
Anuradha Bhardwaj
Basic and Supporting Discipline Unit, National Research Centre on Equines, Sirsa Road, Hisar-125001, Haryana, India
Varij Nayan
BPR Division, Central Institute for Research on Buffaloes, Sirsa Road, Hisar-125001, Haryana, India
Parvati
Basic and Supporting Discipline Unit, National Research Centre on Equines, Sirsa Road, Hisar-125001, Haryana, India
Mamta
Basic and Supporting Discipline Unit, National Research Centre on Equines, Sirsa Road, Hisar-125001, Haryana, India
A.K. Gupta
Basic and Supporting Discipline Unit, National Research Centre on Equines, Sirsa Road, Hisar-125001, Haryana, India
Asian Journal of Animal and Veterinary Advances
Year: 2012 | Volume: 7 | Issue: 9 | Page No.: 771-789







ABSTRACT
Inhibin is a non-steroidal glycoprotein hormone of gonadal origin with major action as negative feedback control of the production of Follicle Stimulating Hormone (FSH) by anterior pituitary gland which in turn modulates male and female reproductive functions. Its physiological role has led to the development of inhibin based immunogens for fertility enhancement in farm animals. It is envisaged that a reduction of endogenous inhibin secretion would increase FSH concentrations and thus offers a potential for increasing the number of ovulatory follicles in the ovary. Immunization against inhibin has been reported to be a useful method for inducing multiple ovulations in farm animals. Inhibins play important roles in the regulation of fertility based on their dual inhibitory action on the process of folliculogenesis in the ovary and FSH secretion by the pituitary. Inhibins are also recognized as paracrine ovarian and testicular regulators and have multiple paracrine effects in the utero-placental unit, representing a promising marker for male and female infertility, gynecological and gestational diseases.
PDF Abstract XML References Citation
Received: November 05, 2011;
Accepted: March 05, 2012;
Published: May 10, 2012
How to cite this article
Anuradha Bhardwaj, Varij Nayan, Parvati, Mamta and A.K. Gupta, 2012. Inhibin: A Role for Fecundity Augmentation in Farm Animals. Asian Journal of Animal and Veterinary Advances, 7: 771-789.
DOI: 10.3923/ajava.2012.771.789
URL: https://scialert.net/abstract/?doi=ajava.2012.771.789
DOI: 10.3923/ajava.2012.771.789
URL: https://scialert.net/abstract/?doi=ajava.2012.771.789
INTRODUCTION
Livestock plays an important role in Indian economy and productivity in farm species is controlled by many factors. Reproduction is essential for the continuation of the farm species and for most of the life form itself. In biological terms, living and reproducing are essentially one and the same. Reproduction is a complex process, affected by environmental factors and involving a series of physiological events that are properly timed and are supported by endocrine system, through production of several hormones. Environmental factors significantly influence the gestation duration (Meliani et al., 2011). Rizwan et al. (2005) in a short term study found significant differences for LH, FSH, progesterone and estradiol due to occupational exposure to pesticides. The cyclic reproduction pattern is controlled by a finely orchestrated balance and interplay between the hormones of the hypothalamus (GnRH), anterior pituitary (LH, FSH) and gonads (estrogen, testosterone and inhibin), which is classically referred to as the hypothalamo-hypophyseal-gonadal axis. Many hormones work in conjunction with each other to regulate reproductive system in farm animals. Inhibin is a non-steroidal glycoprotein hormone of gonadal origin which selectively inhibits the production of follicle stimulating hormone (FSH) by anterior pituitary gland. Inhibin molecule is a heterodimer composed of two dissimilar subunits of 134 and 116 amino acid residues which are designated as alpha and beta, respectively. Dimerization of these subunits leads to two forms of inhibin: inhibin-A and inhibin-B in which a common alpha subunit is either linked to beta-A or beta-B (αβA = Inhibin-A, αβB =Inhibin-B) produced primarily by gonads, which are processed into a diverse array of different molecular mass αβ dimers (inhibins) and free α and β subunits. Like other members of the transforming growth factor-β gene family, they undergo processing from larger precursor molecules as well as assembly into functional dimers. The concept of inhibin emerged from the studies showing that castration cells appear in the pituitary following damage to seminiferous tubules. McCullagh (1932) first described inhibin in the 1930s and proposed the term to denote the activity of an aqueous extract of the testis that has the capacity to suppress castration cell formation in the anterior pituitary gland. A major breakthrough came, when Frank De Jong and Richard Sharpe in the MRC Reproductive Biology Unit in Edinburgh, UK discovered that fluid from cow ovarian follicles contained high amounts of a substance that suppressed FSH secretion (De Jong and Sharpe, 1976). This provided the source for the eventual purification of inhibin by groups in Australia, Japan and the USA (Miyamoto et al., 1985).
The function of inhibin in the control of FSH secretion suggests that it is an important component of many hormones that control the mammalian reproductive cycle. Accordingly, roles suggested for inhibin in farm animals include maintenance of an appropriate FSH/LH ratio triggering of the secondary surge of FSH in late proestrus and control of folliculogenesis (Hillier, 1991). Apart from their essential role in the selective control of FSH secretion, inhibins are currently recognized as paracrine ovarian and testicular regulators and have multiple paracrine effects in the utero-placental unit, representing a promising marker for male and female infertility, gynecological and gestational diseases. Therefore, in addition to its endocrine role, inhibin may also act as a paracrine/autocrine regulator of gonadal functions. Price et al. (1987) reported that the dimers comprised of inhibin subunits possess diverse functions and may act as growth/differentiation factor as well as hormone.
Optimization of reproduction in females requires control of seasonality and litter size. In species like sheep, goat and cattle, ovulation rate is the major factor limiting the number of young, while uterine capacity is limiting in sows and mares. Hence, techniques such as hormone treatments and GnRH injection (Kalaba and Abdel-Kahlek, 2011; Gharbi et al., 2012), whereby ovulation rate can be increased in amounts compatible with uterine capacity and are valuable tools to maximize offspring numbers from naturally cycling animals. Various methods employing hCG, GnRH, PMSG, interferon and supplementing Omega-3 fatty acids have been tried using different preparations in dairy animals for improving pregnancy rates by reducing embryonic mortality (Bajaj and Sharma, 2011). Research has been directed towards developing inhibin immunization treatment for commercial use in both sheep and cattle (Voglmayr et al., 1992). Researchers were able to demonstrate that ewes could respond to an inhibin vaccine with a sustained (at least 3 years) antibody response and a recurrent increase in litter size. Different types of inhibin based immunogens were used by several workers viz. follicular fluid (steroid free), native/purified inhibin, synthetic inhibin-α peptides and recombinant inhibin α-subunits (Findlay et al., 1993). Bovine follicular fluid in sheep and ovine FF in sheep (Mann et al., 1993; Tannetta et al., 1998) and in cattle have been used as an immunogen to achieve higher FSH levels and higher ovulation rate. Synthetic peptides based on amino acid sequence of immunogenic region of inhibin are proved to be potent immunogens. Synthetic peptides based on bovine inhibin (bIα (1-29) Tyr30) were used to achieve higher ovulation rates in sheep (O'Shea et al., 1994) and in cattle (Glencross et al., 1994). Porcine inhibin-α (1-26) in goat (Medan et al., 2003a) and in cattle (Medan et al., 2005) has been used with positive results. Recently synthetic peptides have been used to increase sperm production in rams (Voge and Wheaton, 2007). A three to five fold increase in ovulation rate was observed in ewes immunized against recombinant bovine and human inhibin alpha subunit fusion proteins (Findlay et al., 1993). Active immunization by recombinant alpha subunit has been demonstrated to increase the ovulation rate up to 2-4 folds in sheep (Forage et al., 1987). Isolation of inhibin from natural sources can only produce limited quantities of bioactive protein. To purify large-scale quantities of recombinant inhibin, recombinant DNA technology provides an eminent tool with production of large amount of recombinant protein in suitable host cell.
INHIBIN FAMILY AND STRUCTURE
Inhibin family comprised of three non-steroidal glycoprotein hormones: Inhibin, activin and follistatin. Inhibins and activins belong to the transforming growth factor-β (TGF-β) superfamily, which mediates embryonic growth and development (Kingsley, 1994). The TGF-β superfamily contains molecules that encompass diverse functions during embryogenesis and adult tissue homeostasis. TGF-beta ligands are initially synthesized as precursor proteins that undergo proteolytic cleavage. The mature segments form dimers via disulfide links, which serve as the active molecule. The subunits (alpha, beta A and beta B) can be processed in several ways to produce different isoforms. Each subunit is produced from a separate gene and is produced as a large precursor protein (Pangas and Woodruff, 2002). Each subunit has multiple cleavage sites, such that subunits of different size are routinely found in follicular fluid (Sugino et al., 1992). The free alpha subunit is often found in higher molar levels than the dimer in many biological fluids (Knight et al., 1989). Various combinations of alpha-beta subunits appear possible, giving rise to a number of different dimeric inhibin forms. Commonly found forms in bovine follicular fluid are approx. 29, 34, 48, 58, 68, 77, 122 and >160 kDa (Ireland et al., 1994). Ganguly et al. (2010) reported two major forms of inhibin in buffalo follicular fluid with a little proteolytic cleavage/processing of the larger precursor. Within the TGF-β superfamily, the agonist-antagonist relationship between activin and inhibin is unique and critical to integrated reproductive function. Activin acts at the pituitary to stimulate follicle-stimulating hormone (FSH) and is antagonized by endocrine acting gonadally-derived inhibin (Mather et al., 1992).
Activin only has inhibin-beta subunits: Activin-A has two beta-A subunits; activin-B has two beta-B subunits; Activin-AB has beta-A and beta-B subunits. These beta subunits are expressed in the same cells as inhibin beta subunits. Activin stimulates FSH and LH production and/or secretion from the pituitary gland (DePaolo et al., 1991).
Follistatin (FS) is not a member of the TGF-β superfamily. It exists in two monomeric forms of 288 and 315 kDa. FS was discovered in gonadal extracts as an inhibitor of FSH secretion from pituitary gonadotrophs. Follistatin suppresses FSH by irreversible binding and neutralization of activins (Vale et al., 1988) (Fig. 1).
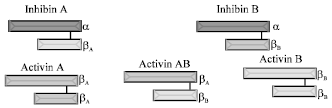 | |
| Fig. 1: | Various molecular forms of inhibins and activins |
PHYSIOLOGICAL/BIOLOGICAL ROLE
In mammals, inhibin is predominately expressed in the testis [sertoli cells] and ovary [granulosa and theca cells] (Robertson et al., 1991; Krummen et al., 1994). Ovarian follicular fluid (FF) is a rich source of inhibin. The physiological role of inhibin in reproduction i.e., regulation of folliculogenesis in females and spermatogenesis in males, can be classified into three types-endocrine, paracrine and autocrine (Findlay, 1993; Knight and Glister, 2001). Inhibin has been shown to exert a negative feedback effect on follicle-stimulating hormone (FSH) at the pituitary gland (Schwartz and Channing, 1977) and to affect gonadal function (Findlay et al., 1993). Apart from their essential role in the selective control of FSH secretion, inhibins are also recognized as paracrine ovarian and testicular regulators and have multiple paracrine effects in the utero-placental unit, representing a promising marker for male and female infertility, gynecological and gestational diseases. In vitro studies have indicated two separate mechanisms of action of inhibin on FSH secretion: at low concentrations, inhibin suppressed FSH release and synthesis while at higher concentrations, the pituitary gonadotroph content of both FSH and LH is reduced due to degradation of intracellular stores of these hormones (Burger, 1992). Various studies indicate that inhibin regulates long term mean levels by producing an overall damping effect.
Physiological role in females: The role of inhibin as an important component in the feedback regulation of FSH secretion from anterior pituitary and thus in folliculogenesis is well established by various researchers.
Functional relationships among FSH, follicles and inhibin and regulation of ovarian follicular development: FSH is the key hormone inducing the recruitment and growth of ovarian follicles. An association between a FSH surge and recruitment of follicles has been demonstrated by various workers in the course of development of reproductive biology (Kaneko et al., 1995; Medan et al., 2005, 2003a). During the estrous cycle of cows (Kaneko et al., 1995, 2002) and goats (Medan et al., 2003a; Medan et al., 2005), periodic fluctuations of FSH concentrations are responsible for the regular emergence of follicular waves (Fig. 2). The rate of reduction in FSH concentrations is directly related to the number of growing cohort follicles. Thus, secretion of inhibins from the growing cohort follicles into the systemic circulation is probably responsible for the decline of FSH, indirectly causing their own atresia (Medan et al., 2007). FSH also stimulates the production of inhibin and follistatin.
Various experiments confirm the involvement of inhibin in the regulation of FSH secretion during the growth phase of the dominant follicle in the early luteal phase in females. Inhibin neutralization during the early luteal phase, produces hypersecretion of FSH with an associated stimulation of follicular development, indicating that inhibin is an important factor for the negative regulation of FSH secretion during the early luteal phase when secretion of estradiol and progesterone are normally high (Hillier, 1991). By mid follicular phase, in the follicle responding most rapidly and extensively to FSH, granulosa cell aromatase activity, inhibin production and LH receptor expression, rise to critical levels. Since aromatase activity and inhibin production are coupled to the LH receptor, this follicle is selected and becomes destined to ovulate. During the second half of follicular phase, inhibin production in the preovulatory follicle continues to increase, paralleling the aromatase activity and estrogen secretion (Hillier, 1991). After induction of luteolysis, FSH stimulates follicular growth resulting in an increase in inhibin and estradiol-17β secretion by the dominant follicle, leading to reduced peripheral FSH levels.
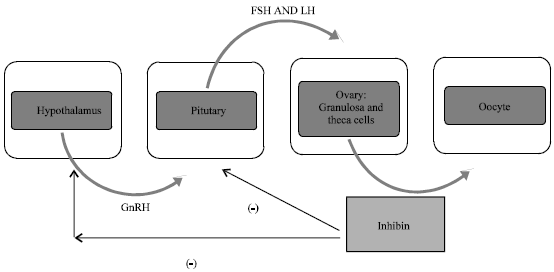 | |
| Fig. 2: | Hypothalamo-hypophyseal axis-role of hormones |
This results in suppression of subordinate follicles from the same wave of recruited follicles.
Secretion of inhibins during the estrous cycle: Inhibins are produced by follicles as they develop from small antral to preovulatory stages (Medan et al., 2007). Follicle turnover continues throughout estrous cycle in a wave-like pattern in domestic animals, such as cattle (Welt et al., 1997; Kaneko et al., 1995), mares (Buratini et al., 1997), sheep (Bartlewski et al., 1999) and goats (Medan et al., 2003b; Medan et al., 2005). The emergence of each follicular wave is preceded by an elevation in FSH secretion. Circulating FSH levels decline after the selection of the dominant follicle(s) through a negative feedback effect of the inhibins produced by the developing follicles. In the golden hamster, Ohshima et al. (1999) found that inhibin A increased from the early morning of day 1 (day 1 = Day of ovulation) and reached plateau levels on day 2, being followed by an abrupt increase at the time of LH surge on day 4. However, inhibin B increased on day 1 and declined on day 4. These differential patterns of inhibin A and B secretion in the hamster were also observed after induced atresia and subsequent follicular development (Ohshima et al., 2002). These findings suggest that the dominant form of dimeric inhibins may shift from inhibin B to inhibin A with follicular development. Also, the distinct patterns of inhibin A and inhibin B secretion suggest the unique regulation of inhibin A and inhibin B by gonadotropins and the stage of follicular development. These differences in regulation are apparent when comparing stimulation of secretion in vivo and in vitro. In vivo, FSH readily stimulates both inhibin A and inhibin B secretion in the early follicular phase, when small antral follicles are present (Welt et al., 1997; Welt and Schneyer, 2001). In contrast, while both FSH and cyclic adenosine monophosphate (cAMP) stimulate inhibin A secretion from the granulosa cells of small antral follicles in vitro, neither stimulates inhibin B. Furthermore, both FSH and LH stimulate inhibin A from the preovulatory follicle but neither stimulates inhibin B in vivo (Welt et al., 1999, 2001). Consistent with the hypothesis that FSH stimulates inhibin A but not inhibin B, inhibin A increases in follicular fluid with increasing follicle maturity, whereas inhibin B does not (Schneyer et al., 2000).
In cattle (Kaneko et al., 2002) and goats (Medan et al., 2003b; Medan et al., 2005) an inverse relationship between FSH and inhibin A was demonstrated, indicating the key role of inhibin A produced by dominant follicle(s) in terminating the transient peaks of FSH secretion. There was a highly significant inverse correlation between inhibin A and FSH and a highly significant positive correlation between inhibin A and estradiol-17β as the mean concentrations of estradiol-17β increased during the follicular phase and reached a peak 2 days before ovulation. These findings clearly indicate that follicles increase the secretion of inhibin A as well as estradiol-17β during final maturation in small ruminants. Moreover, there is a unique phenomenon in the secretion of inhibin, the so-called ovulatory inhibin surge, during the estrous cycle in farm animals such as mares, goats etc. Circulating inhibins (ir-inhibin, inhibin pro-αC and inhibin A) but not estradiol-17β sharply increases on the day of ovulation in mares (Nambo et al., 2002). These findings suggest that circulating inhibins may be useful for determining the time of ovulation in the farm animals.
Inhibins and activins during pregnancy: Circulating levels of inhibin and activin during pregnancy have been reported in human and non-human primates. Also, both inhibin βA and inhibin βB subunit mRNAs and associated proteins have been identified in placentas, deciduas and fetal membranes (Kondo et al., 2003). Placentas and fetal membranes in humans (Florio et al., 2001) and non-human primates (Kondo et al., 2003) are major sites of the production and secretion of inhibin and activin in maternal serum and amniotic fluid. The levels of inhibin A and activin A significantly decreased after the removal of the feto-placental unit (Muttukrishna et al., 1997). There are a few reports on the secretion of inhibins and activins from the placenta in species other than primates. In the rat and hamster, ovarian follicles but not placentas are major source of circulating inhibin, whereas the placenta secretes a large amount of activin A in the hamster (Ohshima et al., 2002). On the other hand, equine female ovaries and testes secrete large amounts of inhibin pro-αC and inhibin A but not activin during late pregnancy (Ohshima et al., 2002; Kondo et al., 2003).
Physiological role in males: Physiological inhibin production by the adult testis requires a normal population of sertoli cells, FSH stimulation and spermatogenesis to be present. The two later factors are not absolutely necessary for a basal inhibin B release, which is seen in some forms of hypogonadism, impaired spermatogenesis (Foresta et al., 1999). Not only is the contribution of leydig cells to the circulating inhibin negligible (Anderson et al., 1998),but the LH effect on inhibin release also appears to be inhibitory rather than stimulatory (Ramaswamy and Plant, 2001).In humans, among infertile males with elevated FSH levels, the FSH concentration is inversely correlated with inhibin B but not with pro-αC inhibin, suggesting that the physiologically important hormone that exerts potential negative feedback upon FSH secretion is inhibin B (Illingworth et al., 1996). In male primates (Illingworth et al., 1996) and rodents (Woodruff et al., 1996; Jin et al., 2001), inhibin B is known or believed to be the primary dimeric form in the blood circulation. Inhibin concentration in rams is increased in late fetal life and in the first few months after birth (Sanford et al., 2000) when sertoli cells are completing proliferation. A comparatively small increase in inhibin occurs in mature rams during seasonal testicular recrudescence (Sanford et al., 2000). Inhibin is involved in suppression of FSH secretion in late fetal life, during puberty and in adulthood (Tilbrook et al., 1999). Furthermore, dimeric inhibin and forms of the α-subunit of inhibin may act as paracrine/ autocrine regulators in the testis (Chong et al., 2000). In rams, FSH administration increases inhibin production by fetal testis and the output of inhibin into the testicular lymphatic and vascular systems in adults (Voglmayr et al., 1992). Corresponding increases in FSH, immunoactive inhibin B (Sanford et al., 2000) and inhibin A (Lincoln et al., 2001) during testicular redevelopment and greater numbers of FSH receptors, in the testis in the breeding season compared with the non-breeding season, provide additional evidence for FSH regulation of inhibin secretion in rams. However, the mode of FSH action (altered secretion versus number of receptors) and the regulation of gene expression for FSH receptors as testes pass through the different stages of the seasonal cycle are poorly understood. Inhibin secretion by sertoli cells may also be regulated by paracrine mechanisms acting within the testis. Sertoli cells in rodent species and human males have androgen receptors and normal cell function requires both testosterone and FSH (Baird and Smith, 1993). Thus, testosterone from neighboring leydig cells is a candidate for inhibin regulation. An effect of testosterone may also be mediated indirectly by other androgen target cells such as peritubular myoid cells. Other studies concerned mainly with sertoli cells in culture denote conflicting indications as to whether testosterone stimulates or inhibits inhibin secretion. Sertoli cells and peritubular cells in ram testis contain androgen receptors and seasonal variations in blood inhibin and testosterone concentrations are positively correlated (Sanford et al., 1993). Razie et al. (2011) studied the inter-relationship between the level of inhibin B and ultra-structure of sertoli cells in contra-lateral testis after unilateral blunt testis trauma in rats and suggested that UTT affected the contra-lateral testis and levels of inhibin B in serum reflected the spermatogenetics and other functions of sertoli cells.
IMPORTANCE OF INHIBIN IN REPRODUCTION MANIPULATION
The combination of equine chorionic gonadotropin and human chorionic gonadotropin has been a common method to induce superovulation in farm animals (Rahman et al., 2008; Moeini et al., 2009) except equines. The pattern of folliculogenesis, eCG and progesterone profile was studied in cyclic, pregnant and irregular cyclic equine mares (Pal and Gupta, 2005; Meenakshi et al., 2006, 2008; Bansal et al., 2006, 2009). It has been demonstrated by various workers that uterine capacity is the major limiting factor for increasing the number of offspring in equines. The assisted reproductive technologies in Farm animals have also contributed significantly in increasing the productivity (Sejian et al., 2010). Inhibin acts directly on pituitary to inhibit FSH synthesis and secretion. A reduction of endogenous inhibin secretion would increase FSH concentrations and thus offers a potential for increasing the number of ovulatory follicles in the ovary. Thus inhibin mediated pathway also found to be potential alternate methodology for superovulation (Findlay et al., 1993; Palta, 1998; Medan et al., 2007).
Practical uses of inhibin as fecundity augmenting agent: Inhibins act as chemical signals to the pituitary gland on the number of growing follicles in the ovary. Inhibins reduce the secretion of FSH to a level, which maintains the species-specific number of ovulation in both single and litter bearing species. By inhibiting FSH release without altering LH release, inhibins may be partly responsible for the differential release of FSH and LH from the pituitary (Hafez and Hafez, 2000). Inhibin in the form of steroid-free follicular fluid preparation or purified bovine inhibin results in the specific suppression of plasma concentrations of FSH in the ewes. This suppression of FSH is associated with a failure of growth of large preovulatory follicles greater than 2.5 mm diameter. On cessation of FF treatment, a rebound release of FSH results in an increase in ovulation rate in the subsequent cycle. These effects are mediated by the direct inhibitory effect of inhibin on the secretion of FSH from pituitary. However, the follicular fluid contains not only inhibin but also many other proteins that could be immunoreactive and the results were variable and inconsistent. Bovine follicular fluid in sheep (Cumminst et al., 1986) and ovine FF in sheep (McNeilly, 1984; McNeilly et al., 1991; Tannetta et al., 1998) and in cattle (Price et al., 1987; Bindon et al., 1988) have been used as an immunogen to achieve higher FSH levels and higher ovulation rate. In early days, native inhibin, purified from follicular fluid, has also been used; bovine inhibin in ewes (Henderson et al., 1984), sheep inhibin in cow (Price et al., 1987) and ovine inhibin in cow (O'Shea et al., 1994) resulted in higher FSH levels and ovulation rate. Purification of inhibin from follicular fluid involved a costly and cumbersome process; moreover, purity or immunoreactivity may be less in these cases as compared to synthetic peptides and recombinant a-inhibin proteins.
Immunization of ewes against recombinant bovine inhibin α-subunit (expressed in prokaryotic system) resulted in increased basal levels of FSH in the luteal phase and GnRH-stimulated concentrations of FSH in the follicular phase of the oestrous cycle (Findlay et al., 1989), though LH levels were unchanged and neither gonadotrophin was affected by immunization during anoestrous. Unconjugated α-βI as an immunogen elicited a strong immune response, reflected by the presence of antibodies in the ewes capable of recognizing native inhibin. It implies that the epitopes present in the non-glycosylated α-subunit expressed by the prokaryotes are also present in the native molecule, presumably on the glycosylated a-subunit. Increase in ovulation rate of three fold were achieved in sheep immunized with a recombinant fusion protein of α-βI-(165-300 amino acid) with an N-terminal extension of 20 amino acid of β-galactosidase (Forage et al., 1987), whilst use of a recombinant human α-inhibin to immunize Rambouillet ewes led to ovulation rates 4-6 times greater than in controls (Mizumachi et al., 1990). The latter study also reported a concomitant enhancement of both pre- and postovulatory levels of FSH, but not LH, the postovulatory surge also persisting for longer periods of time in immunized animals. In guinea pigs, immunized with recombinant ovine alpha inhibin, Shi et al. (2000a) observed increase in ovulation rate in a dose dependent manner and suggested that immunization results in the production of circulating antibodies that are able to bind to endogenous native guinea pig inhibin. Use of recombinant bINH-α to immunize gilts (Brown et al., 1990) led to rise in mean ovulation rates from 12 to over 16, the number of ovulations being highly correlated with levels of inhibin antibodies present in the serum. The cloning and heterologous expression of Indian Sahiwal cattle (Bos indicus) bINH-α (bovine alpha inhibin) encoding gene was successfully done in E. coli and the purified recombinant bINH-α was characterized (Bhardwaj et al., 2007; Bhardwaj, 2008). Recombinant bINH-α (25 μg mL-1) immunized guinea pigs had a significant increase in litter size compared to control group. These results also indicated a role for recombinant bovine alpha inhibin as a fecundity vaccine to enhance the ovulation rate and litter size in animals (Bhardwaj et al., 2006, 2012).
A chimeric recombinant ovalbumin/inhibin (ovalin) vaccine was prepared by Geary and Reeves, 1996. Rabbits were immunized subcutaneously against crude ovalin (300 μg of antigen) and significant antibody titers were observed against ovalbumin and inhibin. Similarly, a novel chimeric antigen (Sewani et al., 1998) consisting of the non-toxic B subunit (EtxB) of an E. coli enterotoxin and the first 14 N-terminal amino acid residues of the C-terminal portion of the α-subunit of bovine inhibin (bINH1-14) was produced in prokaryotic host. Rabbits immunized subcutaneously with EtxB::bINH1-14 developed significant titers of antibodies that recognized an inhibin peptide fragment containing bINH1-14, native inhibins and EtxB during separate enzyme-linked immunosorbent assay (ELISA). Passive immunization of mice with the rabbit anti-EtxB::bINH1-14 serum increased concentrations of follicle-stimulating hormone (FSH) in serum twofold compared with controls, whereas serum concentrations of luteinizing hormone (LH) were unaltered.
The effect of selective immunosuppression of endogenous inhibin in goats on FSH, LH, progesterone and estradiol-17β profiles during the breeding and non-breeding seasons by immunization against the recombinant human inhibin α-subunit (hINH-α) was studied by Hennies et al. (2001). They found that, mean basal concentrations of FSH were not affected by immunosuppression of endogenous inhibin, nor was there a difference in the amplitude of the pre-ovulatory FSH surge. By contrast, concentrations of circulating estradiol were significantly elevated after immunization. Extending immunization into the anoestrous season by a booster injection of hINH-α, implicating oestrus induction with a progestogen and eCG, produced no discernible differences in FSH and LH profiles in comparison with non-immunized control goats. Their findings suggest that, paracrine factors may play a more significant role in controlling follicular activity than a feedback mechanism acting via the pituitary. In one study, Findlay et al. (1993) reported that active immunization against a fusion protein consisting of amino acid 1-166 of the alpha subunit (αN fragment) of bovine inhibin led to impairment of fertility in sheep in terms of lambs born. This could not be attributed to changes in circulating inhibin, progesterone or gonadotrophin levels and the number of corpora lutea were greater than control animals. However, corpora lutea had the appearance of luteinized unruptured follicles and egg recovery was significantly lower, thereby indicating ovulation failure in these animals and implying a facilitatory role for αN fragment in the ovulatory process (Terqui et al., 1995).
Immunization of ewes against an synthetic N-terminal fragment (1-29) of bovine inhibin α-subunit conjugated to tuberculin purified protein derivative resulted in a twofold rise in ovulation rate during the breeding season, 25% increase in FSH levels and 37% increase in lambs born. Although, conception and pregnancy rates and length of gestation were unaffected, lambs born to immunized ewes had lower birth weights and a higher proportion was stillborn. Thus, overall there was no significant increase in the number of viable lambs produced (Wrathall et al., 1990). Kaneko et al. (2002) showed that a dominant follicle during luteal phase secreted inhibin and the peripheral concentration of inhibin increased according to the growth of dominant follicles in cows. The number of ovulations is reduced, when a superovulatory treatment is initiated in the presence of a functional dominant follicle and removing the dominant follicle improves the superovulatory response. Inhibin suppresses the development of FSH sensitive follicles or the response to superovulatory treatment by inhibiting the secretion of endogenous FSH. Active Immunization of goats (Medan et al., 2003b) and cattle (Takedomi et al., 2005) against inhibin (porcine αN-1-26 conjugated to rabbit serum albumin) resulted in significant increase in ovulation rate. These results have attributed to the stimulatory effect of inhibin immunoneutralisation on the ovary, either endocrinologically, by removing the inhibitory effect of inhibin on FSH secretion or locally, through a paracrine effect and finally enhancing ovarian follicular development and increasing the number of growing follicles.
On the other hand, passive immunization against inhibin also increased the ovulation rate through elevated FSH secretion in mice (Medan et al., 2004a), hamsters (Kishi et al., 1997), rats (Rivier and Vale, 1989), guinea pigs (Shi et al., 2000b), ewes (Mann et al., 1993), goats (Medan et al., 2003b, 2004b), cows (Akagi et al., 1997; Takedomi et al., 1997) and mares (Nambo et al., 1998). In cattle, i.v., injections of 25, 37.5 and 50 mL of inhibin antiserum (against inhibin produced in a castrated male goat) were given and multiple ovulations (2-4) were recorded in all animals after injection of 50 mL inhibin antiserum, however all cows in 25 mL group experienced only one ovulation and injection of 37.5 mL resulted in a variable number of ovulations (1-5) (Akagi et al., 1997). In rats, i.p., injection of inhibin antiserum (obtained from a castrated goat immunized against Tyr30-porcine inhibin-α (1-30)-NH2 conjugated to rabbit serum albumin) at doses- 50, 100, 200, 400 μL was administered and higher ovulation rate was achieved in dose dependent manner (Wang et al., 2001). The rate of blastocyst development for animals treated with 50-200 μL inhibin antiserums was significantly higher than control animals (Wang et al., 2001). The oocytes induced to superovulate using immunization against endogenous inhibin have normal developmental competence (Medan et al., 2007).
In another study, two experiments were conducted to induce superovulation in goats using passive and active immunization against inhibin (Sasaki et al., 2006). There was a significant increase in plasma FSH concentration compared with the controls. The numbers of follicles in passively and actively immunized goats were significantly greater than those in the controls. In addition, the ovulation rate was greater in the immunized animals compared with the controls. Therefore, either passive or active immunization against inhibin could be used to induce superovulation.
In males, immunization against inhibin immunogens increases FSH and testis functions (Terqui et al., 1995). In rams, immunization against inhibin increased testis diameter and daily sperm output in one study (Al-Obaidi et al., 1987), whereas body weight, scrotal circumference and plasma FSH were unaffected in another (Wheaton and Godfrey, 2003). However, in the latter study, the age at puberty was delayed and Luteinizing Hormone (LH) and testosterone were reduced, in the inhibin-immunized lambs. In rams, immunomodulation of inhibin led to delayed, amplified and extended season related increases in serum gonadotropins (Voglmayr et al., 1990). Al-Obaidi et al. (1987) observed significant rise in both testis diameter and daily sperm production in rams after inhibin immunization. Whereas, in bulls, Kaneko et al. (2002) found levels of plasma immunoreactive inhibin and FSH to be relatively low for the first few weeks of life and then to rise steadily thereafter, treatment at a young age with inhibin antiserum (passive immunization) produced elevations in plasma FSH without affecting. Schanbacher (1991) showed enhancements in serum FSH and testicular sperm density, although testes and epididymides weight, daily sperm production and blood levels of LH and testosterone were not altered. Similarly, Martin et al. (1991) found active immunization against inhibin to increase serum levels of FSH but ineffective in altering testicular growth (measured as scrotal circumference). However, unlike Schanbacher (1991), Martin et al. (1991) found inhibin immunoneutralization increased circulating concentrations of testosterone and daily sperm production along with a decrease in blood levels of LH. In another study, Holstein bulls (Bame et al., 1999) were actively immunized against bovine inhibin α1–26 gly-tyr (bINH) conjugated to human alpha globulin and enhancement of sperm output after immunization against inhibin was observed LH. Satterlee et al. (2006) found that, injection of quail and breeder hens with a recombinant protein antigen (MBP-cINA521)-a fusion of the bacterial maltose-binding protein (MBP) and a fragment of the a-subunit of chicken inhibin (cINA521) accelerate puberty and enhances lay. The gonadal hormone inhibin regulates daily sperm production (DSP) indirectly through negative feedback control of FSH secretion and may also affect DSP via direct actions within the testis (Voge and Wheaton, 2007).
Hence, immunization against inhibin in the males and females increases FSH secretion with no significant changes in LH. However, in addition to the classical endocrine feedback role of inhibin, inhibin molecules also subserve local actions at the intragonadal. Such local actions may also be perturbed following inhibin immunization. Moreover, consideration must be given to the natural fecundity of the species or breeds to be immunized, since although technically feasible, there may be little to be gained by artificially raising litter size beyond the optimum. Maternal behaviour and milk production, for example, have to be adequate to raise extra litters. As with other approaches to increase fecundity (genetic selection, gonadotrophin treatment etc.), the increased ovulation rates resulting from inhibin immunization may also have a detrimental effect on perinatal survival owing to decreased birth weight.
Other clinical uses of inhibin
Inhibin measurements in ovarian and gestational disorders: One of the most important developments in the field of reproduction is the use of inhibin as an ovarian tumor marker. Inhibin α-subunit and dimeric inhibin A and B have been detected in the serum of females with granulosa cell tumors and epithelial ovarian cancers (Burger et al., 2001). In farm animals, studies were conducted in mares only for the detection of tumors with inhibins. Granulosa theca cell tumors (GTCT) in the mare secrete high levels of ir-inhibin (Yoshida et al., 2000; Bailey et al., 2002); therefore measuring inhibins may be useful in diagnosing and confirming GTCT. Davis et al. (2005) reported that two cases of GTCT in mares displayed positive immunoreactivity for inhibin α-subunit, but were negative for inhibin βA and βB subunits. This indicates that the appearance of the inhibin α-subunit can be used as an immunohistochemical marker in the diagnosis of GTCT in the small ruminants.
Several reports have measured inhibins and activins in pregnancy disorders (studies yet limited to humans). Inhibin A and activin A levels were elevated in women carrying a child with Down syndrome (Dalgliesh et al., 2001) and the levels of α, βA mRNAs also increased in the placentas of Down syndrome pregnancies (Lambert-Messerlian et al., 1998). Pre-eclampsia is a pregnancy specific disease that is a major cause of maternal and fetal morbidity. Inhibin A levels in the maternal serum of women who developed pre-eclampsia were higher than that of controls (Aquilina et al., 1999). Studies investigating the source responsible for the rise in serum activin A and inhibin A in pre-eclampsia have reported an increased expression of inhibin α and inhibin/activin β subunit genes (Florio et al., 2002; Casagrandi et al., 2003) and proteins (Manuelpillai et al., 2001; Bersinger et al., 2002) in the placenta. Serum inhibin and activin levels were found to be high in patients with the hydatidiform mole (Florio et al., 2002), suggesting that inhibins and activins can be used as reliable tumor markers. In addition, measurement of inhibins is useful in hypothalamic amenorrhoea (Welt et al., 1997), polycystic ovarian syndrome (Lockwood et al., 1998) and premature ovarian failure (Petraglia et al., 1991). Inhibin B has been found to be a reliable marker in ovarian aging and the menopause transition (Sowers et al., 2008).
CONCLUSION AND FUTURE PERSPECTIVES
Inhibins are multifunctional molecules involved in the control of pituitary FSH secretion. They are glycoprotein hormones produced in the gonads and are capable of regulating FSH secretion by the pituitary gland. Inhibins show a reciprocal relationship with FSH during estrous cycle. Immunization against inhibin enhances FSH secretion and follicular growth and finally increases the ovulation rate. The different studies on inhibin for increasing ovulation rate and fecundity in animals opens the possibility of eliminating the use of exogenous gonadotropins for fertility enhancement. The significance of inhibin as a marker in other pregnancy related and ovarian functions paves the way for further study and research.
REFERENCES
- Akagi, S., H. Kaneko, Y. Nakanishi, T. Takedomi, G. Watanabe and K. Taya, 1997. Ovarian response and FSH profile in cows following injection of various doses of inhibin antiserum. J. Vet. Med. Sci., 59: 1129-1135.
CrossRef - Al-Obaidi, S.A.R., B.M. Bindon, M.A. Hillard and T. O'Shea, 1987. Reproductive characteristics of lambs actively immunized early in life with inhibin-enriched preparations from follicular fluid of cows. J. Reprod. Fertil., 81: 403-414.
Direct Link - Anderson, S.T., B.M. Bindon, M.A. Hillard and T. O'Shea, 1998. Increased ovulation rate in Merino ewes immunized against small synthetic peptide fragments of the inhibin a-subunit. Reprod. Fertil. Dev., 10: 421-431.
Direct Link - Aquilina, J., A. Barnett, O. Thompson and K. Harrington, 1999. Second-trimester maternal serum inhibinA concentration as an early marker for preeclampsia. Am. J. Obstet. Gynecol., 181: 131-136.
CrossRef - Bajaj, N.K. and N. Sharma, 2011. Endocrine causes of early embryonic death: An overview. Curr. Res. Dairy Sci., 3: 1-24.
CrossRefDirect Link - Ganguly, A., I. Ganguly and S.K. Meur, 2010. Multiple molecular forms of inhibin in buffalo (Bubalus bubalis) ovarian follicular fluid. Res. Vet. Sci., 89: 14-19.
CrossRef - Bailey, M.T., M.H.T. Troedsson and J.E. Wheaton, 2002. Inhibin concentrations in mares with granulosa cell tumors. Theriogenology, 57: 1885-1895.
CrossRef - Baird, D.T. and K.B. Smith, 1993. Inhibin and related peptides in the regulation of reproduction. Oxf. Rev. Reprod. Biol., 15: 191-232.
PubMed - Bame, J.H., J.C. Dalton, S.D. Degelos, T.E.M. Good and J.L.H. Ireland et al., 1999. Effect of long-term immunization against inhibin on sperm output in bulls. Biol. Reprod., 60: 1360-1366.
Direct Link - Bersinger, N.A., N.P. Groome and S. Muttukrishna, 2002. Pregnancy-associated and placental proteins in the placental tissue of normal pregnant women and patients with pre-eclampsia at term. Eur. J. Endocrinol., 147: 785-793.
Direct Link - Bhardwaj, A., S. De, T.K. Datta, S.L. Goswami, Y. Yadav, M. Thakur and P. Yadav, 2006. Recombinant Inhibin: A step towards fertility augmentation in farm animals. Proceedings of National Symposium on Frontiers in Reproduction: Concepts and Applications in Genomic Era, February 23-25, 2006, Karnal, India.
- Bhardwaj, A., V. Nayan, P. Yadav, S. De, T.K. Datta and S.L. Goswami, 2012. Evaluation of biological efficacy of recombinant bovine inhibin-alpha in guinea pigs. Proceedings of the National Symposium on Recent Advances in Reproductive Biotechnology: Retrospective and Prospective Vision, January 30-31, 2012, Karnal, India,.
- Brown, R.W., J.W. Hungerford, P.E. Greenwood, R.J. Bloor, D.F. Evans, C.G. Tsonis and R.G. Forage, 1990. lmmunization against recombinant bovine inhibin a subunit causes increased ovulation rates in gilts. J. Reprod. Fertil., 90: 199-205.
Direct Link - Buratini, J., A.A.M.R. e Silva, C.M.Q. Barros, F.O. Papa, M.C.S. Caldas and C. Meira, 1997. Follicular dynamics in Mangalarga mares. Equine Vet. J., 29: 7-11.
CrossRef - Casagrandi, D., C. Bearfield, J. Geary, C.W. Redman and S. Muttukrishna, 2003. Inhibin, activin, follistatin, activin receptors and b-glycan gene expression in the placental tissue of patients with pre-eclampsia. Mol. Hum. Reprod., 9: 199-203.
CrossRef - Chong, H., S.A. Pangas, D.J. Bernard, E. Wang and J. Gitch et al., 2000. Structure and expression of a membrane component of the inhibin receptor system. Endocrinology, 141: 2600-2607.
CrossRef - Cumminst, L.J., T. O`Shea, S.A.R. Al-Obaidi, B.M. Bindon and J.K. Findlay, 1986. Increase in ovulation rate after immunization of Merino ewes with a fraction of bovine follicular fluid containing inhibin activity. J. Reprod. Fertil., 77: 365-372.
CrossRef - Dalgliesh, G.L., D.A. Aitken, F. Lyall, A.G. Howatson and J.M. Connor, 2001. Placental and maternal serum inhibin-a and activin-a levels in Down`s syndrome pregnancies. Placenta, 22: 227-234.
CrossRef - Davis, W.P., M.S. Medan, W.Z. Jin, R.E. Wells, G. Watanabe and K. Taya, 2005. Immunohistochemical localization of inhibin a-subunit in two equine granulosa-theca cell tumors. J. Equine Sci., 16: 45-49.
CrossRef - De Jong, F.H. and R.M. Sharpe, 1976. Evidence for inhibin-like activity in bovine follicular fluid. Nature, 263: 71-72.
CrossRef - DePaolo, L.V., T.A. Bicsak, G.F. Erickson, S. Shimasaki and N. Ling, 1991. Follistatin and activin: A potential intrinsic regulatory system within diverse tissues. Proc. Soc. Exp. Biol. Med., 198: 500-512.
PubMedDirect Link - Findlay, J.K., 1993. An update on the roles of inhibin, activin and follistatin as local regulators of folliculogenesis. Biol. Reprod., 48: 15-23.
PubMedDirect Link - Findlay, J.K., B. Doughton, D.M. Robertson and R.G. Forage, 1989. Effects of immunization against recombinant bovine inhibin a subunit on circulating concentrations of gonadotrophins in ewes. J. Endocrinol., 120: 59-63.
CrossRef - Findlay, J.K., B.W. Doughton, C.G. Tsonis, R.W. Brown, J.W. Hungerford, P.E. Greenwood and R.G. Forage, 1993. Inhibin as a fecundity vaccine. Anim. Reprod. Sci., 33: 325-343.
Direct Link - Florio, P., L. Cobellis, S. Luisi, P. Ciarmela, F.M. Severi, C. Bocchi and F. Petraglia, 2001. Changes in inhibins and activin secretion in healthy and pathological pregnancies. Mol. Cell. Endocrinol., 180: 123-130.
CrossRef - Florio, P., F.M. Severi, L. Cobellis, S. Danero, A. Bome, S. Luisi and F. Petraglia, 2002. Serum activin A and inhibin A. New clinical markers for hydatidiform mole. Cancer, 94: 2618-2622.
CrossRef - Forage, R.G., R.W. Brown, K.J. Oliver, B.T. Atrache and P.L. Devine et al., 1987. Immunization against an inhibin subunit produced by recombinant DNA techniques results in increased ovulation rate in sheep. J. Endocrinol., 114: Rl-R4.
CrossRef - Foresta, C., A. Bettella, F. Petraglia, M. Pistorello, S. Luisi and M. Rossat, 1999. Inhibin B levels in azoospermic subjects with cytologically characterized testicular pathology. Clin. Endocrinol., 50: 695-701.
CrossRef - Glencross, R.G., E.C. Bleach, S.C. Wood and P.G. Knight, 1994. Active immunization of heifers against inhibin: effects on plasma concentrations of gonadotropins, steroids and ovarian follicular dynamics during prostaglandin-synchronized cycles. J. Reprod. Fertil., 100: 599-605.
PubMed - Henderson, K.M., P. Franchimont, M.J. Lecomte-Yerna, N. Hudson and K. Ball, 1984. Increase in ovulation rate after active immunization of sheep with inhibin partially purified from bovine follicular fluid. J. Endocrinol., 102: 305-309.
PubMed - Hennies, M., J.K. Voglmayr, E. Dietrich, M. Stollmann, R. Moeller, and W. Holtz, 2001. Hormonal response of female goats to active immunization against a recombinant human inhibin a-subunit and establishment of an enzyme-linked immunosorbent assay for caprine folliclestimulating hormone. Reprod. Domest. Anim., 36: 65-71.
CrossRef - Hillier, S.G., 1991. Regulatory functions for inhibin and activin in human ovaries. J. Endocrinol., 131: 171-175.
CrossRef - Ireland, J.L.H., Good, T.E.M., Knight, P.G. and J.J. Ireland, 1994. Alterations in amounts of different forms of inhibin during follicular atresia. Biol. Reprod., 50: 1265-1276.
Direct Link - Illingworth, P.J., N.P. Groome, W.C. Duncan, V. Grant, S. Tovanabutra, D.T. Baird and A.S. McNeilly, 1996. Measurement of circulating inhibin forms during the establishment of pregnancy. J. Clin. Endocrinol. Metab., 81: 1471-1475.
CrossRef - Jin, W., S. Wada, K.Y. Arai, H. Kishi and C.B. Herath et al., 2001. Testicular secretion of inhibin in the male golden hamster (Mesocricetus auratus). J. Androl., 22: 207-211.
Direct Link - Kalaba, Z.M. and A.E. Abdel-Khalek, 2011. Effect of repeated GnRH injection and intra-vaginal mechanical stimulation on reproductive performance of doe rabbits. Asian J. Anim. Vet. Adv., 6: 1010-10108.
CrossRefDirect Link - Kaneko, H., Y. Nakanishi, S. Akagi, K. Arai and K. Taya et al., 1995. Immunoneutralization of inhibin and estradiol during the follicular phase of the estrous cycle in cows. Biol. Reprod., 53: 931-939.
PubMedDirect Link - Kaneko, H., J. Noguchi, K. Kikuchi, J. Todoroki and Y. Hasegawa, 2002. Alterations in peripheral concentrations of inhibin A in cattle studied using a time-resolved immunofluorometric assay: Relationship with estradiol and folliclestimulating hormone in various reproductive conditions. Biol. Reprod., 67: 38-45.
CrossRef - Kingsley, D.M., 1994. The TGF-beta superfamily: New members, new receptors and new genetic tests of function in different organisms. Genes. Dev., 8: 133-146.
CrossRefDirect Link - Knight, P.G. and C. Glister, 2001. Potential local regulatory functions of inhibins, activins and follistatin in the ovary. Reproduction, 121: 503-512.
PubMedDirect Link - Knight, P.G., A.J. Beard, J.H.M. Wrathall and R.J. Castillo, 1989. Evidence that the bovine ovary secretes large amounts of inhibin a-subunit and its isolation from bovine follicular fluid. J. Mol. Endocrinol., 2: 189-200.
CrossRef - Kondo, M., C. Kojima, G. Watanabe, K. Shimizu, M. Itoh, T. Udono and K. Taya, 2003. Endocrinologic comparison of activin A secretion during pregnancy and early lactation in Japanese monkeys, chimpanzees and humans. Endocrine, 22: 239-243.
CrossRef - Krummen, L.A., A. Moore, T.K. Woodruff, R. Covello, P. Taylor, P. Working and J.P. Mather, 1994. Localization of inhibin and activin binding sites in the testis during development by in situ ligand binding. Biol. Reprod., 50: 734-744.
Direct Link - Lambert-Messerlian, G.M., S. Luisi, P. Florio, V. Mazza, J.A. Canick and F. Petraglia, 1998. Second trimester levels of maternal serum total activin A and placental inhibin/activin α and βA subunit messenger ribonucleic acids in down syndrome pregnancy. Eur. J. Endocrinol., 138: 425-429.
Direct Link - Lincoln, G.A., J. Townsend and H.N. Jabbour, 2001. Prolactin actions in the sheep testis: A test of the priming hypothesis. Biol. Reprod., 65: 936-943.
Direct Link - Lockwood, G.M., S. Muttukrishna and W.L. Ledger, 1998. Inhibins and activins in human ovulation, conception and pregnancy. Hum. Reprod. Update, 4: 284-295.
CrossRefDirect Link - Mann, G.E., B.K. Campbell, A.S. McNeilly and D.T. Baird, 1993. Follicular development and ovarian hormone secretion following passive immunization of ewes against inhibin or oestradiol. J. Endocrinol., 136: 225-233.
CrossRef - Manuelpillai, U., M. Schneider-Kolsky, A. Dole and E.M. Wallace, 2001. Activin A and activin receptors in gestational tissue from preeclamptic pregnancies. J. Endocrinol., 171: 57-64.
CrossRef - Martin, T.L., G.L.Williams, D.D. Lunstra and J.J. Ireland, 1991. Immunoneutralization of inhibin modifies hormone secretion and sperm production in bulls. Biol. Reprod., 45: 73-77.
Direct Link - Mather, J.P., T.K. Woodruff and L.A. Krummen, 1992. Paracrine regulation of reproductive function by inhibin and activin. Proc. Soc. Exp. Biol. Med., 201: 1-15.
PubMed - McNeilly, A.S., 1984. Changes in FSH and the pulsatile secretion of LH during the delay in oestrus induced by treatment of ewes with bovine follicular fluid. J. Reprod. Fertil., 72: 165-172.
Direct Link - McNeilly, A.S., W. Crow and B.K. Campbell, 1991. Effect of follicular fluid and inhibin immunoneutralization on FSH-induced pre-ovulatory follicle growth in the ewe. J. Endocrinol., 131: 401-409.
CrossRef - Medan, M.S., G. Watanabe, K. Sasaki, N.P. Groome, S. Sharawy and K. Taya, 2005. Follicular and hormonal dynamics during the estrous cycle in goats. J. Reprod. Dev., 51: 455-463.
Direct Link - Medan, M.S., G. Watanabe, K. Sasaki, Y. Nagura and H. Sakaime et al., 2003. Ovarian and hormonal response of female goats to active immunization against inhibin. J. Endocrinol., 177: 287-294.
CrossRef - Medan, M.S., G. Watanabe, K. Sasaki, S. Sharawy, N.P. Groome and K. Taya, 2003. Ovarian dynamics and their associations with peripheral concentrations of gonadotropins, ovarian steroids, and inhibin during the estrous cycle in goats. Biol. Reprod., 69: 57-63.
PubMed - Medan, M.S., H. Wang, G. Watanabe, A.K. Suzuki and K. Taya, 2004. Immunization against endogenous inhibin increases normal oocyte/embryo production in adult mice. Endocrine, 24: 115-119.
CrossRef - Medan, M.S., G. Watanabe, Y. Nagura, H. Kanazawa, M. Fujita and K. Taya, 2004. Passive immunoneutralization of endogenous inhibin increases ovulation rate in miniature Shiba goats. J. Reprod. Dev., 50: 705-710.
Direct Link - Medan, M.S., K.Y. Arai, G. Watanabe and K. Taya, 2007. Inhibin: Regulation of reproductive function and. practical use in females. Anim. Sci. J., 78: 16-27.
CrossRef - Meliani, S., B. Benallou, S.A. Abdelhadi, M. Halbouche and A. Naceri, 2011. Environmental factors affecting gestation duration and time of foaling of pure bred arabian mares in algeria. Asian J. Anim. Vet. Adv., 6: 599-608.
CrossRef - Miyamoto, K., Y. Hasegawa, M. Fukuda, M. Nomura, M. Igarashi, K. Kangawa and H. Matsuo, 1985. Isolation of porcine follicular fluid inhibin of 32K Daltons. Biochem. Biophys. Res. Commun., 129: 396-400.
CrossRef - Moeini, M.M., F. Alipour and A. Moghadam, 2009. The effect of human chorionic gonadotropin on the reproduction performance in lory sheep synchronized with different doses of pregnant mare serum gonadotrophin outside the breeding season. Asian J. Anim. Vet. Adv., 4: 9-15.
CrossRefDirect Link - Muttukrishna, S., T.J. Child, N.P. Groome and W.L. Ledger, 1997. Source of circulating levels of inhibinA, pro a C-containing inhibins and activin A in early pregnancy. Hum. Reprod., 12: 1089-1093.
Direct Link - Nambo, Y., H. Kaneko, S. Nagata, M. Oikawa and T. Yoshihara et al., 1998. Effect of passive immunization against inhibin on FSH secretion, folliculogenesis and ovulation rate during the follicular phase of the estrous cycle in mares. Theriogenology, 50: 545-557.
PubMed - Nambo, Y., K. Nagaoka, Y. Tanaka, N. Nagamine and H. Shinbo et al., 2002. Mechanisms responsible for increase in circulating inhibin levels at the time of ovulation in mares. Theriogenology, 57: 1707-1717.
CrossRef - O'Shea, T., M.A. Hillard, S.T. Anderson, B.M. Bindon, J.K. Findlay, C.G. Tsonis and J.F. Wilkins, 1994. lnhibin immunization for increasing ovulation rate and superovulation. Theriogenology, 41: 317-322.
CrossRef - Ohshima, K., K. Ohshima, K.Y. Arai, H. Kishi and M. Itoh et al., 2002. Potential role of activin A in follicular development during the second half of pregnancy in the golden hamster: Utero-placental source of activin A. J. Endocrinol., 172: 247-253.
Direct Link - Ohshima, K., H. Kishi, M. Itoh, G. Watanabe and K. Arai et al., 1999. Secretion of inhibinA, inhibin B and inhibin pro-aC during the oestrous cycle of the golden hamster (Mesocricetus auratus). J. Endocrinol., 162: 451-456.
Direct Link - Pangas, S.A. and T.K. Woodruff, 2002. Production and purification of recombinant human inhibin and activin. J. Endocrinol., 172: 199-210.
Direct Link - Petraglia, F., G.C. Garuti, L. Calza, V. Roberts and L. Giardino et al., 1991. Inhibin subunits in human placenta: Localization and messenger ribonucleic acid levels during pregnancy. Am. J. Obstet. Gynecol., 165: 750-758.
Direct Link - Price, C.A., B.A. Morris, T. O'Shea and R. Webb, 1987. Active immunization of cattle against partly purified follicular fluid from sheep. J. Reprod. Fertil., 81: 161-168.
CrossRef - Ramaswamy, S. and T.M. Plant, 2001. Operation of the Follicle-Stimulating Hormone (FSH)-inhibin B feedback loop in the control of primate spermatogenesis. Mol. Cell Endocrinol., 180: 93-101.
CrossRef - Razie, S., M. Panahi, A. Ahangarpor, F. Rahim and G. Saki, 2011. Study of level of inhibin B and ultra structure of sertoli cells in contra-lateral testis after unilateral blunt testis trauma in rat. Asian J. Biol. Sci., 4: 70-76.
CrossRefDirect Link - Rivier, C. and W. Vale, 1989. Immunization of endogenous inhibin modifies hormone secretion and ovulation rate in the rat. Endocrinology, 125: 152-157.
CrossRef - Rizwan, S., A. Iftikhar, A. Muhammad, A. Shagufta, Y. Tahira and S. Adeela, 2005. Advance effect of pesticides on reproduction hormones of women cotton pickers. Pak. J. Biol. Sci., 8: 1588-1591.
CrossRefDirect Link - Robertson, D.M., L.M. Foulds, R.C. Fry, J.T. Cummins and I. Clarke, 1991. Circulating half-lives of follicle-stimulating hormone and luteinizing hormone in pituitary extracts and isoform fractions of ovariectomized and intact ewes. Endocrinology, 129: 1805-1813.
Direct Link - Sanford, L.M., C. Moore, J.K. Voglmayr and M.H. Fahmy, 2000. Sexualmaturational changes in circulatory inhibin concentration in relation to FSH concentration and testicular size in Suffolk and DLS rams. Theriogenology, 54: 719-730.
CrossRef - Sanford, L.M., J.K. Voglmayr, W.W. Vale and B. Robaire, 1993. Photoperiodmediated increases in serum concentrations of inhibin, folliclestimulating hormone, and luteinizing hormone are accentuated in adult shortened-scrotum rams without corresponding decreases in testosterone and estradiol. Biol. Reprod., 49: 365-373.
Direct Link - Sasaki, K., M.S. Medan, G. Watanabe, S. Sharawy and K. Taya, 2006. Immunization of goats against inhibin increased follicular development and ovulation rate. J. Reprod. Dev., 52: 543-550.
Direct Link - Satterlee, D.G., S.A. Castille and W.C. Fioretti, 2006. Active immunization of broiler breeder cockerels against chicken inhibin accelerates puberty and prevents age-induced testicular involution. Poult. Sci., 85: 1087-1094.
Direct Link - Schanbacher, B.D., 1991. Pituitary and testicular responses of beef buls to active immunization against inhibin alpha. J. Anim. Sci., 69: 252-257.
Direct Link - Schneyer, A.L., T. Fujiwara, J. Fox, C.K. Welt, J. Adams, G.M. Messerlian and A.E. Taylor, 2000. Dynamic changes in the intrafollicular inhibin/activin/follistatin axis during human follicular development: Relationship to circulating hormone concentrations. J. Clin. Endocrinol. Metab., 85: 3319-3330.
- Sejian, V., T.V. Meenambigai, M. Chandirasegaran and S.M.K. Naqvi, 2010. Reproductive technology in farm animals: New facets and findings: A review. J. Biol. Sci., 10: 686-700.
CrossRefDirect Link - Sewani, C.R., M.M. Bagdasarian, J.J. Ireland and M. Bagdasarian, 1998. Display of an inhibin epitope in a surface-exposed loop of the E.coli heat labile enterotoxin B subunit. Vaccine, 16: 1611-1619.
CrossRef - Shi, F., K. Mochida, O. Suzuki, J. Matsuda and A. Ogura et al., 2000. Development of embryos in superovulated guinea pigs following active immunization against the inhibin a-subunit. Endocr. J., 47: 451-459.
Direct Link - Sowers, M.R., A.D. Eyvazzadeh, D. McConnell, M. Yosef and M.L. Jannausch et al., 2008. Anti-mullerian hormone and inhibin B in the definition of ovarian aging and the menopausetransition. J. Clin. Endocrinol. Metab., 93: 3478-3483.
CrossRef - Sugino, K., T. Nakamura, K. Takio, K. Miyamoto and Y. Hasegawa et al., 1992. Purification and characterization of high molecular weight forms of inhibin from bovine follicular fluid. Endocrinology, 130: 789-796.
PubMed - Takedomi, T., H. Kishi, M.S. Medan, Y. Aoyagi and M. Konishi et al., 2005. Active immunization against inhibin improves superovulatory response to exogenous FSH in cattle. J. Reprod. Dev., 51: 341-346.
PubMed - Takedomi, T., H. Kaneko, Y. Aoyagi, K. Konishi, H. Kishi, G. Watanabe and K. Taya, 1997. Effects of passive immunization against inhibin on ovulation rate and embryo recovery in Holstein heifers. Theriogenology, 47: 1507-1518.
Direct Link - Tannetta, D.S., S.A. Feist, E.C. Bleach, N.P. Groome, L.W. Evans and P.G. Knight, 1998. Effects of active immunization of sheep against an amino terminal peptide of the inhibin a C subunit on intrafollicular levels of activin A, inhibin A and follistatin. J. Endocrinol., 157: 157-168.
PubMed - Terqui, M., J.H.M. Wrathall, M.A. Driancourt and P.G. Knight, 1995. Modulation of ovarian function by steroid and inhibin immunization. Livest. Prod. Sci., 42: 181-192.
Direct Link - Tilbrook, A.J., D.M. de Kretser and I.J. Clarke, 1999. Changes in the suppressive effects of recombinant inhibinA on FSH secretion in ram lambs during sexual maturation: Evidence for alterations in the clearance rate of inhibin. J. Endocrinol., 161: 219-229.
Direct Link - Vale, W., C. Rivier, A. Hsueh, C. Campen and H. Meunier et al., 1988. Chemical and biological characterization of the inhibin family of protein hormones. Rec. Prog. Horm Res., 44: 1-34.
Direct Link - Meenakshi, V., A.K. Gupta and S.K. Garg, 2006. Determination of gestation period of mare with maximum eCG concentration by bioassay. Indian J. Anim. Res., 40: 25-29.
Direct Link - Meenakshi, V., A.K. Gupta, V. Nitin and S.K. Garg, 2008. Haemagglutination inhibition assay for equine chorionic gonadotropin (eCG) detection in pregnant mare serum. J. Immunol. Immunopathol., 10: 55-59.
Direct Link - Voge, J.L. and J.E. Wheaton, 2007. Effects of immunization against two inhibin antigens on daily sperm production and hormone concentrations in ram lambs. J. Anim. Sci., 85: 3249-3255.
CrossRefDirect Link - Voglmayr, J.K., M. Mizumachi, D.W. Washington, C.C. Chen and W. Bardin, 1990. Immunization of rams against human recombinantinhibin α-subunit delays, augments and extends season-related increase in blood gonadotropin levels. Biol. Reprod., 42: 81-86.
Direct Link - Voglmayr, J.K., D. Jolley, W. Vale, D. Willoughby and A. Moser et al., 1992. Effects of follicle-stimulating hormone on inhibin release by different testicular compartments in the adult ram. Biol. Reprod., 47: 573-581.
Direct Link - Wang, H., C.B. Herath, G. Xia, G. Watanabe and K. Taya, 2001. Superovulation, fertilization and in vitro embryo development in mice after administration of an inhibinneutralizing antiserum. Reproduction, 122: 809-816.
PubMed - Welt, C., K.A. Martin, A. Taylor, G. Lambert-Messerlian and W.F. Crowley Jr. et al., 1997. Frequency modulation of follicle-stimulating hormone (FSH) during the luteal-follicular transition: Evidence for FSH control of inhibin-B in normal women. J. Clin. Endocrinol. Metabol., 82: 2645-2652.
Direct Link - Welt, C.K. and A.L. Schneyer, 2001. Differential regulation of inhibin B and inhibin A by follicle-stimulating hormone and local growth factors in human granulosa cells from small antral follicles. J. Clin. Endocrinol. Metabol., 86: 330-336.
Direct Link - Welt, C.K., J.M. Adams, P.M. Sluss and J.E. Hall, 1999. Inhibin A and inhibin-B responses to gonadotropin withdrawal depends on stage of follicle development. J. Clin. Endocrinol. Metabol., 84: 2163-2169.
CrossRefDirect Link - Welt, C.K., Z.A. Smith, D.K. Pauler and J.E. Hall, 2001. Differential regulation of inhibin A and inhibin B by luteinizing hormone, follicle-stimulating hormone, and stage of follicle development. J. Clin. Endocrinol. Metabol., 86: 2531-2537.
Direct Link - Wheaton, J.E. and R.W. Godfrey, 2003. Plasma LH, FSH, testosterone, and age at puberty in ram lambs actively immunized against an inhibin α-subunit peptide. Theriogenology, 60: 933-941.
Direct Link - Woodruff, T.K., L.M. Besecke, N. Groome, L.B. Draper, N.B. Schwartz and J. Weiss, 1996. Inhibin A and inhibin B are inversely correlated to follicle-stimulating hormone, yet are discordant during the follicular phase of the rat estrous cycle and inhibin A is expressed in a sexually dimorphic manner. Endocrinology, 137: 5463-5467.
Direct Link - Wrathall, J.H.M., B.J. McLeod, R.G. Glencross, A.J. Beard and P.G. Knight, 1990. lnhibin immunoneutralization by antibodies raised against synthetic peptide sequences of inhibin a subunit: Effects on gonadotrophin concentrations and ovulation rate in sheep. J. Endocrinol., 124: 167-176.
CrossRef - Gharbi, I., M. Ferrouk, A. Dechicha, G. Baril, J.F. Beckers and D. Guetarni, 2012. Follicular status and embryo production in ouled djellal (Algeria) ewes breed pretreated with a GnRH agonist. Asian J. Anim. Vet. Adv., 70: 117-127.
CrossRefDirect Link