
H.H. Azzaz
Department of Dairy Science, National Research Center, Dokki, Giza, Egypt
A.M. Kholif
Department of Dairy Science, National Research Center, Dokki, Giza, Egypt
H.A. Murad
Department of Dairy Science, National Research Center, Dokki, Giza, Egypt
M.A. Hanfy
Department of Animal Production, Faculty of Agriculture, Cairo University, Giza, Egypt
M.H. Abdel Gawad
Department of Animal Production, Faculty of Agriculture, Cairo University, Giza, Egypt
Asian Journal of Animal and Veterinary Advances
Year: 2012 | Volume: 7 | Issue: 8 | Page No.: 664-673







ABSTRACT
An in vitro study was conducted to evaluate the effect of cellulases addition to banana wastes on dry matter (IVDMD) and organic matter (IVOMD) disappearances. Laboratory produced cellulase (Asperozym) and a commercial cellulolytic enzyme source (Bacillozym®) were added separately to banana wastes at 4 levels (0, 0.77, 1.54, 2.31 and 3.08 Unit/kg DM). Increasing the Asperozym levels up to 3.08 U kg-1 DM exhibited the highest (p<0.05) IVDMD and IVOMD, while Bacillozym® recorded the highest (p<0.05) IVDMD and IVOMD values at 1.54 U kg-1 DM compared with the untreated banana wastes (Control). Nine lactating Zaraibi goats (about 3 years old and weight on average 31±0.2 kg) after parturition were divided into three groups of three animals each, using 3x3 Latin square designs to evaluate the effect of Asperozym and Bacillozym® addition to diets on the productivity of lactating goats. Animals were fed on 50% Concentrate Feed Mixture (CFM), 25% banana wastes and 25% berseem (clover) straw (control diet). Control diet+ Asperozym at level of 3.08 U kg-1 DM (T1); control diet+Bacillozym® at level of 1.54 U kg-1 DM. (T2). Apparent digestibility for all nutrients were improved (p<0.05) by cellulases treatments. Milk and 4% Fat Corrected Milk (FCM) yields were higher (p<0.05) for T1 group followed by T2 group than control group while milk composition was not affected (p<0.05). Blood plasma Aspartate aminotransferase (AST) and glucose concentration were not affected by treatments. The addition of Asperozym and Bacillozym® to diets improved the performances of lactating Zaraibi goats with no deleterious effects on general health.
PDF Abstract XML References Citation
Received: November 04, 2011;
Accepted: February 07, 2012;
Published: May 10, 2012
How to cite this article
H.H. Azzaz, A.M. Kholif, H.A. Murad, M.A. Hanfy and M.H. Abdel Gawad, 2012. Utilization of Cellulolytic Enzymes to Improve the Nutritive Value of Banana Wastes and Performance of Lactating Goats. Asian Journal of Animal and Veterinary Advances, 7: 664-673.
DOI: 10.3923/ajava.2012.664.673
URL: https://scialert.net/abstract/?doi=ajava.2012.664.673
DOI: 10.3923/ajava.2012.664.673
URL: https://scialert.net/abstract/?doi=ajava.2012.664.673
INTRODUCTION
In some small ruminant production systems, roughages constitute the major portion of all available feed resources (Kabir et al., 2002; Hossain et al., 2004). Because of continuously elevating feed prices, attempts to use new sources of roughages such as banana wastes had been evaluated by several workers (Khattab et al., 2000; El-Ashry et al., 2003; Amarnath and Balakrishnan, 2007).
Each hectare of banana crop generates 13 to 20 tones dry matter/year of plant residual waste (leaves and pseudostems) that consists mainly of lignocellulose material (Amarnath and Balakrishnan, 2007). The main shortcoming of agricultural by-products like banana wastes as a sole ruminant feed lies in their low protein and high crude fiber content, low digestibility coefficients and containing some anti-nutrients factors such as tannins and alkaloids (Kholif et al., 2005; Aritonang, 2009). Thus, to increase digestibility of banana wastes, it is important to degrade their compact lignocellulytic tissue. There have been attempts to achieve such objective by biological treatments which could be conducted by administration of the microbial cells, microbial extracts or microbial enzymes (Ghorbani et al., 2007; Khadem et al., 2007; Murad and Azzaz, 2010; Akinyele et al., 2011; Murad and Azzaz, 2011).
Recent research has demonstrated that supplementing diets of ruminants with cellulase can improve feed utilization and animal performance by enhancing fiber degradation in vitro (El-Adawy et al., 2008; Rodrigues et al., 2008), in situ (Tricarico et al., 2005; Krueger and Adesogan, 2008) and in vivo (Gado et al., 2007; Gado et al., 2009; Khattab et al., 2011). Also, milk production was increased by adding cellulolytic enzyme preparations to lactating small ruminant’s diets (Titi and Lubbadeh, 2004; Stella et al., 2007).
Cellulase with its immense importance is being imported for use in Egypt at a high cost. The local production of such enzymes may reduce the cost of importation and encourages self-reliance.
This study was conducted to, (1) Evaluate potential use of the laboratory produced cellulase in vitro for degradation of banana waste compared with commercial cellulolytic enzyme source, (2) Study effects of adding these cellulolytic enzymes to lactating goat's diets on nutrients digestibility, blood parameters, as well as on milk yield and its composition.
MATERIALS AND METHODS
This study was carried out at the Experimental Farm Station of the Faculty of Agriculture, Cairo University and Dairy Department, National Research Center, Dokki, Giza, Egypt.
Collecting banana wastes: Green banana wastes (leaves and pseudostems) were collected from banana farms after harvesting at Om Dinar, Embaba, Giza province. The wastes were cut and sun-dried, then chopped to 0.5-1 cm and stored in dry place at room temperature until used.
Enzyme sources: Bacillozym®; Product of IBEX International is a commercial cellulolytic enzyme source produced from Bacillus subtilis. Each kg contains 15000 commercial units of cellulase, Bacillus subtilis 0.75x1010 (CFU).
Asperozym; Laboratory produced cellulase from Asperigillus niger. Each kg contains 770 international units (IU) of cellulase. One unit of Asperozym is equivalent to 19.48 unit of Bacillozym®.
Enzymes assay: The carboxymethyl-cellulase activity (CMC) for Bacillozym® and Asperozym was determined according to Mandels et al. (1974). The reducing sugar liberated was determined by modified Dinitrosalicylic acid method (DNS) of Miller (1972). One cellulase unit is defined as the amount of enzyme that liberates reducing sugar at the rate of one μmol/mL/min under assay condition.
In vitro study: In vitro dry matter and organic matter disappearance (IVDMD and IVOMD) was determined for banana waste powder. A 500 mg samples of banana waste powder were weighed into 120 mL serum bottles. The experimental banana waste (five replicates) was separately supplemented with rumen liquor, buffer solution and Asperozym and Bacillozym solutions at different levels (0, 0.77, 1.54, 2.31 and 3.08 U kg-1 DM). Rumen contents were collected by stomach tube from rams fed berseem hay ration before the morning feeding, then moved directly to the laboratory in separate warmed oxygen-free plastic jars. Rumen liquor contents were strained through two layers of cheese-cloth and the obtained liquor was mixed with the buffer solution at 39°C under continuous flushing with CO2 using two stage technique according to method of Norris et al. (1976). Bottles were sealed with rubber stoppers and incubated at 39°C for 48 h.
Lactation trial: According to results of in vitro trial, the proper dry matter and organic matter disappearances of different levels of Asperozym and Bacillozym® addition, Asperozym at 3.08 U kg-1 DM and Bacillozym® at 1.54 U kg-1 DM were chosen to be used in the lactation trial.
Feeding and management: Nine Zaraibi lactating goats (about 3 years old and weighting on average 31±0.2 kg) after parturition were assigned randomly into three groups of three animals each using 3x3 Latin square design. The experimental periods were 12 weeks (84 days) and consisted of three equal periods (28 day each). The goats were individually fed at 3% of body weight changed continuously according to animal weight changes. The concentrate:roughage ratio was 1:1 on DM basis. The concentrate feed mixture consisted of 60% corn, 20% soybean meal, 15% wheat bran, 3% limestone, 1% minerals and 1% NaCl. The first group was fed on 50% concentrate Feed Mixture (CFM), 25% berseem (clover) straw and 25% banana waste (control diet). The second group was fed control diet+Asperozym at 3.08 U kg-1 DM (T1). The third group was fed control diet+Bacillozym® at 1.54 U kg-1 DM (T2). The concentrate feed mixture was offered once daily at 8.00 a.m., berseem straw and banana waste were offered once daily at 9.00 a.m. The enzymes were introduced to each animal as a capsule before roughage feeding. The control group was getting a capsule free from any enzyme. The chemical composition of feed ingredients is shown in Table 1.
Apparent digestibility: Three digestibility trials were applied during the last seven days every each experimental period (28 day) using all animals from each group. Silica was used as an internal marker for determining the digestibility (Ferret et al., 1999). Four hours after the distribution of morning meal (09:00 h) feces were collected in cloth bag connected to the animal back. The collected feces were dried at 55°C for 48 h and then ground to pass a 1 mm sieve in a feed mill (FZ102, Shanghai Hong Ji instrument Co., Ltd., Shanghai, China) for chemical analysis. Dry matter excreted in feces was calculated by dividing silica input in the feeds (grams of silica per day) by silica output in the feces (grams of silica per day).
| Table 1: | Chemical analysis of feed ingredients (on DM basis) |
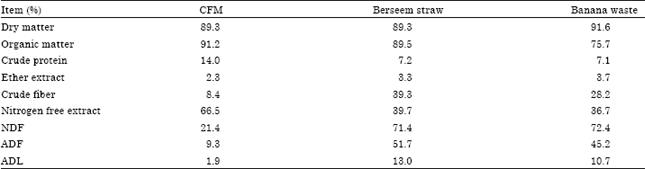 | |
| NDF: Neutral detergent fiber, ADF: Acid detergent fiber, ADL: Acid detergent lignin, CFM: Concentrate feed mixture consisted of 60% corn, 20% soybean meal, 15% wheat bran, 3% ground limestone, 1% NaCl and 1% Mineral and vitamin mix contained 42 ppm Co, 3500 ppm Cu, 20,000 ppm Fe, 12,000 ppm Mn, 12, 00 ppm Zn, 1200 ppm I, 3800 IU g-1 of vitamin A, 1200 IU g-1 of vitamin D and 3 IU g-1 of vitamin E | |
The digestibility coefficient of nutrient was calculated according to the following formula (Ferret et al., 1999):
Feed and fecal analysis: Feedstuffs and fecal samples were analyzed according to the AOAC (1995) methods to determine Crude Protein (CP), Ether Extract (EE), Crude Fiber (CF) and ash contents. Organic Matter (OM) and Nitrogen Free Extract (NFE) contents were calculated by difference. The Neutral Detergent Fiber (NDF) and Acid Detergent Fiber (ADF) contents were determined using the methods described by Van Soest et al. (1991).
Blood plasma analysis: Blood samples were collected from the jugular vein of each animal at the last day of each period (4 h after the 09:00 h feeding). They were centrifuged at 4000 r.p.m./ 20 min. The plasma was stored at -18°C till analysis. Plasma was collected and plasma total protein was determined as described by Armstrong and Carr (1964), albumin (Doumas et al., 1971), urea (Fawcett and Soctt, 1960), glucose (Siest et al., 1981) and plasma Aspartate aminotransferase (AST) and Alanin aminotransferase (ALT) (Reitman and Frankel, 1957). Globulin and albumin/globulin ratio were calculated.
Sampling and analysis of milk: Animals were milked twice daily at 8.00 a.m. and 4.00 p.m. during the last three days of each experimental period (28 day). Samples of milk were collected immediately from each animal after morning and evening milking and milk yield was recorded. Milk samples were analyzed for total solids, fat, true protein and lactose by infrared spectrophotometry (Foss 120 Milko-Scan, Foss Q3 183 Electric, Hillerød, Denmark) according to AOAC (1995) procedures. Solids-not-fat (SNF) was calculated. Fat corrected milk (4% fat) was calculated by using the following equation according to Gaines (1928):
FCM = 0.4 M+15 F |
where, M is milk yield (g) and F is fat yield (g).
Statistical analysis: Data obtained from this study were statistically analyzed by SAS (1998) as follow:
Latin square design for milk yield and composition, nutrients digestibility and blood parameters using the general linear model procedure:
Yijk = μ+Ri+Cj+Tk+eijk |
where, Yijkl is the parameter under analysis of the ijk goat, μ is the overall mean, Ri is the effect due to the lactation period on the parameter under analysis, Cj is the effect due to the animals on the parameter under analysis, Tk is the effect due to treatment on the parameter under analysis, eijk is the experimental error for ijk on the observation. The Duncan's multiple range tests was used to test the significance between means (Duncan, 1955).
RESULTS AND DISCUSSION
In vitro study: All levels of Asperozym and Bacillozym® increased (p<0.05) DM and OM disappearance of banana waste compared with the untreated banana waste (Control), which gave the lowest values of IVDMD and IVOMD (Table 2). This may be due to that Asperozym and Bacillozym® were able to degrade complex substrate (cellulose) to simpler ones which might have altered the structure of banana wastes making them more amenable to ruminal microorganisms and allowing a faster ruminal microbial colonization and fermentation. Till now, little is known about the way that exogenous fibrolytic enzymes improve feed by rumen microorganisms. Several potential modes of action have been proposed. These include: (a) increase in microbial colonization of feed particles (Yang et al., 1999) enhancing attachment and/or improve access to the cell wall matrix by ruminal microorganisms and by doing so, accelerate the rate of digestion (Nsereko et al., 2000b) and (c) enhancing the hydrolytic ability of the ruminal microorganisms due to added enzyme activities and/or synergy with rumen microbial enzymes (Morgavi et al., 2000). On the other hand the IVDMD and IVOMD were highest with the highest level of Asperozym (3.08 U kg-1 DM) and Bacillozym® (1.54 U kg-1 DM). This may be related to some different biochemical properties of the experimental enzymes such as source organism, molecular size, etc. (Vahjen and Simon, 1999). Also, Eun and Beauchemin (2007) suggested that the relationship between enzymatic activity and substrate degradation may depend on the amount of enzymatic activity added or may be due to the different kinds of enzymes used.
Digestibility and nutritive values: Diets treated with Asperozym (T1) and Bacillozym® (T2) increased (p<0.05) all nutrients digestibility and fiber fraction digestibility compared with the control diet (Table 3). The goats fed (T1) diet showed increase (p<0.05) most of nutrients digestibility than those fed (T2) diet, also, the nutritive values of the experimental diets expressed as Total Digestible Nutrients (TDN) and Digestible Crude Protein (DCP) take the same trend of digestibility (Table 3). These data in line with Gado et al. (2009) who reported increase total tract digestibility of DM, OM and NDF, following treatment with fibrolytic enzymes. The higher values of CF, NDF, ADF and ADL digestibility of the diets treated with Asperozym or Bacillozym® compared to the control diet could be attributed to that Asperozym contains combination of fungal cellulases and; Bacillozym® contains combination of bacterial cellulases which solubilize fibers and subsequently may provide some essential nutrients or growth factors to rumen microorganisms.
| Table 2: | Effect of cellulases on in vitro dry matter and organic matter disappearance of banana wastes |
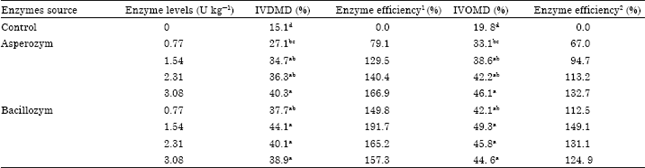 | |
| 1Enzyme efficiency % (DM) = IVDMD% (sample)-IVDMD% (control) / IVDMD% (control) x100, 2 Enzyme efficiency % (OM) = IVOMD% (sample) – IVOMD% (control) / IVOMD% (control) x100. Each value of means obtained from five samples. a, b, c and d with different superscripts in the same column are significantly different (p<0.05) | |
| Table 3: | Effects of cellulases on digestion coefficients and nutritive values of experimental diets fed to goats |
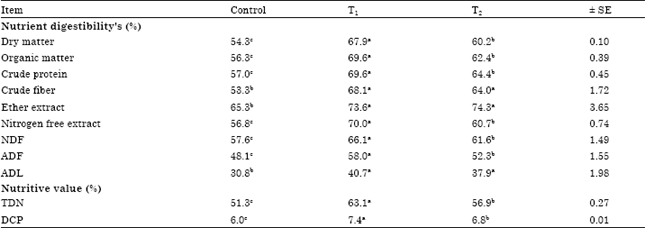 | |
| Each value represents an average of six samples, TDN: Total digestible nutrients, DCP: Digestible crude protein. NDF: Neutral detergent fiber, ADF: Acid detergent fiber, ADL: Acid detergent lignin, Means in the same row within each treatment having different superscripts differ significantly (p<0.05), SE: standard error, T1: control diet+Asperozym at 3.08 U kg-1 DM, T2: control diet+Bacillozym® at 1.54 U kg-1 DM | |
| Table 4: | Effect of cellulases on blood plasma parameters of lactating goats fed with different experimental diets |
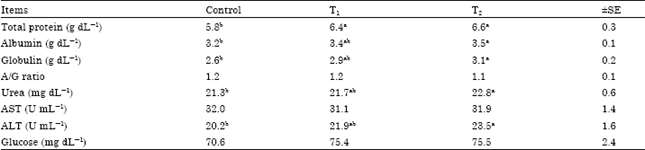 | |
| Each value represents an average of eight samples. a, b, c: Means in the same row within each treatment having different superscripts differ significantly (p<0.05), SE: Standard error, T1: Control diet+Asperozym at 3.08 U kg-1 DM, T2: Control diet+bacillozym® at 1.54 U kg-1 DM, AST: Aspartate amino transferase, ALT: Alanin amino transferase | |
Superiority of Asperozym over Bacillozym® for improving apparent digestibility of treated diets may be due to differences of cellulase type used and enzyme stability (Nsereko et al., 2000a, b) reported that variation of responses to fibrolytic enzymes supplementation could be attributed to the retention time of different types of fiber in the rumen; exposure time of fiber to the fibrolytic enzymes process, rate of particle size reduction, particle density and rate of digestion.
Blood plasma parameters: Cellulases treated diets (T1 and T2) had higher (p<0.05) plasma total protein than those fed the control diet. This may be attributed to the improvements occurred in metabolic process as a response to the cellulases additives and indicate that these goats cover their protein needs. Our finding are in line with those reported by Gado et al. (2007) who reported that biological treatment (cellulase; rumen liquor and Cellumonas cellulasea) of bagasse increased plasma total protein. In addition, plasma albumin, globulin, urea and alanin aminotransferase (ALT) concentrations were higher (p<0.05) in goats fed (T2) diet than in those fed control diet (Table 4). This may be due to a higher organic matter and crude protein (CP) digestibility for these goats compared with those fed control diet. The variation of plasma urea is in line with that reported by Gado et al. (2007) who stated that biological treatment (cellulase; rumen liquor and Cellumonas cellulasea) of bagasse increased plasma urea concentrations.
| Table 5: | Effects of cellulases on goat's milk yield and composition |
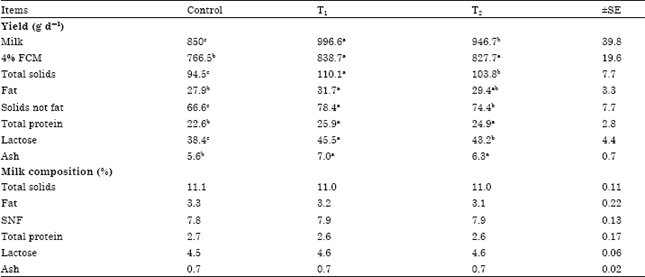 | |
| Each value represents an average of twenty seven samples, Means in the same row within each treatment having different superscripts differ significantly (p<0.05), SE: Standard error, T1: Control diet+asperozym at 3.08 U kg-1 DM, T2: Control diet+Bacillozym® at 1.54 U kg-1 DM, SNF: Soliels-not-fat | |
Blood plasma, aspartate aminotransferase (AST) and glucose concentration were not affected by cellulases treatments. Our results are in line with those obtained by Kholif (2006) who found that animals fed on fibrolytic enzymes or fungi treated silage had no significant increase in serum glucose and AST concentration. The concentrations of ALT and AST were in the normal range for healthy animals. These results indicated that adding cellulases to lactating goat's diets were not negatively affected liver activity or animal’s health.
Milk yield and composition: Milk composition was not affected by cellulases treatments, while milk and 4% Fat Corrected Milk (FCM) yields were higher (p<0.05) for goats fed (T1) and (T2) diets than those fed the control diet. Goats fed (T1) diet produce more milk than those fed (T2) diet (Table 5). Adding Asperozym to lactating goat's diets increased milk production by 18% and fat corrected milk production by 9% , while adding Bacillozym® to lactating goat's diets increased milk production by 11% and fat corrected milk production by 8% compared with untreated diets (control). Our findings are in agreement with the results obtained by Yang et al. (1999), Titi and Lubbadeh (2004) and Gado et al. (2009). This response may be attributed to improved nutrient digestion after cellulases supplementation by goats. Milk fat and protein yields were higher (p<0.05) for goats fed (T1) diet than goats fed control diet, reflecting the higher milk yields but goats fed (T2) diet had higher (p<0.05) milk protein yield but not fat yield compared with goats fed control diet. It is not clear why the fat and protein yields of milk were higher when goats were fed cellulases-treated diets but is likely indirectly related to changes in energy and protein digestion (Beauchemin et al., 1997). The use of enzyme additives has been associated with an improved efficiency of synthesis of microbial protein in the rumen (Jacobs and McAllan, 1992). Therefore, it is probable that improved efficiency of microbial protein synthesis is a result of enzyme action on the roughages structural polysaccharides altering the rate of ruminal degradation of structural carbohydrates (Lewis et al., 1996) and the provision of a suitable ruminally degradable nitrogen source (Beauchemin et al., 1999). However, yield of total solids was significantly increased due to the cumulative effect of cellulases treatment on the fat and protein concentrations as both were numerically higher for the treated groups compared to the control group. Milk lactose yield was higher (p<0.05) for goats fed (T1) and (T2) diets than those fed the control diet, this could be attributed to the generation of more nutrients which become available as a result of improvements in feed digestibility. Specifically, the increase in ruminally fermented OM, which resulted in a numerical downward shift in the ratio of acetate to propionate, would have increased delivery of glucogenic precursors to the mammary gland (Yang et al., 1999).
It could be concluded that Asperozym and Bacillozym® supplementation were more effective for in vitro degradation of banana waste and improve nutrient digestion and milk production by goats fed the experimental diets. Asperozym showed superiority compared with Bacillozym® for improving feed digestion and milk production by Zaraibi goats.
REFERENCES
- Akinyele, B.J., O.O. Olaniyi and D.J. Arotupin, 2011. Bioconversion of selected agricultural wastes and associated enzymes by Volvariella volvacea: An edible mushroom. Res. J. Microbiol., 6: 63-70.
CrossRefDirect Link - Amarnath, R. and V. Balakrishnan, 2007. Evaluation of the banana (Musa paradisiaca) plant by-product`s fermentation characteristics to asses their fodder potential. Int. J. Dairy. Sci., 2: 217-225.
CrossRefDirect Link - Beauchemin, K.A., W.Z. Yang and L.M. Rode, 1999. Effects of grain source and enzyme additive on site and extent of nutrient digestion in dairy cows. J. Dairy Sci., 82: 378-390.
CrossRefDirect Link - Doumas, B.T., W.A. Watson and H.G. Biggs, 1971. Albumin standards and the measurement of serum albumin with bromcresol green. Clin. Chim. Acta, 31: 87-96.
CrossRefPubMedDirect Link - El-Adawy, M.M., A.Z.M. Salem, B.E. Borhami, H.M. Gado, M.S. Khalil and A. Abo-Zeid, 2008. In vitro cecal gas production and dry matter degradability of some browse leaves in presence of enzymes from anaerobic bacterium in NZW rabbit. Proceedings of the 9th WRSA World Rabbit Congress, June 10-13, Verona, Italy, pp: 643-647.
Direct Link - Eun, J.S., and K.A. Beauchemin, 2007. Assessment of the efficacy of varying experimental exogenous fibrolytic enzymes using in vitro fermentation characteristics. Anim. Feed Sci., Technol., 132: 298-315.
CrossRef - Ferret, A., J. Plaixats, G. Caja, J. Gasa and P. Prio, 1999. Using markers to estimate apparent dry matter digestibility, faecal output and dry matter intake in dairy ewes fed Italian ryegrass hay or alfalfa hay. Small Rumin. Res., 33: 145-152.
CrossRefDirect Link - Gado, H.M., A.Z.M. Salem, P.H. Robinson and M. Hassan, 2009. Influence of exogenous enzymes on nutrient digestibility, extent of ruminal fermentation as well as milk production and composition in dairy cows. Anim. Feed Sci. Technol., 154: 36-46.
CrossRefDirect Link - Ghorbani, G.R., A. Jafari, A.H. Samie and A. Nikkhah, 2007. Effects of applying exogenous, non-starch polysaccharidases to pre-weaning starter concentrate on performance of holstein calves. Int. J. Dairy. Sci., 2: 79-84.
CrossRefDirect Link - Hossain, S.M.J., M.R. Alam, N. Sultana, M.R. Amin and M.M. Rashid, 2004. Milk production from indigenous black Bengal goat in Bangladesh. J. Biological Sci., 4: 262-265.
CrossRefDirect Link - Jacobs, J.L. and A.B. McAllan, 1992. Protein supplementation of formic acid- and enzyme-treated silages. 2. Nitrogen and amino acid digestion. Grass Forage Sci., 47: 114-120.
CrossRefDirect Link - Kabir, F., M. Shahjalal, S.A. Chowdhury, J. Alam and M.R. Islam, 2002. Effect of protein supplementation to grazing on growth and reproductive performance in female goats and sheep. Pak. J. Biol. Sci., 5: 719-721.
CrossRefDirect Link - Khadem, A.A., M. Pahlavan, A. Afzalzadeh and M. Rezaeian, 2007. Effects of live yeast Saccharomyces cerevisiae on fermentation parameters and microbial populations of rumen, total tract digestibility of diet nutrients and on the in situ Degradability of alfalfa hay in Iranian Chall sheep. Pak. J. Biol. Sci., 10: 590-597.
CrossRefPubMedDirect Link - Khattab, H.M., A.M. Kholif, H.A. El-Alamy, F.A. Salem and A.A. El-Shewy, 2000. Ensiled banana wastes with molasses or whey for lactating buffaloes during early lactation. Asian-Australasian J. Anim. Sci., 13: 619-624.
CrossRefDirect Link - Khattab, H.M., H.M. Gado, A.E. Kholif, A.M. Mansour and A.M. Kholif, 2011. The potential of feeding goats sun dried rumen contents with or without bacterial inoculums as replacement for berseem clover and the effects on milk production and animal health. Int. J. Dairy Sci., 6: 267-277.
CrossRefDirect Link - Krueger, N.A. and A.T. Adesogan, 2008. Effect of different mixtures of fibrolytic enzymes on the digestion and fermentation of bahiagrass hay. Anim. Feed Sci. Technol., 145: 84-94.
Direct Link - Lewis, G.E., C.W. Hunt, W.K. Sanchez, R. Treacher, G.T. Pritchard and P. Feng, 1996. Effect of direct-fed fibrolytic enzymes on the digestive characteristics of a forage-based diet fed to beef steers. J. Anim. Sci., 74: 3020-3028.
PubMedDirect Link - Mandels, M., L. Hontz and J. Nystrom, 1974. Enzymatic hydrolysis of waste cellulose. Biotechnol. Bioeng., 16: 1471-1493.
CrossRefDirect Link - Morgavi, D.P., K.A. Beauchemin, V.L. Nsereko, L.M. Rode and A.D. Iwaasa et al., 2000. Synergy between ruminal fibrolytic enzymes and enzymes from Trichoderma longibrachiatum. J. Dairy Sci., 83: 1310-1321.
CrossRefDirect Link - Murad, H.A. and H.H. Azzaz, 2010. Cellulase and dairy animal feeding. Biotechnology, 9: 238-256.
CrossRefDirect Link - Murad, H.A. and H.H. Azzaz, 2011. Microbial pectinases and ruminant nutrition. Res. J. Microbiol., 6: 246-269.
CrossRefDirect Link - Nsereko, V.L., D.P. Morgavi, K.A. Beauchemin and L.M. Rode, 2000. Inhibition of ruminant feed enzyme polysaccharidase activities by extracts from silages. Can. J. Anim. Sci., 80: 523-526.
Direct Link - Nsereko, V.L., D.P. Morgavi, L.M. Rode, K.A. Beauchemin and T.A. McAllister, 2000. Effects of fungal enzyme preparations on hydrolysis and subsequent degradation of alfalfa hay fiber by mixed rumen microorganisms in vitro. Anim. Feed Sci. Technol., 88: 153-170.
CrossRefDirect Link - Reitman, S. and S. Frankel, 1957. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol., 28: 56-63.
CrossRefPubMedDirect Link - Rodrigues, M.A.M., P. Pinto, R.M.F. Bezerra, A.A. Dias and C.V.M. Guedes et al., 2008. Effect of enzyme extracts isolated from white-rot fungi on chemical composition and in vitro digestibility of wheat straw. Anim. Feed Sci. Technol., 141: 326-338.
CrossRefDirect Link - Aritonang, S.N., 2009. The effect of forage energy level on production and reproduction performances of Kosta female goat. Pak. J. Nutr., 8: 251-255.
CrossRefDirect Link - Stella, A.V., R. Paratte, L. Valnegri, G. Cigalino and G. Soncini et al., 2007. Effect of administration of live Saccharomyces cerevisiae on milk production, milk composition, blood metabolites and faecal flora in early lactating dairy goats. Small Rumin. Res., 67: 7-13.
CrossRefDirect Link - Titi, H.H. and W.F. Lubbadeh, 2004. Effect of feeding cellulase enzyme on productive responses of pregnant and lactating ewes and goats. Small Rumin. Res., 52: 137-143.
CrossRefDirect Link - Tricarico, J.M., J.D. Johnston, K.A. Dawson, K.C. Hanson, K.R. McLeod and D.L. Harmon, 2005. The effects of an Aspergillus oryzae extract containing alpha-amylase activity on ruminal fermentation and milk production in lactating Holstein cows. Anim. Sci., 81: 365-374.
CrossRefDirect Link - Vahjen, W., and O. Simon, 1999. Biochemical characteristics of non starch polysaccharide hydrolyzing enzyme preparations designed as feed additives for poultry and piglet nutrition. Arch. Anim. Nutr., 52: 1-14.
PubMedDirect Link - van Soest, P.J., J.B. Robertson and B.A. Lewis, 1991. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci., 74: 3583-3597.
CrossRefPubMedDirect Link - Yang, W.Z., K.A. Beauchemin and L.M. Rode, 1999. Effects of an enzyme feed additive on extent of digestion and milk production of lactating dairy cows. J. Dairy Sci., 82: 391-403.
CrossRefPubMedDirect Link - Fawcett, J.K. and J.E. Scott, 1960. A rapid and precise method for the determination of urea. J. Clin. Pathol., 13: 156-159.
CrossRefPubMedDirect Link