
Jun Fang
National R & D Center for Egg Processing, Huazhong Agricultural University, Wuhan, 430070, China
Meihu Ma
National R & D Center for Egg Processing, Huazhong Agricultural University, Wuhan, 430070, China
Yongguo Jin
National R & D Center for Egg Processing, Huazhong Agricultural University, Wuhan, 430070, China
Ning Qiu
National R & D Center for Egg Processing, Huazhong Agricultural University, Wuhan, 430070, China
Chan Wang
National R & D Center for Egg Processing, Huazhong Agricultural University, Wuhan, 430070, China
Guodong Ren
National R & D Center for Egg Processing, Huazhong Agricultural University, Wuhan, 430070, China
Xin Huang
National R & D Center for Egg Processing, Huazhong Agricultural University, Wuhan, 430070, China
Asian Journal of Animal and Veterinary Advances
Year: 2012 | Volume: 7 | Issue: 7 | Page No.: 556-567







ABSTRACT
Egg white was considered to play an important role in bacterial-resistance nature of developing embryo especially during early incubation. Previous works were not quite informative in the antimicrobial action of egg white. This study was performed to investigate the survival ability of Salmonella enteritidis in egg white during early incubation and to tentatively reveal underline anti-Salmonella enteritidis mechanisms of egg white using the combination of flow cytometric analysis and fluorescent strategy. Such properties were also tested at different incubation temperature and pH. The traditional plate count method was also included in this study and the results were in accordance with those of flow cytometric assessment. Following egg white treatment, Salmonella enteritidis was discriminated into three subpopulations by flow cytometric assessment: viable, dead and membrane-compromised cells. Furthermore, the results showed that with the extension of incubation time, the population giving green fluorescence increased steadily while red fluorescence emittance didn’t change a lot. The incubation temperature and pH were both significant factors in anti-Salmonella enteritidis properties of egg white. Besides, the main target for antimicrobial factors in egg white on Salmonella enteritidis cells was membrane integrity and the action mechanism of egg white was bacteriostatic rather than bactericidal effect during early incubation.
PDF Abstract XML References Citation
Received: November 02, 2011;
Accepted: January 25, 2012;
Published: February 25, 2012
How to cite this article
Jun Fang, Meihu Ma, Yongguo Jin, Ning Qiu, Chan Wang, Guodong Ren and Xin Huang, 2012. Assessment of Salmonella enteritidis Viability in Egg White during Early Incubation Stages by Fluorescent Staining Method. Asian Journal of Animal and Veterinary Advances, 7: 556-567.
DOI: 10.3923/ajava.2012.556.567
URL: https://scialert.net/abstract/?doi=ajava.2012.556.567
DOI: 10.3923/ajava.2012.556.567
URL: https://scialert.net/abstract/?doi=ajava.2012.556.567
INTRODUCTION
Egg white would provide immune protection to the developing embryo and is considered to be an unfavourable medium for microbial growth (Alabdeh et al., 2011; Stevens, 1996; Tranter and Board, 1984); Lysozyme and ovotransferrin are well-characterized antimicrobials in egg white (Messens et al., 2004). Besides, there are some other biologically active substances with antimicrobial properties in egg white such as cystatin (Jerzy and Pysniak, 2009; Trziszka et al., 2004). Previous researches showed that dramatic changes in egg white would occur during egg incubation, such as pH, viscosity, stability of antimicrobial proteins (Araki et al., 2000; Benton Jr et al., 2001; Tona et al., 2010). These changes in egg white might be important for immune protection to the developing embryo. However, it has been revealed that Salmonella enteritidis possessed a strong ability to survive and persist for a long period of time in egg white (Cox et al., 1990; Manijeh et al., 2008; Prakash et al., 2005). Furthermore, researches showed that eggs inoculated with high numbers of Salmonella enteritidis could still hatch (Cason et al., 1994). These findings implicated that Salmonella enteritidis have the ability to transmit from generation to generation through eggs which would bring a series of hazards (Gast et al., 2009). To our knowledge, little was known about the viability state and survival mechanism of Salmonella enteritidis in egg white during incubation up to now.
Application of multiple stains combined with Flow cytometric (FCM) techniques could provide valuable details about cell physiology and allow analysis of metabolic activity and membrane integrity (Barbesti et al., 2000; Davey and Kell, 1996; Mandal et al., 2011). It also has been proved to be useful for the assessment of viability state of bacteria. The viability state of Salmonella typhimurium in dairy products was evaluated by using fluorescent probe Fluorescein Isothiocyanate (FITC) and ethidium bromide (McClelland and Pinder, 1994). In this study, we conducted a rapid assessment of Salmonella enteritidis based on FCM detection with a double-staining strategy using carboxyfluorescein diacetate (cFDA) and Propidium Iodide (PI). cFDA is a non-fluorescent dye which would be converted into a fluorescent compound, carboxyfluorescein (cF) by cellular unspecific esterases (Petit et al., 1993). However, the complete outer membrane of gram-negative bacteria has been proved to hinder the entering of viability marker cFDA into the cell (Vaara, 1992). Thus, the positive reaction of cFDA indicates a membrane compromised state of gram-negative bacteria. In contrary, the negative reaction indicates a viable (membrane integrity) state. As to PI, it is a nucleotide-binding probe which was considered not to penetrate and stain cells unless membrane integrity didn’t exist (Ananta et al., 2004) and frequently used to indicate death of bacteria. The aim of this study was to illustrate physiological state of Salmonella enteritidis cells after explosure to egg white during early incubation by using a rapid assessment based on flow cytometric analysis in combination with double-staining of cFDA and PI.
MATERIALS AND METHODS
Preparation of cell suspension: Three milliliters of original Salmonella enteritidis(ATCC 13076) cell suspension in Tryptic Soy Broth (TSB) and one milliliter of sterilized glycerol were mixed and then stored at -80°C until use. It was streaked onto Tryptic Soy Agar (TSA) and then cultured at 37°C for 24 h after thawing at room temperature. As soon as the strain was resuscitated, a loop of SE from TSA was transferred to 100 mL of TSB to grow overnight at 37°C. The harvested bacteria cells were centrifugated at 8000 rpm for 5 min at 4°C. After washed 3 times by 50 mM phosphate buffered saline, pH 7.4 (PBS), the Salmonella enteritidis was adjusted to an OD490 of 1.5 (ca. 1.8x1010 CFU mL-1). The suspension was then used for inoculating.
Egg white inoculating: Fresh fertilized eggs from Single Comb White Leghorn laid within 24 h were collected from the poultry research centre farm of Huazhong Agricultural University and used in this study. Eggs were incubated at 38°C and 65-75% relative humidity in a forced air incubator for 0, 1, 2, 3 days (Kamanli et al., 2010; Alabi et al., 2012; Seker et al., 2004). Investigations were carried out in daily intervals. At each experimental time, ten eggs were disinfected by immersion in 75% ethanol and under aseptic conditions, they were dried, cracked, and the egg white was collected into a sterile container and thoroughly mixed. Two milliliter of inoculum was injected into 18 mL of albumen samples and then incubated at 38°C for 8 h. To study the effect of incubation temperature on Salmonella enteritidis viability in egg white, fresh fertilized egg white inoculated with Salmonella enteritidis was incubated for 8 h at 4, 25 and 38°C (King’ori, 2011). The pH of fresh fertilized egg white (pH 9.16) was adjusted to 7.0 and 9.56 by using NaOH and HCl solution to research in the changes of pH of egg white on Salmonella enteritidis viability during early incubation stages (Yesillik et al., 2011; Wagdy et al., 2011). After double staining with cFDA and PI, all the samples were assessed on a flow cytometer.
Fluorescence labeling: Carboxyfluorescein diacetate (cFDA, Beyotime Institute of Biotechnology, China) and propidium iodide (PI, Beyotime Institute of Biotechnology, China) were used in this study to determine the esterase activity and membrane integrity of Salmonella enteritidis cells treated by egg white. Untreated controls and egg white treated samples were filtrated by 8 μm cellulose acetate filter. After that, samples were centrifugated at 8000 rpm for 5 min at 4°C which was followed by washing and resuspension. An appropriate volume of cFDA was added to produce a target concentration of 2 μl mL-1 cell suspension. The cells were incubated for 20 min at 37°C and then excessive cFDA was removed by centrifugation before PI staining. The concentration for PI staining was 2 μL mL-1 cell suspension. The cells were incubated in ice bath for 10 min before FCM analysis.
Observation with fluorescence microscopy: Stained cells were imaged on Olympus BX51 microscope equipped with a OLYMPUS DP71 CCD (OLYMPUS, Japan). Aliquots of 10 μL ’of each cell suspension were transferred to a clean glass slide under coverslip. All of the slides were one by one put on the microscopy. CFDA-stained and PI-stained cells were visualized by excitation with blue light (400-440 nm) and UV light (330-385 nm). In addition, Salmonella enteritidis controls added glutaraldehyde with a final concentration of 10 g L-1 cell suspension were also included in this observation.
Flow cytometric measurement: The stained Salmonella enteritidis cells were analyzed with a BD FACSCalibur (America) flow cytometer equipped with a 15 mW, 488 nm air cooled laser. The flow rate in this experiment was set at 600-800 events per second, up to a total sum of 50000 events per sample. The green fluorescence of cF-stained cells was collected in the FL1 channel (525±20 nm) and red fluorescence of PI-labelled was collected in the FL2 channel (620±15 nm).
Bacterial plate counts: Enumeration of Salmonella enteritidis in egg white was carried out by serial decimal dilution of cell suspensions using 9 mL of sterile PBS. Afterwards, 250 μL portions of the appropriate dilutions of controls and treated samples were added on TSA plates and incubated for 48 h at 37°C.
RESULTS AND DISCUSSION
Basic staining behavior of Salmonella enteritidis: The photographs taken with fluorescence microscopy displayed that viable Salmonella enteritidis cells unstained or stained with cFDA could hardly emit any fluorescence (Fig. 1a, b). While membrane damage induced by 10 g L-1 Glutaraldehyde (GTA) would promote cFDA staining on Salmonella enteritidis (Fig. 1c). It was obvious that heat-killed (70°C, 30 min) Salmonella enteritidis cells without viability were prone to be labelled by PI and to give a detectable red fluorescence (Fig. 1d). Both green and red fluorescence can be detected in Salmonella enteritidis cells after egg albumen treatment following with double staining (Fig. 1e) which indicated that Salmonella enteritidis in egg white can be divided into different subpopulations: viable, dead and membrane-compromised.
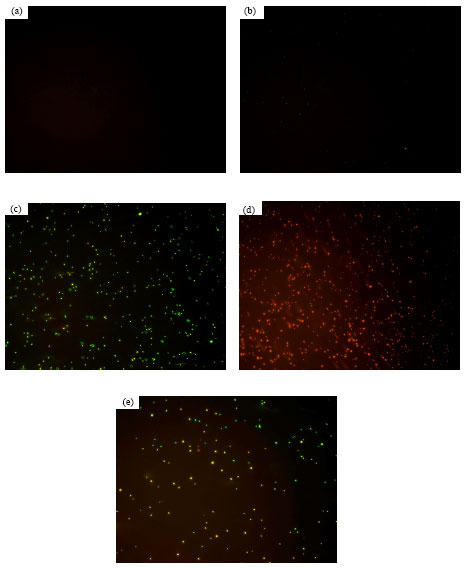 | |
| Fig. 1(a-e): | Fluorescent microscopy images of viable Salmonella enteritidis (a) unstained, (b) stained solely with cFDA, (c) GTA treated Salmonella enteritidis stained with cFDA, (d) heat-damaged (70°C, 30 min) Salmonella enteritidis stained with PI and (e) egg white treated Salmonella enteritidis double-stained with cFDA and PI |
To differentiate bacterial population based on their fluorescence properties as well as to assess treatment effect on Salmonella enteritidis, the dual-parameter density plot of the green fluorescence (x-axis) and the red fluorescence (y-axis) was used (Ananta et al., 2005). Fluorescence density plots of viable Salmonella enteritidis cells and heat-treated cells following staining with cFDA and PI were shown in Fig. 2. The results showed that viable cells that could be stained by cFDA were only 2-3% of total sum (Fig. 2a, in quadrants 2, 4) which was in agreement with the report that cFDA can not permeate into the outer membrane of gram-negative bacteria (Vaara, 1992), 96.3% of Salmonella enteritidis cells were stained with PI after heat treatment for 30 min at 70°C (Fig. 2b, in quadrant 1). Double staining treatment of viable Salmonella enteritidis showed that most of the cell particles are distributed in quadrant 3 (Fig. 2c).
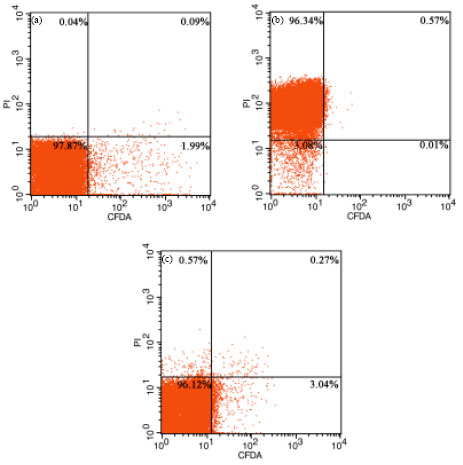 | |
| Fig. 2(a-c): | (a)Fluorescence density plots of intact Salmonella enteritidis cells solely stained with cFDA, (b) heat-treated cells stained with PI and (c) intact Salmonella enteritidis cells double stained with cFDA and PI. Heat treatment at 70°C was performed to yield dead, membrane-compromised cells. The figures (in %) in different quadrant are associated with the percentage of the cells in the corresponding quadrant |
Changes of viability of Salmonella enteritidis in egg white during early incubation: Figure 3a-d showed fluorescence properties of Salmonella enteritidis in egg white collected from the early incubation stages. The population accumulating cF (in quadrant 2 and 4) increased steadily from about 46 to 96% with the extension of incubation time, suggesting that albumen with longer incubation time facilitated penetration of cFDA and its transition into cF by intracellular esterase. It was generally considered that the ability of gram-positive bacteria to accumulate and retain cF was an indicator of metabolic activity because the conversion was dependent on active esterase. However, gram-negative bacteria had a thin cell wall surrounded by an outer membrane as a selective permeability barrier that could limit the passage of certain substances (Kim et al., 2007). Thus, gram-negative bacteria showed a deviating behaviour in their fluorescence property. Only when their special structure of outer membrane was damaged, cFDA could enter into the cells to interact with the esterase and to yield green fluorescence. From this point of view, staining cells of gram-negative bacteria with cFDA would not show green fluorescence unless membrane injury and esterase activity simultaneously exist. Changes of fluorescence property of Salmonella enteritidis in egg white indicated that longer incubation time could result in more serious membrane injury.
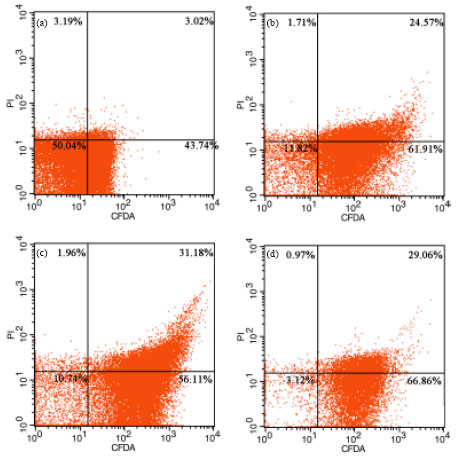 | |
| Fig. 3(a-d): | Fluorescence density plots of Salmonella enteritidis cells treated by egg white collected from fertilized eggs incubated for (a) 0 day, (b) 1 day, (c) 2 day and (d) 3 day. The figures (in %) in different quadrant are associated with the percentage of the cells in the corresponding quadrant |
The results of traditional plate count method also showed that anti-Salmonella enteritidis activity of egg white was improved as incubation progressed during the early embryonic development (Fig. 6a). Compared the results of flow cytometry with plate counts, it can be inferred that some cells of bacteria with membrane ruptured (in quadrant 4) in egg white could regrow on agar. In another side, the loss of membrane integrity was at least related to the loss of viability of Salmonella enteritidis. However, the percentages of Salmonella enteritidis cells labelled by PI in quadrant 1 didn’t change a lot and all were considerably low (Fig. 3a-d). These findings leaded to a presumption that the major part of Salmonella enteritidis cells treated by egg albumen during the first 3 days of incubation was not dead, indicating the bactericidal activities of egg albumen during early incubation were so limited that they could not reach directly to the death of bacteria cells.
Effect of incubation temperature on Salmonella enteritidis viability in egg white: Eggs are incubated at hens’ body temperature (38°C), so, we conducted a series of experiments so as to figure out the effect of temperature on the anti-Salmonella enteritidis properties of egg white. The viability status of Salmonella enteritidis cells treated by egg white at different temperature was depicted in Fig. 4a-c.
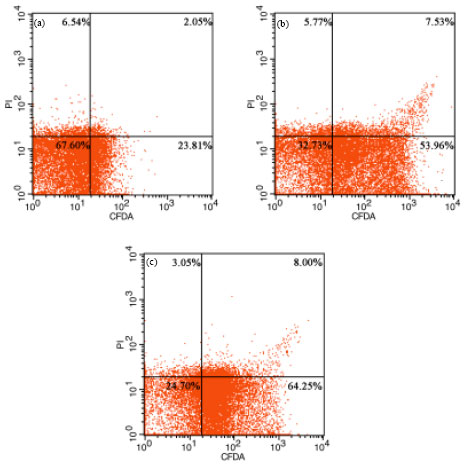 | |
| Fig. 4(a-c): | Fluorescence density plots of Salmonella enteritidis cells inoculated in egg white following incubated at (a) 4°C, (b) 25°C and (c) 38°C for 8 h. The figures (in %) in different quadrant are associated with the percentage of the cells in the corresponding quadrant |
Percentage of cFDA stained cells was 25.86% at 4°C, 61.49% at 25°C and 72.25% at 38°C of total sum of cells. We selected these three temperatures in this study in order to represent typical refrigeration temperature, room temperature and incubation temperature, respectively. The result suggested that egg white incubated at 38°C (typical incubation temperature) was most able to facilitate the penetration and conversion of cFDA. So, egg white incubated at 38°C possessed the strongest ability to prevent Salmonella enteritidis growth. The result was in accordance with that obtained from the traditional plate count method (Fig. 6b). Researches have demonstrated that egg white at high temperature kept higher bacteriostatic properties than at low temperature (Aykes and Taylor, 1956). Avian incubation would provide additional protection through increasing the temperature of the egg white to levels at which its antimicrobial enzymes work optimally (Tranter and Board, 1984) and that exceed the optimum for growth of most microorganisms (Cook et al., 2005). Previous reports have also discovered that there were nuclease activities inside egg albumen which could lacerate both naked and intracellular bacterial DNA (Lu et al., 2003). The nuclease activities were likely to be dependent on temperature, showing higher at 37°C and lower at 4 and 25°C.
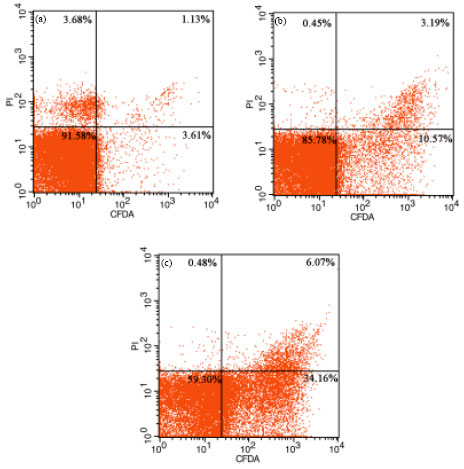 | |
| Fig. 5(a-c): | Fluorescence density plots of Salmonella enteritidis cells inoculated in pH adjusted egg white: (a) 7.0, (b) 9.16 and (c) 9.5. The figures (in %) in different quadrants are associated with the percentage of the cells in the corresponding quadrant |
Based on this study, it was confirmed that the antimicrobial activity of egg white was increased under the incubation conditions. And the anti-Salmonella enteritidis mechanism of egg white during early incubation would be cell membrane damage.
Effect of rising of pH during early incubation on Salmonella enteritidis viability in egg white: The pH of chicken egg white has been proved to be an important antibacterial defence factor in preventing bacteria growth. During early incubation stages, the albumen pH increased rapidly as a result of sudden mounting up of internal temperature which in turn accelerated CO2 loss from egg white. In this study, it was evaluated the effect of changes in pH during early incubation on the viability state of Salmonella enteritidis in egg white. The pH of fresh egg white used in this study was 9.16. After two days of incubation, the pH of egg white would rised to as high as 9.56 (data not shown). Thus, pH of egg white samples was adjusted by acid and alkali solutions to 7.0, 9.16 and 9.56 to study the effect of pH on Salmonella enteritidis viability in egg white during incubation. The result showed that cells in quadrant 2 and 4 were 13.76% at pH 9.16, 40.23% at pH 9.56 and lower than 5% at pH 7.0 (Fig. 5a-c). It can see that the anti-Salmonella enteritidis activity was increased considerably due to the increase of pH. The survival rate of Salmonella enteritidis in response to egg white with different pH confirmed the result of assessment by flow cytometry (Fig. 6c).
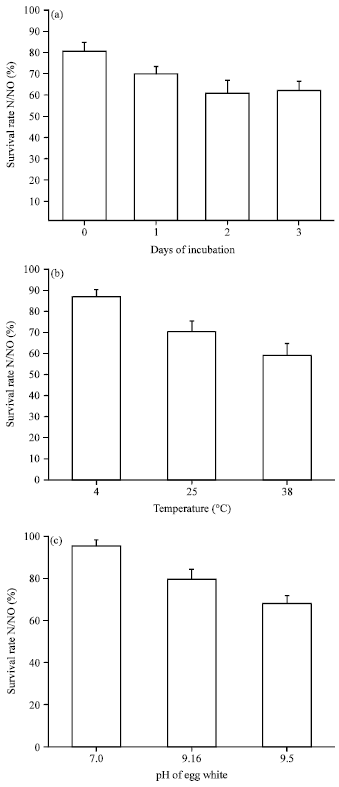 | |
| Fig. 6(a-c): | Effect of different incubation period (a), incubation temperature (b) and pH during incubation (c), on survival rate of Salmonella enteritidis cells in egg white. Data are the mean of three replicates of egg white treatment. Error bars indicate the standard deviations of the mean |
Previous research has shown that egg white was much more toxic to the bacteria at pH 9-10 rather than pH 6-8 (Sharp and Whitaker, 1927). That was probably because high alkalinity of egg white harassed bacterial iron metabolism which prevented bacteria from gaining enough iron for their growth (Tranter and Board, 1984).
CONCLUSION
Based on double-staining strategy by using cFDA and PI, it was found that Salmonella enteritidis in egg white could be differentiated into three subpopulations: viable, dead and membrane-compromised cells. As incubation progressed, the viability state of Salmonella enteritidis in egg white would change. The main target for antimicrobial factors in egg white was membrane integrity. Moreover, the flow cytometric assessment allowed us to monitor that bacteriostatic properties were much stronger and more obvious than bactericidal activities of egg white during early incubation. It was generally believed that many factors synergistically induce the antimicrobial properties of egg white during early incubation. Based on the data from this study, we could reasonably consider that temperature and pH play a significant role in controlling Salmonella enteritidis growth in egg white.
ACKNOWLEDGEMENT
This study was supported by the earmarked fund for Modern Agro-industry Technology Research System. Project code no: CARS-41-K23.
REFERENCES
- Alabdeh, M., V. Lechevalier, F. Nau, M. Gautier and M.F. Cochet et al., 2011. Role of incubation conditions and protein fraction on the antimicrobial activity of egg white against salmonella enteritidis and escherichia coil. J. Food. Prot., 74: 24-31.
PubMed - Alabi, O.J., J.W. Ng'ambi and D. Norris, 2012. Effect of egg weight on physical egg parameters and hatchability of indigenous venda chickens. Asian J. Anim. Vet. Adv., 7: 166-172.
CrossRefDirect Link - Ananta, E., V. Heinz and D. Knorr, 2004. Assessment of high pressure induced damage on Lactobacillus rhamnosus GG by flow cytometry. Food. Microbiol., 21: 567-577.
CrossRefDirect Link - Ananta, E., D. Voigt, M. Zenker, V. Heinz and D. Knorr, 2005. Cellular injuries upon exposure of Escherichia coli and Lactobacillus rhamnosus to high-intensity ultrasound. J. Appl. Microbiol., 99: 271-278.
PubMed - Araki, T., Y. Kuwamura, H. Kuwahara, S. Masuda and T. Torikata, 2000. Study of the changes of protein and lysozyme of egg white on embryonic development and storage. Proc. School Agric. Kyushu Tokai Uni., 19: 26-36.
Direct Link - Aykes, J.C. and B. Taylor, 1956. Effect of temperature on microbial proliferation in shell eggs. Applied Microbiol., 4: 355-359.
Direct Link - Barbesti, S., S. Citterio, M. Labra, M.D. Baroni, M.G. Neri and S. Sgorbati, 2000. Two and three-color fluorescence flow cytometric analysis of immunoidentified viable bacteria. Cytometry, 40: 214-218.
CrossRef - Benton, Jr.C.E., T.J. Walsh and J. Brake, 2001. Effects of presence of a blastoderm on albumen height and pH of broiler hatching eggs. Poult. Sci., 80: 955-957.
CrossRefDirect Link - Cason, J.A., N.A. Cox and J.S. Bailey, 1994. Transmission of Salmonella typhimurium during hatching of broiler chicks. Avian. Dis., 38: 583-588.
PubMed - Cook, M.I., S.R. Beissinger, G.A. Toranzos, R.A. Rodriguez and W.J. Arendt, 2005. Microbial infection affects egg viability and incubation behavior in a tropical passerine. Behav. Ecol., 16: 30-36.
CrossRef - Cox, N.A., J.S. Bailey, J.M. Mauldin and L.C. Blankenship, 1990. Presence and impact of Salmonella contamination in commercial broiler hatcheries. Poult. Sci., 69: 1606-1609.
PubMedDirect Link - Davey, H.M. and D.B. Kell, 1996. Flow cytometry and cell sorting of heterogeneous microbial populations: the importance of single-cell analyses. Microbial. Rev., 60: 641-696.
PubMed - Jerzy, R. and D.S. Pysniak, 2009. Antimicrobial defence mechanisms of chicken eggs and possibilities for their use in protecting human and animal health. Annales UMCS Medicina Veterinaria, 64: 1-8.
Direct Link - Gast, R.K., J. Guard-Bouldin, R. Guraya and P.S. Holt, 2009. Effect of prior passage through laying hens on invasion of reproductive organs by Salmonella enteritidis. Int. J. Poult. Sci., 8: 116-121.
CrossRefDirect Link - Seker, I., S. Kul and M. Bayraktar, 2004. Effects of parental age and hatching egg weight of Japanese quails on hatchability and chick weight. Int. J. Poult. Sci., 3: 259-265.
CrossRefDirect Link - Trziszka, T., Y. Saleh, W. Kopeae, M. Siewinski and E. Wesierska, 2004. Effect of hen`s age on the level of cystatin in the chicken egg white. Int. J. Poult. Sci., 3: 471-477.
CrossRefDirect Link - Tona, K., K. Agbo, B. Kamers, N. Everaert, H. Willemsen, E. Decuypere and M. Gbeassor, 2010. Comparison of lohmann white and lohmann brown strains in embryo physiology. Int. J. Poult. Sci., 9: 907-910.
CrossRefDirect Link - Kamanli, S., I. Durmus and S. Demir, 2010. Hatching characteristics of abnormal eggs. Asian J. Anim. Vet. Adv., 5: 271-274.
CrossRefDirect Link - Kim, Y.M., S. Farrah and R.H. Baney, 2007. Membrane damage of bacteria by silanols treatment. Electron. J. Biotechn., 10: 252-259.
Direct Link - Kingori, A.M., 2011. Review of the factors that influence egg fertility and hatchabilty in poultry. Int. J. Poult. Sci., 10: 483-492.
CrossRefDirect Link - Lu, S., P.B. Killoran and L.W. Riley, 2003. Association of salmonella enterica Serovar enteritidis YafD with resistance to chicken egg albumen. Infect. Immun., 71: 6734-6741.
CrossRef - Manijeh, M., J. Mohammad and K.K. Roha, 2008. Biofilm formation by Salmonella enteritidis on food contact surfaces. J. Boil. Sci., 8: 502-505.
CrossRefDirect Link - Mandal, P.K., A.K. Biswas, K. Choi and U.K. Pal, 2011. Methods for rapid detection of foodborne pathogens: An overview. Am. J. Food Technol., 6: 87-102.
CrossRefDirect Link - McClelland, R.G. and A.C. Pinder, 1994. Detection of salmonella typhimurium in dairy products with flow cytometry and monoclonal antibodies. Applied Environ. Microbial, 60: 4255-4262.
Direct Link - Messens, W., L. Duboccage, K. Grijspeerdt, M. Heyndrickx and L. Herman, 2004. Growth of Salmonella serovars in hen's egg albumen as affected by storage prior to inoculation. Food Microbiol., 21: 25-32.
CrossRef - Petit, J.M., M. Denis-Gay and M.H. Ratinaud, 1993. Assessment of fluorochromes for cellular structure and function studies by flow cytometry. Biol. cell., 78: 1-13.
PubMed - Prakash, B., G. Krishnappa, L. Muniyappa and B.S. Kumar, 2005. Epidemiological characterization of avian Salmonella enterica serovar infections in India. Int. J. Poult. Sci., 4: 388-395.
CrossRefDirect Link - Sharp, P.F. and R. Whitaker, 1927. The relation of the hydrogen concentration of egg white to its germicidal action. J. Bacteriol., 14: 17-46.
Direct Link - Wagdy, S.M., S.H. Mohamed and F.S. Taha, 2011. Solubility pattern of simmondsins, proteins and phenolics of defatted jojoba meal. Am. J. Food Technol., 6: 963-973.
CrossRefDirect Link - Tranter, H.S. and R.G. Board, 1984. The influence of incubation temperature and pH on the antimicrobial properties of hen egg albumen. J. Applied Microbiol., 56: 53-61.
CrossRefDirect Link - Vaara, M., 1992. Agents that increase the permeability of the outer membrane. Microbiol. Rev., 56: 395-411.
PubMedDirect Link - Yesillik, S., N. Yildirim, A. Dikici, A. Yildiz and S. Yesillik, 2011. Antibacterial effects of some fermented commercial and homemade dairy products and 0.9% lactic acid against selected foodborne pathogens. Asian J. Anim. Vet. Adv., 6: 189-195.
CrossRefDirect Link