had lower frequency of TAG haplotype (r = -0.8377, p<0.025). In the association, moreover, of the CAG haplotype frequency with MFG (r = -0.8633, p<0.025) and CFG (r = -0.7390, p<0.1), the goat breeds with lower frequencies of NF-κB binding sites had more fat in the muscle and carcass. The results in this investigation indicated that the adaptability of goat to ecological environment in habitat depended on the change of THRSP gene in fat metabolism regulation for reducing fatty deposits in muscle and carcass of goat.
CONCLUSION
According to the results of this study, the SC, EC and EI are three ecological factors affecting the frequencies of CAG and TGA haplotypes of goat THRSP gene associated with MFG and CFG levels of goats. EC was significantly proportional to the frequency of CAG haplotype while EI was inversely proportional to the frequency of TGA haplotype. The results suggest that the polymorphism of goat THRSP gene may be associated with the ecological factors and affect the goat fatty deposits ability.
ACKNOWLEDGMENT
The work was supported by The Modern Technology System of National Sheep Industry (nycytx-39), The Modern Technology System of Sheep Industry of Anhui Province and New Varieties of GMO Cultivation of Major Projects of the Ministry of Agriculture (2009ZX08008-007B).
REFERENCES
- Brownsey, R.W., R. Zhande and A.N. Boone, 1997. Isoforms of acetyl-CoA carboxylase: Structures, regulatory properties and metabolic functions. Biochem. Soc. Trans., 25: 1232-1238.
PubMedDirect Link - D'Andre Hirwa, C., W. Paul, W. Yan, C. Luo, Q. Nie, G. Yang and X. Zhang, 2009. Allelic frequency in chicken thyroid hormone responsive spot 14 alfa gene (THRSPα). Asian J. Anim. Sciences, 3: 85-91.
CrossRefDirect Link - Chen, H.Q., H.Q. Wei, J. Qin and H. Chen, 2011. The novel genetic change in 5'-untranslated region of goose prolactin gene and their distribution pattern in different goose breeds. Asian J. Anim. Vet. Adv., 6: 1069-1075.
CrossRefDirect Link - Chen, H.Q., Y.P. Zhang, H. Chen, J. Qin and L.S. Wang, 2011. Polymorphisms in the 5' regulatory region of the gpr54 gene and their association with sexual maturity in cattle. Mol. Reprod. Dev., 78: 894-894.
CrossRefPubMedDirect Link - Chou, W.Y., Y.S. Cheng, C.L. Ho, S.T. Liu and P.Y. Liu et al., 2007. Human spot14 protein interacts physically and functionally with the thyroid receptor. Biochem. Biophys. Res. Commun., 357: 133-138.
Direct Link - Decuypere, E., P. van As, S. van der Geyten and V.M. Darras, 2005. Thyroid hormone availability and activity in avian species: A review. Domest. Anim. Endocrinol., 29: 63-77.
CrossRefDirect Link - Esmaeelkhanian, S., A.J. Aliabad and H. Seyedabadi, 2007. Genetic relationships among six Iranian goat populations based on random amplified polymorphic DNA markers. Pak. J. Biol. Sci., 10: 2955-2959.
CrossRefPubMedDirect Link - Etherton, T.D., 2000. The biology of somatotropin in adipose tissue growth and nutrient partitioning. J. Nutr., 130: 2623-2625.
Direct Link - Fang, Y., W.J. Liu, F.Q. Zhang, Y.G. Shao and S.G. Yu, 2010. The polymorphism of a novel mutation of KAP13.1 gene and its associations with cashmere traits on Xinjian local goat breed in China. Asian J. Anim. Vet. Adv., 5: 34-42.
CrossRef - Graugnard, D.E., P. Piantoni, M. Bionaz, L.L. Berger, D.B. Faulkner and J.J. Loor, 2009. Adipogenic and energy metabolism gene networks in longissimus lumborum during rapid post-weaning growth in Angus and Angus x Simmental cattle fed high- or low-starch diets. BMC Genomics, 10: 142-142.
Direct Link - Hardini, D. and I.H. Djunaidi, 2010. Influence of dietary Bacillus Sp. fermented shrimp waste on broiler meat quality. Int. J. Poult. Sci., 9: 455-458.
CrossRefDirect Link - Askari-Hemmat, H., 2006. A year-by-year event scheduled simulation approach to the design of meat sheep crossbreeding systems. Pak. J. Biol. Sci., 9: 2189-2197.
CrossRefDirect Link - Jin, H., H.Q. Chen, J. Qin, Y.J. Zhu and H. Chen et al., 2012. The polymorphism in 5 regulatory region and exon 13 of PRKAG3 gene and its distribution pattern in different goat breeds. Asian J. Animal Vet. Adv., 7: 568-577.
CrossRefDirect Link - Kim, C.W., Y.A. Moon, S.W. Park, D. Cheng, H.J. Kwon and J.D. Horton, 2010. Induced polymerization of mammalian acetyl-CoA carboxylase by MIG12 provides a tertiary level of regulation of fatty acid synthesis. Proc. Natl. Acad. Sci. USA., 107: 9626-9631.
PubMed - Kriete, A., W.J. Bosl and G. Booker, 2010. Rule-based cell systems model of aging using feedback loop motifs mediated by stress responses. PLoS Comput. Biol., Vol. 6.
CrossRefDirect Link - Orhan, H. and A. Kaygisiz, 2009. Genetic and environmental parameters effecting racing performance of turk-arabian horses raised at anatolian state farm. Asian J. Anima. Vet. Adv., 5: 112-119.
CrossRefDirect Link - Pal, U.K., P.K. Mandal, V.K. Rao and C.D. Das, 2011. Quality and utility of goat milk with special reference to India: An overview. Asian J. Anim. Sci., 5: 56-63.
CrossRefDirect Link - Paracchini, V., P. Pedotti and E. Taioli, 2005. Genetics of leptin and obesity: A HuGE review. Am. J. Epidemiol., 162: 101-114.
Direct Link - Rohde, K., 1992. Latitudinal gradients in species diversity: The search for the primary cause. Oikos, 65: 514-527.
Direct Link - Sebsibe, A., 2008. Sheep and Goat Meat Characteristics and Quality. In: Ethiopia Sheep and Goat Productivity Improvement Program Yami, A. and R.C. Merkel (Eds.), Branna Printing Interprise, Addis Ababa, Ethiopia, pp: 326-340.
Direct Link - Salari, P. and M. Abdollahi, 2011. A comprehensive review of the shared roles of inflammatory cytokines in osteoporosis and cardiovascular diseases as two common old people problem; Actions toward development of new drugs. Int. J. Pharmacol., 7: 552-567.
CrossRefDirect Link - Wakil, S.J. and L.A. Abu-Elheiga, 2009. Fatty acid metabolism: Target for metabolic syndrome. J. Lipid Res., 50: S138-S143.
CrossRefPubMedDirect Link - Wang, Y.H., N.I. Bower, A. Reverter, S.H. Tan and N. De Jager et al., 2009. Gene expression patterns during intramuscular fat development in cattle. J. Anim. Sci., 87: 119-130.
CrossRefPubMedDirect Link - Wang, Y.H., A. Reverter, H. Mannen, M. Taniguchi and G.S. Harper et al., 2005. Transcriptional profiling of muscle tissue in growing Japanese black cattle to identify genes involved with the development of intramuscular fat. Aust. J. Exp. Agric., 45: 809-820.
CrossRefDirect Link - Xu, Y., K.K. Kiningham, M.N. Devalaraja, C.C. Yeh, H. Majima, E.J. Kasarskis and D.K. St Clair, 1999. An intronic NF-kappaB element is essential for induction of the human manganese superoxide dismutase gene by tumor necrosis factor-α and interleukin-1beta. DNA Cell. Biol., 18: 709-722.
PubMed - Zeder, M.A. and B. Hesse, 2000. The initial domestication of goats (Capra hircus) in the Zagros mountains 10,000 years ago. Science, 287: 2254-2257.
CrossRefDirect Link - Zhou, Q.Q., H.Q. Chen, H.Q. Wei, J. Qin, H. Chen and Y.P. Zhang, 2011. Association of polymorphism in the coding region of THRSP gene with lipogenesis capability in pigs. Prog. Biochem. Biophys., 38: 84-90.
Direct Link - Wu, S., M. Tan, Y. Hu, J.L. Wang, D. Scheuner and R.J. Kaufman, 2004. Ultraviolet light activates NfκB through translational inhibition of IκBα synthesis. J. Biol. Chem., 279: 34898-34902.
CrossRefDirect Link