
E. Christaki
Laboratory of Nutrition, Faculty of Veterinary Medicine, Aristotle University of Thessaloniki, Greece
M. Karatzia
Clinic of Farm Animals, Faculty of Veterinary Medicine, Aristotle University of Thessaloniki, Greece
E. Bonos
Animal Production, Faculty of Technology of Agronomics, Technological Educational Institute of Western Macedonia, Florina, Greece
P. Florou-Paneri
Laboratory of Nutrition, Faculty of Veterinary Medicine, Aristotle University of Thessaloniki, Greece
C. Karatzias
Clinic of Farm Animals, Faculty of Veterinary Medicine, Aristotle University of Thessaloniki, Greece
Asian Journal of Animal and Veterinary Advances
Year: 2012 | Volume: 7 | Issue: 7 | Page No.: 597-604







ABSTRACT
Nowadays, there is a great interest towards Spirulina-a bluegreen microalga because of its beneficial compounds. The purpose of the present study was to evaluate the effect of dietary Spirulina platensis on milk fatty acid profile of dairy cows. Twenty healthy Holstein crossbreed, average age 4.3 years, in the second month of lactation were allocated in 2 groups. Ten cows in group A served as controls and were fed individually a basal diet, while the other 10 cows in group B consumed the same diet with the daily addition of 40 g Spirulina per cow. The experiment lasted 7 weeks and every 15 days the milk fatty acid profile was evaluated. The results showed that dietary Spirulina decreased saturated fatty acids, whereas it increased monounsaturated and polyunsaturated fatty acids. In conclusion, Spirulina is a promising dietary supplement for the enhancement of cow milk with health-associated unsaturated fatty acids.
PDF Abstract XML References Citation
Received: November 05, 2011;
Accepted: February 01, 2012;
Published: February 25, 2012
How to cite this article
E. Christaki, M. Karatzia, E. Bonos, P. Florou-Paneri and C. Karatzias, 2012. Effect of Dietary Spirulina platensis on Milk Fatty Acid Profile of Dairy Cows. Asian Journal of Animal and Veterinary Advances, 7: 597-604.
DOI: 10.3923/ajava.2012.597.604
URL: https://scialert.net/abstract/?doi=ajava.2012.597.604
DOI: 10.3923/ajava.2012.597.604
URL: https://scialert.net/abstract/?doi=ajava.2012.597.604
INTRODUCTION
Nowadays, consumers’ demands for natural and healthy products are constantly increasing. Therefore, there is great interest in the research for novel functional foods. One strategy for producing such foods is the modification of the animal diets, using bioactive feed supplements, such as macroalgae (Madhusudan et al., 2011) or microalgae (Christaki et al., 2011a; Hoa et al., 2011).
Microalgae, these primitive aquatic organisms are reproduced by simple division once or twice per day and they are characterized as the most productive plants in the world (Marshall, 2007). Among the most known edible microalgae are Spirulina spp. (Arthrospira), belonging to the cyanobacteria, blue-green colored microalgae which are considered as intermediate species between plants and bacteria (Bold and Wynne, 1985; Christaki et al., 2011a). Spirulina is an important source of nutritional compounds of high biological value, known centuries ago by many populations such as China, Greece, Mexico (Mirada et al., 1998; Simpore et al., 2005; Christaki et al., 2010; Nasseri et al., 2011). Spirulina contains:
| • | Proteins about 60-70% (Becker, 2007) and all the essential aminoacids, with excellent bioavailability (Lordan et al., 2011) |
| • | Pigments, being rich in chlorophyll and phycocyanin (Abd El-Baky, 2003; Gouveia et al., 2008) |
| • | Polyunsaturated fatty acids (PUFA), mainly those of the n-3 series, such as docosahexaenoic acid (DHA). Moreover, it is an interesting source of γ-linolenic acid being precursor of prostaglandins, leucotrienes and thromboxanes (Lordan et al., 2011) |
| • | Antioxidants, such as phycocyanin and carotenoids (Gouveia et al., 2008; Christaki et al., 2011b), which can act as provitamin A and can prevent the formation of reactive oxygen species (Lordan et al., 2011), substances that are responsible for chronic diseases like cancer or aging (El-Baz et al., 2002; Rasool et al., 2008) |
| • | Vitamins, containing nearly all the essential, i.e., vitamin A, vitamins of complex B and tocopherols (Spolaore et al., 2006). The peculiar finding for a herbal product is that Spirulina contains vitamin B12, showing the close phylogenic link between these algae and bacteria that produce the above vitamin (Becker, 1994; Abd El-Baky, 2003) |
| • | Minerals like the macrominerals Na, K, Ca, Mg and the microminerals Fe, Zn, Mn and Cu (Becker, 1994) |
Due to the fact that Spirulina has the above biologically active substances, it has been used in animal nutrition, for example rabbits (Colla et al., 2008), pigs (Grinstead et al., 2000), poultry (Carrillo et al., 2008), acquacultures (Gouveia et al., 2008) or ruminants (Kuplys et al., 2009a).
Milk and dairy products can be used as functional foods (Bhat and Bhat, 2011), therefore, nowadays, much attention has been focused on the enrichment of cows milk fat with PUFA, since these are beneficial acids, especially the n-3 series, that cannot be synthesized by humans or animals and can protect against cardiovascular disease, atherosclerosis, skin diseases and arthritis (Simopoulos, 2002; Gouveia et al., 2008; Lee and Hiramatsu, 2011). These fatty acids are absent or at a minimal level in traditional dairy cows rations (Singh and Sachan, 2011a), while they are present in very low proportions of less than 0.1% of total fatty acids in dairy products (Lock and Bauman, 2004).
The present study was designed to examine the effect of dietary Spirulina platensis on the milk production and fatty acid profile of dairy cows.
MATERIALS AND METHODS
Animals: The study was carried out in a commercial farm in northern Greece, in 2010. Twenty healthy cows of Holstein crossbreed were allocated into two groups (A and B) of ten cows each. The cows had an average age of 4.3 years and were on the second month of lactation. They had similar productivity and their average body weight (group A 523.1 kg; group B 526.3 kg) did not differ significantly (p>0.05). The whole experimentation was performed under commercial conditions and lasted 7 weeks.
Diets: Cows in group A served as controls and received a diet without Spirulina, whereas in group B a total of 40 g powdered Spirulina was additionally incorporated daily in the concentrate of each cow. The control diet consisted of corn silage, alfalfa, molasses and concentrate. This diet was formulated to meet the cows’ nutrient requirements in order to balance the milk production at a rate of 1 kg concentrate for 3.3 L of milk. The composition of the basal diet is presented in Table 1. All cows were fed the concentrate individually, twice a day, in the morning and evening, in two equal meals immediately after milking, while they had free access to water.
Measurements in milk samples: Throughout the experiment, milk production was recorded daily. Milk was sampled on the 15, 30 and 45th day of the experimentation to determine the profile of fatty acids.
| Table 1: | Composition of cows’ basal diet |
 | |
Aliquots of milk were collected in duplicate. Before the fatty acid analysis from each cow samples of two consecutive milkings were pooled (i.e., morning and evening).
The fatty acid composition of the milk was determined according to Folch et al. (1957) and AOAC (2005). Separation and quantification of the methyl esters of the fatty acids was carried out with a gas chromatographic system (TraceGC model K07332, ThermoFinnigan, ThermoQuest, Milan, Italy) equipped with a flame ionization detector, a model CSW 1.7 chromatography station (CSW, DataApex Ltd, Prague, Czech republic) and a fused silica capillary column, 30 mx0.25 mm i.d., coated with cyanopropyl polysiloxane (phase type SP-2380) with a film thickness of 0.20 μm (Supelco, Bellefonte, PA, USA). The chromatographic conditions were: Carrier: N2, Flow: 1 mL min-1; Oven: Temperature 70°C for 0.5 min, increase 30°C min-1 to 180°C for 10 min, increase 5°C min-1 to 225°C for 10 min; Inlet temperature: 250°C; Detector temperature: 250°C; Injection: 2 μL, with split 1/40.
Statistical analysis: The experimental data were subjected to statistical analysis with the aid of SPSS 16.0.1 statistical package (SPSS Inc., Chigaco, IL, USA). The general linear model function was used for the Analysis of Variance (ANOVA). A value of p<0.05 was considered significant. The homogeneity of the variances was examined with Levene’s test (Levene, 1960).
RESULTS
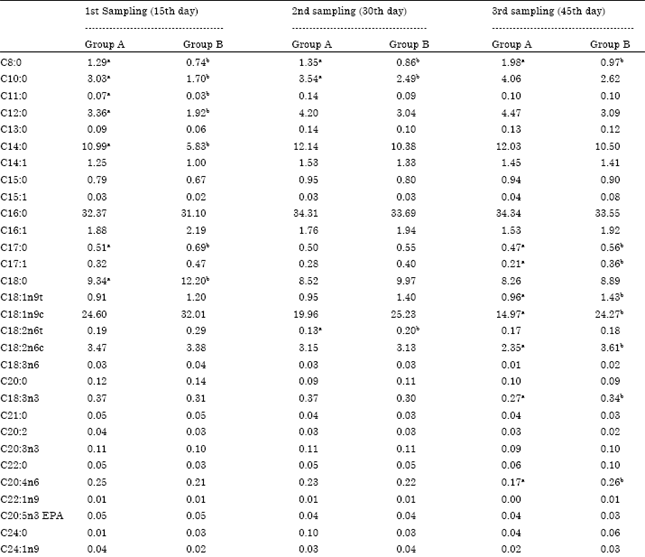
The average milk production did not differ significantly (p>0.05) between the two groups during the whole experiment (Table 2). The dietary addition of Spirulina modified the milk fatty acid composition (Table 3). In the 1st sampling the milk of group B had significantly (p<0.05) lower C8:0, C10:0, C12:0 and C14:0, compared to the milk of group A, whereas C17:0 and C18:0, were significantly (p<0.05) higher. Also, in the second sampling, the milk of group B had significantly (p<0.05) lower C8:0 and C10:0 compared to the milk of group A, whereas C18:2n6t was significantly (p<0.05) higher. Moreover, in the 3rd sampling, the milk of group B had significantly (p<0.05) lower C8:0, while it had significantly higher (p<0.05) C17:0, C17:1, C18:1n9t, C18:1n9c, C18:2n6c, C18:3n3 and C20:4n6.
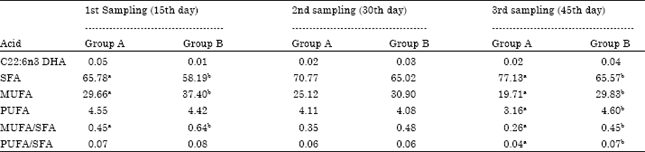
Regarding the total Saturated Fatty Acids (SFA), Monounsaturated Fatty Acids (MUFA) and Polyunsaturated Ratty Acids (PUFA) proportions, it was noticed that Spirulina significantly (p<0.05) decreased SFA in the first and third sampling, significantly increased (p<0.05) MUFA in the first and third sampling and significantly increased (p<0.05) PUFA in the third sampling.
| Table 2: | Milk production of cows receiving either 0 g Spirulina per cow every day (Group A) or 40 g Spirulina per cow every day (Group B) |
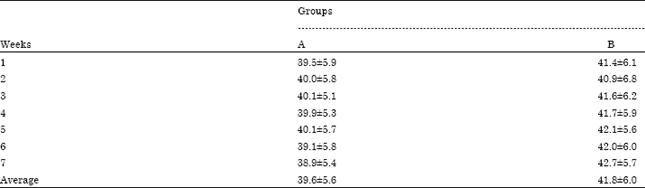 | |
| Values are presented as Means±SD | |
| Table 3: | Milk fatty acid profile (%) of cows receiving either 0 g Spirulina per cow every day (Group A) or 40 g Spirulina per cow every day (Group B) |
 | |
 | |
| Values are presented as Means. Values in the same row with different superscripts differ (p<0.05) | |
Furthermore, the ratio of MUFA/SFA was significantly increased (p<0.05) in the first and third sampling, whereas the ratio of PUFA/SFA was significantly increased (p<0.05) only in the third sampling.
DISCUSSION
Spirulina platensis supplementation to the cows diets had no influence (p>0.05) on milk production during the experimental period. However, literature data have shown that consumption of 200 g day-1 of feed supplement with 5% Spirulina platensis (Kulpys et al., 2009b) or 2 g day-1 per cow biomass of fresh Spirulina platensis (Simkus et al., 2007) or 200 g day-1 dry Spirulina platensis (Kulpys et al., 2009a) significantly increased milk yield.
In present study, the daily incorporation of 40 g Spirulina platensis per cow, in the concentrate, decreased (p<0.05) some of the SFA in the milk fat. Analogous results have been reported by Franklin et al. (1999) and Boeckaert et al. (2008a) who examined the dietary use of microalgae Schizochytrium sp. Moreover, in our experiment Spirulina increased (p<0.05) the MUFA and PUFA, in comparison with the control diet. Boeckaert et al. (2008a) and Singh et al. (2004) reported that the algal meal supplemention (Schizochytrium sp. or Crypthecodinium, respectively) altered the milk unsaturated fatty acid profile.
Among the PUFA in milk fat, Conjugated Linoleic Acid (CLA) was altered significantly. CLA is composed of positional and geometric isomers of linoleic acid (C18:2n) which can be found in food derived from ruminants, exerting inhibitory effects in carcinogenesis (Belury, 2002). In addition, α-linolenic acid (C18:3n3) was increased. This acid can play an important role not only in cardiovascular diseases (Simopoulos, 2002; Singh and Sachan, 2011b) but also in brain development and regeneration of cells in the nervous system, especially in infancy (Christaki et al., 2011c).
When dietary lipids enter the rumen there is a conversion of unsaturated fatty acids to saturated, a process known as biohydrogenation (Lock and Bauman, 2004; Boeckaert et al., 2008b). On the other hand, marine products such as fish oil and algae are rich in PUFA and they can be used as dairy feedstuffs to enhance n-3 fatty acids, since these acids are absent or at minimum levels in traditional dairy cows diets (Lock and Bauman, 2004; Ming et al., 2012). The marine products can change the total bacterial community structure (Kim et al., 2008) and they have been proved to possess a protective mechanism against the rumen biohydrogenation of unsaturated fatty acids (Harfoot and Hazlewood, 1997; Boeckaert et al., 2008b). Algal PUFA remain encapsulated inside the cells of microorganisms and could be protected by the cell membrane (Barclay et al., 1994; Gulati et al., 1999; Papadopoulos et al., 2002).
CONCLUSION
Dietary daily supply of 40 g Spirulina platensis per cow over a period of 7 weeks resulted in decreased (p<0.05) milk SFA. Simultaneously, it increased (p<0.05) milk MUFA and PUFA. Therefore, Spirulina represents one of the most promising sources of functional ingredients and should be further investigated for the enhancement of cow milk with health-associated unsaturated fatty acids.
REFERENCES
- Abd El-Baky, H.H., 2003. Over production of phycocyanin pigment in blue green alga Spirulina sp. and It's inhibitory effect on growth of ehrlich ascites carcinoma cells. J. Med. Sci., 3: 314-324.
CrossRefDirect Link - Barclay, W.R., K.M. Meager and J.R. Abril, 1994. Heterotrophic production of long chain omega-3 fatty acids utilizing algae and algae-like microorganisms. J. Applied Phycol., 6: 123-129.
CrossRefDirect Link - Bhat, Z.F. and H. Bhat, 2011. Milk and dairy products as functional foods: A review. Int. J. Dairy Sci., 6: 1-12.
CrossRefDirect Link - Becker, E.W., 2007. Micro-algae as a source of protein. Biotechnol. Adv., 25: 207-210.
CrossRefDirect Link - Belury, M.A., 2002. Inhibition of carcinogenesis by conjugated linoleic acid: Potential mechanisms of action. J. Nut., 132: 2995-2998.
Direct Link - Boeckaert, C., B. Vlaeminck, V. Fievez, L. Maignien, J. Dijkstra and N. Boon, 2008. Accumulation of trans C18: 1 fatty acids in the rumen after dietary algal supplementation is associated with changes in the butyrivibrio community. Applied Environ. Microbiol., 74: 6923-6930.
CrossRef - Carrillo, S., E. Lopez, M.M. Casas, E. Avila, R.M. Castillo, M.E. Carranco, C. Calvo and F. Perez-Gil, 2008. Potential use of seaweeds in the laying hen ration to improve the quality of n-3 fatty acid enriched eggs. Nineteenth Int. Seaweed Symp. Dev. Applied Phycol., 2: 271-278.
CrossRef - Ming, L.C., R. Nurliyana, A.B. Syah, M.N. Azizah, H.L. Sim and M.Y. Hirzun, 2012. Identification and biochemical composition of a green microalgae. Asian J. Biotechnol., (In press).
Direct Link - Christaki, E., E. Bonos, D. Foskolos, D. Paneri, E. Sarikaki and P.F. Paneri, 2011. The effect of dietary spirulina in infant brain Curr. Opin. Biotechnol., 22: S91-S91.
Direct Link - Colla, L.M., A.L. Muccillo-Baisch and J.A.V. Costa, 2008. Spirulina platensis Effects on the levels of total cholesterol, HDL and triacylglycerols in rabbits fed with a hypercholesterolemic diet. Braz. Arch. Biol. Technol., 51: 405-411.
Direct Link - El-Baz, F.K., A.M. Aboul-Enein, G.S. El-Baroty and H.H. Abd El-Baky, 2002. Anticarcinogenic activity of algal extracts. J. Med. Sci., 2: 243-251.
Direct Link - Folch, J., M. Lees and G.H.S. Stanley, 1957. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem., 226: 497-509.
CrossRefPubMedDirect Link - Franklin, S.T., K.R. Martin, R.J. Baer, D.J. Schingoethe and A.R. Hippen, 1999. Dietary marine algae (Schizochytrium sp.) increases concentrations of conjugated linoleic, docosahexaenoic and transvaccenic acids in milk of dairy cows. J. Nutr., 129: 2048-2054.
Direct Link - Grinstead, G.S., M.D. Tokach, S.S. Dritz, R.D. Goodband and J.L. Nelssen, 2000. Effects of Spirulina platensis on growth performance of weanling pigs. Anim. Feed Sci. Technol., 83: 237-247.
CrossRefDirect Link - Hoa, L.T.P., D.N. Quang, N.T.H. Ha and N.H. Tri, 2011. Isolating and screening mangrove microalgae for anticancer activity. Res. J. Phytochem., 5: 156-162.
CrossRef - Kim, E.J., S.A. Huws, M.R.F. Lee, J.D. Wood, S.M. Muetzel, R.J. Wallace and N.D. Scollan, 2008. Fish oil increases the duodenal flow of long chain polyunsaturated fatty acids and trans-11 18:1 and Decreases 18:0 in steers via changes in the rumen bacterial community. J. Nutr., 138: 889-896.
Direct Link - Kulpys, J., E. Paulauskas, A. Simkus and A. Jeresiunas, 2009. The influence of weed Spirulina platensis on production and profitability of milking cows. Vet. Med. Zoot., 46: 24-29.
Direct Link - Kulpys, J., E. Paulauskas, V. Pilipavicius and R. Stankevicius, 2009. Influence of cyanobacteria Arthrospira (Spirulina) platensis biomass additives towards the body condition of lactation cows and biochemical milk indexes. Agron. Res., 7: 823-835.
Direct Link - Lee, A.H. and N. Hiramatsu, 2011. Role of n-3 series polyunsaturated fatty acids in cardiovascular disease prevention. Nutr. Diet. Suppl., 3: 93-100.
CrossRefDirect Link - Lock, A.L. and D.E. Bauman, 2004. Modifying milk fat composition of dairy cows to enhance fatty acids beneficial to human health. Lipids, 39: 1197-1206.
CrossRef - Lordan, S., R.P. Ross and C. Stanton, 2011. Marine bioactives as functional food ingredients: Potential to reduce the incidence of chronic diseases. Mar. Drugs, 9: 1056-1100.
PubMed - Madhusudan, C., S. Manoj, K. Rahul and C.M. Rishi, 2011. Seaweeds: A diet with nutritional, medicinal and industrial value. Res. J. Med. Plant, 5: 153-157.
CrossRefDirect Link - Mirada, M.S., R.G. Cintra, S.B.M. Barros and J. Mancini-Filho, 1998. Antioxidant activity of the microalga Spirulina maxima. Braz. J. Med. Biol. Res., 31: 1075-1079.
CrossRefDirect Link - Nasseri, A.T., S. Rasoul-Amini, M.H. Morowvat and Y. Ghasemi, 2011. Single cell protein: Production and process. Am. J. Food Technol., 6: 103-116.
CrossRefDirect Link - Papadopoulos, G., C. Goulas, E. Apostolaki and R. Abril, 2002. Effects of dietary supplements of algae, containing polyunsaturated fatty acids, on milk yield and the composition of milk products in dairy ewes. J. Dairy Res., 69: 357-365.
PubMed - Rasool, M., E.P. Sabina, P. Nithya and K. Lavanya, 2008. Studies on the analgesic, antipyretic and ulcerogenic properties of Spirulina fusiformis in mice. J. Pharmacol. Toxicol., 3: 47-52.
CrossRefDirect Link - Simkus, A., V. Oberauskas, J. Laugalis, R. Zelvyte and I. Monkeviciene et al., 2007. The effect of weed Spirulina platensis on the milk production in cows. Veterinarija Ir Zootechnika T., 38: 74-77.
Direct Link - Simopoulos, A.P., 2002. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharmacother., 56: 365-379.
CrossRefDirect Link - Singh, A.P., C.A. Avramis, J.K. Kramer and A.G. Marangoni, 2004. Algal meal supplementation of the cows diet alters the physical properties of milk fat. J. Dairy Res., 71: 66-73.
PubMed - Simpore, J., F. Zongo, Y. Ouattara, F. Kabore and D. Dansou et al., 2005. Nutrition rehabilitation of the HIV-infected and negative undernourished children utilizing spiruline. Pak. J. Biol. Sci., 8: 589-595.
CrossRefDirect Link - Singh, V.P. and N. Sachan, 2011. Nutraceutical properties of milk and milk products: A review. Am. J. Food Technol., 6: 864-869.
CrossRefDirect Link - Singh, V.P. and N. Sachan, 2011. Conjugated lenoleic acid: A good fat for human health-A review. Int. J. Dairy Sci., 6: 165-171.
CrossRefDirect Link - Spolaore, P., C. Joannis-Cassan, E. Duran and A. Isambert, 2006. Commercial applications of microalgae. J. Biosci. Bioeng., 101: 87-96.
CrossRefPubMedDirect Link