
Abulizi Wusiman
Laboratory of Animal Genetics, Breeding and Reproduction, Ministry of Agriculture and National Engineering Laboratory for Animal Breeding, College of Animal Science and Technology, China Agricultural University, Beijing 100193, People�s Republic of China
Yan-Ping Wang
Laboratory of Animal Genetics, Breeding and Reproduction, Ministry of Agriculture and National Engineering Laboratory for Animal Breeding, College of Animal Science and Technology, China Agricultural University, Beijing 100193, People�s Republic of China
Kang Ren
Laboratory of Animal Genetics, Breeding and Reproduction, Ministry of Agriculture and National Engineering Laboratory for Animal Breeding, College of Animal Science and Technology, China Agricultural University, Beijing 100193, People�s Republic of China
Guang-Bin Zhou
Laboratory of Animal Genetics, Breeding and Reproduction, Ministry of Agriculture and National Engineering Laboratory for Animal Breeding, College of Animal Science and Technology, China Agricultural University, Beijing 100193, People�s Republic of China
Xiang-Wei Fu
Laboratory of Animal Genetics, Breeding and Reproduction, Ministry of Agriculture and National Engineering Laboratory for Animal Breeding, College of Animal Science and Technology, China Agricultural University, Beijing 100193, People�s Republic of China
Lun Suo
Laboratory of Animal Genetics, Breeding and Reproduction, Ministry of Agriculture and National Engineering Laboratory for Animal Breeding, College of Animal Science and Technology, China Agricultural University, Beijing 100193, People�s Republic of China
Zhi-Qiang Fan
Laboratory of Animal Genetics, Breeding and Reproduction, Ministry of Agriculture and National Engineering Laboratory for Animal Breeding, College of Animal Science and Technology, China Agricultural University, Beijing 100193, People�s Republic of China
Liang Wang
Laboratory of Animal Genetics, Breeding and Reproduction, Ministry of Agriculture and National Engineering Laboratory for Animal Breeding, College of Animal Science and Technology, China Agricultural University, Beijing 100193, People�s Republic of China
Shi-En Zhu
Laboratory of Animal Genetics, Breeding and Reproduction, Ministry of Agriculture and National Engineering Laboratory for Animal Breeding, College of Animal Science and Technology, China Agricultural University, Beijing 100193, People�s Republic of China
Asian Journal of Animal and Veterinary Advances
Year: 2012 | Volume: 7 | Issue: 4 | Page No.: 299-308







ABSTRACT
In order to provide more options to sheep artificial insemination (AI) technology this study evaluated the influence of preservation methods on sperm quality and subsequent fertility in Small-tail Hun sheep. For the following treatments: Diluted fresh semen as a control, diluted semen stored at 4°C for 24, 48, 72, 96 or 120 h, Separate aliquots of diluted semen were stored at 23°C for 24 or 48 h and diluted semen stored at -196°C, The semen was evaluated for sperm viability, acrosomal integrity, mitochondrial function and capacitation status using Propidium Iodide (PI), fluoresceinated peanut lectin (FITC-PNA), 5,5',6,6'-tetrachloro-1,1',3,3'-tetraethylbenzimidazolyl-carbocya nine iodide (JC-1) and chlortetracycline staining, respectively. Proportions of spermatozoa showing high mitochondrial membrane potential and an incapacitated status were lower (p<0.05) in frozen semen than in samples stored at either 4°C for 72 h or at 23°C for 24 h. Sperm viability did not differ (p>0.05) among: Fresh semen, semen stored at 4°C for 48 h and semen stored at 23°C for 24 h. Following cervical insemination, semen stored at 4°C for 72 h or at 23°C for 24 h resulted in similar pregnancy rates (p>0.05) to those from fresh sperm (16/23, 69.6%, 19/25, 76.0% and 22/28, 78.6%), all of which were higher (p<0.05) than those from semen cryopreserved at -196°C (12/24, 50.0%). In conclusion, semen stored at either 4°C for 72 h or at 23°C for 24 h is structurally superior to semen cryopreserved at -196°C and achieved similar pregnancy rates as from AI using fresh semen.
PDF Abstract XML References Citation
Received: October 04, 2011;
Accepted: January 02, 2012;
Published: January 19, 2012
How to cite this article
Abulizi Wusiman, Yan-Ping Wang, Kang Ren, Guang-Bin Zhou, Xiang-Wei Fu, Lun Suo, Zhi-Qiang Fan, Liang Wang and Shi-En Zhu, 2012. Semen Storage at 23, 4 or -196°C and its Application to Artificial Insemination in Small-tail Han Sheep. Asian Journal of Animal and Veterinary Advances, 7: 299-308.
DOI: 10.3923/ajava.2012.299.308
URL: https://scialert.net/abstract/?doi=ajava.2012.299.308
DOI: 10.3923/ajava.2012.299.308
URL: https://scialert.net/abstract/?doi=ajava.2012.299.308
INTRODUCTION
Small-tail Han sheep, a popular breed for meat and fur production in China, is characterized by high fecundity. In order to fertilize large numbers of offspring with semen of outstanding rams semen required to be transported from centers of collection to the sites of insemination (Salamon and Maxwell, 2000). Effective semen storage is important for effective genetic improvement and selection programs for this breed. There are two main methods to store semen (1) a liquid (unfrozen) state, using reduced temperatures or other means to depress sperm metabolism and (2) in a frozen state which involved preservation at sub-zero temperatures. Both purposes are achieved by methods that reduced or arrested the metabolism of spermatozoa and thereby prolonged their fertile life (Salamon and Maxwell, 2000).
Besides, the necessity to use the rams over extended periods, or at different times of the year, stimulated research on storage of spermatozoa under frozen conditions (Salamon and Maxwell, 2000). To develop technology for effective cryopreservation of ram semen at -196°C, many studies have focused on selection of freezing media (Purdy, 2006) protocols for freezing (Gravance et al., 1997; Blash et al., 2000) and methods to remove seminal plasma (Maxwell and Johnson, 1999). Ram semen is sensitive to cold shock (Salamon and Maxwell, 1995a). Thus, the freezing-thawing procedures during cryopreservation can induce ultrastructural, biochemical and functional changes in sperm (Peris et al., 2004; Marti et al., 2008; Watson, 1995) can result in reduced fertilizing ability (Salamon and Maxwell, 1995b). Post-thaw sperm spermatozoa following semen cryopreservation ranging from 47 to 60.1% has been reported in Small-tail Han sheep (Hu et al., 2002; Wang et al., 2004) but pregnancy rates following AI with frozen-thawed semen have not been reported.
Because there have been few comparable systematic investigations, the aim of the present study was (1) to assess viability, acrosomal integrity, Mitochondrial Membrane Potential (MMP) and capacitation status of fresh semen and semen stored at 4, 23 or -196°C, (2) and to compare the pregnancy rates after artificial insemination with semen stored at 4, 23 or -196°C in Small-tail Han sheep.
MATERIALS AND METHODS
The duration of this experiment is from August 2007 to March 2009. Unless indicated otherwise, all chemicals and media were purchased from Sigma Chemical Co. St. Louis, MO, USA and all plastics from Corning, Inc. Corning, NY, USA.
Animals and semen collection: Semen was collected with an artificial vagina from one of the 20 mature Small-tail Han rams whose semen volume (ranging from 0.75 to 2 mL), macroscopic visual mass activity, sperm concentration (all samples≥2.5x109 spermatozoa/mL), motility (all samples≥70%), morphology (all samples≥10% total sperm abnormalities) and the motility after cryopreservation over 35% at Beijing Breed Selection Centre. The fresh semen samples were collected three times a week, on different and non-consecutive days.
Semen dilution, cooled preservation, freezing and thawing: The cooled preservation medium contained 80% (v:v) basal solution (100 mL distilled water, 3.2 g glucose, 3.2 g sodium citrate, penicillin (10 IU), streptomycin (10 IU)) and 20% (v:v) egg yolk, while the cryopreservation medium was composed of cooled preservation extender supplemented with 6% (v:v) glycerol. Prior to use, both media were warmed to 37°C in sterile glass vials.
Within 10-20 min after semen collection, fresh semen was randomly allocated into two portions: (1) Semen diluted with cooled preservation extender, (2) Semen diluted with cryopreservation extender to a final concentration of 5x108 sperm mL-1.
Semen preservation at 4°C (cooled preservation): Semen diluted with cooled preservation extender was cooled to 4°C and maintained at that temperature in a refrigerator. Semen samples were assessed for sperm quality (sperm viability, acrosomal integrity, mitochondrial function and capacitation status) every 24 h over 5 days. Semen preservation at 23°C: Semen diluted with cooled preservation extender was maintained at room temperature (23°C) and samples were evaluated for sperm quality (sperm viability, acrosomal integrity, mitochondrial function and capacitation status) after 24 h and 48 h storage.
Cryopreservation: Semen with cryopreservation extender was loaded into 0.25 mL straws (I.V.M., L’Aigle, France). After equilibrating for 3 h at 4°C, straws were put into straw holders and frozen in liquid nitrogen vapor, 2 cm above the LN2 level for 7 min, before being plunged into LN2 for storage. For thawing, frozen straws were removed from liquid nitrogen and put into a 40°C water bath for 10s.
Flow cytometric analysis
Assessment of sperm viability: Five micro-liters of 1.0 mmol L-1 Propidium Iodide (PI) was added to 400 μL PBS diluted semen samples with a concentration of 7.5x106 sperm/400 μL, which then were incubated at 37°C for 15 min after gentle mixing of the sample. Following incubation, sperm were analyzed in the flow cytometer. Sperm emitting a red fluorescence were considered as non-viable. Sperm emitting no fluorescence were considered alive.
Assessment of sperm acrosomal integrity: Five micro liters of 1.03 mmol L-1 fluoresceinated peanut lectin (FITC-PNA) was added to 400 μL Phosphate Buffered Saline (PBS) diluted semen samples with a concentration of 7.5x106 sperm/400 μL and samples were incubated at 37°C for 30 min after gentle mixing of the sample. Following incubation, sperm were analyzed in the flow cytometer. Observed images were classified into two groups: (1) Sperm displaying intensively bright fluorescence of the acrosomal cap (intact acrosome) and (2) Sperm displaying either disrupted fluorescence of the acrosomal cap or no fluorescence (classified as a damaged acrosome).
Assessment of sperm mitochondrial membrane potential (MMP): Four hundreds microlitersof PBS diluted semen samples with the concentration of 7.5x106 sperm/400 μL were pipetted into micro-centrifuge tubes and 5 μL 1.53 mM JC-1 stain in DMSO was added to each sample. After thorough mixing, they were incubated at 37°C for 30 min. The MMP was assessed in a flow cytometer. Sperm emitting a green fluorescence (JC-1 monomers) were assumed to have a low membrane potential and sperm emitting orange/red fluorescence (JC-1 aggregates) a high Mitochondrial Membrane Potential (hMMP) (Martinez-Pastor et al., 2004).
Chlortetracycline fluorescence assessment of sperm: Sperm functional status was evaluated using fluorescence, as described previously by Ward and Storey (1984) and Perez et al. (1996). Two slides from each sample were prepared, and 100 sperm per slide were counted according to the three different CTC staining patterns described by Gillan et al. (1997). The three categories used in the classification were ‘F’ or non-capacitated sperm for those with uniform head fluorescence; ‘B’ or capacitated sperm for those with fluorescence only along the acrosome and ‘AR’ for acrosome-reacted sperm with dull fluorescence over the head.
Flow cytometry analysis of sperm: Information on >10, 000 events was collected using FACSC alibur in list mode on a FACS Analyzer flow cytometer (Becton-Dickinson, San Jose, CA, USA). Data were analyzed with CellQuest (BD software) using User Manual procedures. The flow cytometer was equipped with (1) a standard FITC and phycoerythrin dichroic filter set (2) LP 400 long-pass and DF 485/22 band-pass excitation filters at 485 nm; 3) a DM-560 dichromic mirror to separate fluorescent signals; 4) photomultiplier tube 1 (FL1) collecting 530 nm light through a DF 530/30 band-pass filter and 5) FL2 collecting light through an LP 570 filter. Compensation was used to minimize spillover of green fluorescence into the red channels.
Artificial insemination: Ewes were furnished by a Beijing Breed Selection Centre. A total of 123 adult females with established fertility were selected according to their age 2-4 years old, favorable body condition and reproductive history. All inseminated females were multiparous or at least had one previous parity.
The aim of this experiment was to compare the fertilizing ability of semen stored at 23, 4 or -196°C. To this end, each ejaculate was divided into equal numbers of artificial insemination doses stored at the three temperatures. Ewes were randomly assigned to five experimental groups which were inseminated with fresh semen (n = 28), semen stored at 23°C for 1 day (n = 25), semen stored at 4°C for 3d (n = 23), semen stored at 4°C for 4d (n = 23) or cryopreserved semen (n = 24). Pregnancy status and birth rate were assessed.
Estrus was synchronised by insertion on day 0 of a progestagen sponge (45 mg of fluorogestone acetate, FGA, Chronogest 1, Intervet Pty Ltd., Australia) at unknown stages of estrous. On day 13 the sponges were removed and the ewes were treated once with PMSG (Ningbo Hormone Products Co., Ltd., China) 260-280 IU per ewe, Within 24 h of sponge removal, a teaser ram equipped with an apron was penned with the ewes.
Semen was thawed for 10 s in a 40°C water bath just before insemination. Using a speculum with an attached light source and ovine-caprine AI catheter, each ewe was inseminated intracervically at 8-12 h and at 16-24 h (5x107∼9x107 sperm/one time) after observed estrus, using a dose from the same ejaculate and extender each time. Thawed semen was deposited as deeply as possible in the genital tract.
Pregnancy status was determined by trans-abdominal ultrasound (5.0 MHZ) 45 days after insemination.
Statistic analysis: All experiment was repeated three times. The percentage data were subjected to arcsine transformation before statistical analysis. Data were analyzed by one-way ANOVA applying Duncan’s test. The data were presented as Mean±SEM. A value of p<0.05 was considered statistically significant.
RESULTS
Effect of different preservation methods on the sperm viability of Small-tail Han sheep: As shown in Fig. 1, the percentage of viability did not differ significantly among the sperm stored at 4°C for 48 h, sperm stored at 23°C for 24 h or the fresh samples (p>0.05), all of which, however, were higher (p<0.05) than those of the cryopreserved treatment group. Sperm viability of semen stored at 23°C for 24 h was significantly higher than that of samples stored at 4°C for 72 h (p<0.05).
Effect of different preservation methods on the sperm acrosomal integrity of Small-tail Han sheep: As shown in Fig. 2, sperm stored at 4°C for 72 h had a lower proportion of intact acrosomes than those of fresh sperm or of cryopreserved sperm (p<0.05).
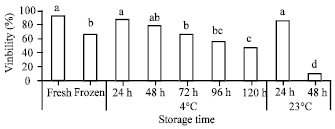 | |
| Fig. 1: | Effect of different preservation methods on the sperm viability of Small-tail Han sheep. Different letters on the top of histograms identify treatments that differ at p<0.05 |
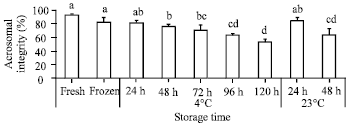 | |
| Fig. 2: | Effect of different preservation methods on the sperm acrosomal integrity (percentage of cells with intact acrosomes) of Small-tail Han sheep. Different letters on the top of histograms identify treatments that differ at p<0.05 |
In addition, the percentage of intact acrosomes did not differ among sperm stored at 23°C for 24 h, fresh sperm and cryopreservated sperm (p>0.05).
Effect of different preservation methods on the sperm Mitochondrial Membrane Potential (MMP) of Small-tail Han sheep: As shown in Fig. 3, sperm held either at 4°C for 72 h or at 23°C for 24 h had a lower percentage of high Mitochondrial Membrane Potential cells (hMMP) than that of the fresh semen group (p<0.05). The percentage of hMMP did not differ between sperm stored at 4°C for 72 h or at 23°C for 24 h (p>0.05). The frequency of sperm with hMMP was lower in cryopreserved semen, compared with semen stored at 4°C for 72 h or at 23°C for 24 h (p<0.05).
Effect of different preservation methods on sperm capacitation status of Small-tail Han sheep: The percentage of incapacitated sperm was lower for semen stored at 4°C for 72 h or at 23°C for 24 h than for fresh semen (p<0.05) and it was higher (p<0.05) than that of the cryopreserved sperm (Fig. 4).
Effect of different preservation methods on the fertility rates of semen after artificial insemination: Pregnancy rates after cervical insemination with semen stored at different temperatures and times are presented in Table 1. No difference was observed between ewes inseminated with semen stored at 4°C for 72 h or stored at 23°C for 24 h or with fresh semen (69.6, 76.0 and 78.6%) but those pregnancy rates were higher than those from using frozen-thawed semen (50.0%, p<0.05).
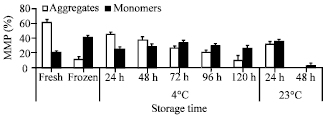 | |
| Fig. 3: | Effect of different preservation methods on the sperm mitochondrial membrane potential (MMP) of Small-tail Han sheep. |
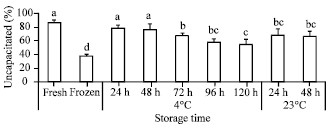 | |
| Fig. 4: | Effect of different preservation methods on sperm capacitation status of Small-tail Han sheep. Different letters on the top of histogram differed significantly (p<0.05) |
| Table 1: | Effect of different preservation methods on fertility rates following artificial insemination |
 | |
| 1Pregnancy rate = No. of ewes diagnosed pregnant 45 days post-insemination/number of ewes inseminatedx100, 2Birth rate = No. of lambs born/number of ewes lambingx100. Values in the same column with different superscripts (a and b) are differ at p<0.05 °C | |
DISCUSSION
In this experiment, sperm viability, acrosomal integrity, sperm MMP (mitochondrial membrane potential) and capacitation status were examined after semen samples were preserved at 23, 4 or -196°C. Viability of semen stored at 4°C for 48 h was not significantly different from that of the fresh semen and retained viability of 55.3% after another 48 h (Fig. 1). At 4°C, spermatozoa metabolism and energy are low, which promotes high motility of sperm cells after warming to 37°C (Paulenz et al., 2000). It is reported that high fertilization rates (75%) have been achieved after intrauterine insemination of ram spermatozoa stored at 5°C for 6 days (Salamon et al., 1979) and such methods have been widely used in swine (Johnson et al., 2000), cattle (Seidel et al., 1999), horse (Lindsey et al., 2002) and goat (Hollinshead et al., 2002). In one experiment, sperm viability was 60% after 10.7 h of storage of Small-tail Han sheep semen at 4°C (Zhou et al., 2002). This result might be attributable to the reduced metabolism of sperm in cooled storage (Salamon and Maxwell, 2000). However, viability of sperm stored at 4°C for 72 or 96 h in our study was significantly lower than that of fresh semen.
In practical production, semen sometimes needs to be preserved for only a short time, and in those circumstances, storage at 23°C may be advantageous to storage at 4°C. There is no cold shock of the semen stored at 23°C. It is reported that motility and the proportion of cells with intact acrosomes were higher in semen stored at 21°C for 24 h than in semen stored at 5°C (Paulenz et al., 2002) As storage time of semen in the liquid state is prolonged, increased concentrations of toxic metabolic products (Griggers et al., 2001) such as ROS (Reactive Oxygen Species) (Vishwanath and Shannon, 1997) may have caused reduced sperm viability. Also in our experiment, semen stored at 23°C for 24 h had better sperm viability than that of semen stored at 4°C for 48 h. This finding is similar to previous work in which sperm stored at 21°C for 24 h had higher motility than sperm stored at 5°C prior to sorting by flow cytometry (Hollinshead et al., 2002). Semen stored at 4°C for 48 h had significantly lower acrosomal integrity than fresh semen. The release of ROS may have been responsible for this loss (Vishwanath and Shannon, 1997) and cold shock may have changed ratios among unsaturated fatty acids and lowered cholesterol content, which have been shown to result in a less stable sperm membrane (White, 1993). This result is in agreement with (Roca et al., 1997) which showed that the percentage of sperm with intact membranes decreased from 94 to 68% when semen was stored at 4°C for 3 days. In this study, the percentage of acrosomally intact sperm in semen held at 23°C for 24 h did not differ from that of fresh semen and was significantly higher than that of the semen stored at 4°C for 72 h. This result could be due to shorter storage time and freedom from cold-shock in semen stored at room temperature (23°C).
Kasai et al. (2002) reported that mitochondrial status played an important role in both motility and fertility of sperm because of its relationship with the energetic status of the cell. As shown in Fig. 3, the percentage of cryopreserved sperm with hMMP (high Mitochondrial Membrane Potential) was significantly lower than that of semen stored at 4°C for 72 h or 23°C for 24 h. This is in agreement with Thomas et al. (1998) in which sperm hMMP rate was 30.9% after storage at 4°C for 24 h, but only 1.3% after freezing. The reason may be that freezing and thawing procedures reduce sperm mitochondrial membrane integrity (Jones and Stewart, 1979; Lindemann et al., 1982), leading to increased permeability to ions and decreased production of ATP (Lindemann et al., 1982) Sperm stored at 4°C for 72 h had a significantly lower proportion of hMMP than fresh semen. Previous reports indicated that during semen storage at 4°C, mitochondria appear in a condensed configuration because of cold shock (Jones and Martin, 1973; Simpson et al., 1986) which is indicative of low cellular energy status (Hackenbrock, 1972).
In the present study, semen stored at 4 or 23°C for any duration and semen stored at -196°C had a significantly lower proportion of incapacitated sperm than fresh semen. In addition, the proportion of incapacitated sperm was significantly lower in cryopreserved semen, compared with semen stored at either 4°C for 72 h or 23°C for 24 h. It has been reported that cryopreservation causes a proportion of the surviving sperm to undergo capacitation-like changes (Cormier and Bailey, 2003). Our results suggest that during semen storage at 4°C or at room temperature (23°C), sperm also undergo capacitation-like changes, but to a lesser extent than in cryopreserved sperm. Pregnancy rate following insemination either with semen stored at 23°C for 24 h or at 4°C for 72 h did not differ from that of fresh semen and was significantly higher than that with cryopreserved semen. It has been reported that freezing has detrimental effects on sperm ultrastructure (mitochondrial, acrosomal and DNA integrity) which reduces survival time of sperm cells in the female reproductive tract (Lightfoot and Salamon, 1970), thereby reducing reproductive success. In this experiment, pregnancy rates following artificial insemination were consistent with our assessment of sperm quality of semen stored at various temperatures and times. Sperm viability, hMMP and uncapacitation rates of the semen stored at 23°C for 24 h all were better than those of frozen semen and pregnancy rate from the former was significantly higher than that of the latter as well. Although, sperm hMMP and incapacitation rates of semen stored at 23°C for 24 h or at 4°C for 72 h (Fig. 3, 4) were lower than that of fresh group, the pregnancy rates were not significantly different for semen stored at 23°C for 24 h, 4°C for 72 h or fresh samples. Apparently, the number of fertile sperm in semen held at 23°C for 72 h or 4°C for 24 h could meet the requirement for normal fertilization in AI.
CONCLUSION
To sum up, semen stored either at 4°C for 72 h or 23°C for 24 h had the same efficacy for AI as that of fresh semen and both were superior to that of semen cry preserved at -196°C. This data suggest that store sheep semen at 4°C and room temperature (23°C) can be an alternative method to cryopreservation. These results can also provide valuable information to other farm animals and wild animals.
ACKNOWLEDGMENTS
The authors thank BillHohenboken for revisingthe English version of this manuscript. This work was supported by Beijing Municipal Science and Technology Commission and Beijing Municipal Agriculture Technology Committee Foundation (No. 20060602) and National Key Technology Research and Development Program Foundation (No.2006BAD14B08).
REFERENCES
- Blash, S., D. Melican and W. Gavin, 2000. Cryopreservation of epididymal sperm obtained at necropsy from goats. Theriogenology, 54: 899-905.
CrossRefPubMedDirect Link - Cormier, N. and J.L. Bailey, 2003. A differential mechanism is involved during heparin- and cryopreservation-induced capacitation of bovine spermatozoa. Biol. Reprod., 69: 177-185.
CrossRef - Gillan, L., G. Evans and W.M. Maxwell, 1997. Capacitation status and fertility of fresh and frozen-thawed ram spermatozoa. Reprod. Fertil. Dev., 9: 481-487.
PubMed - Gravance, C.G., C. White, K.R. Robertson, Z.J, Champion and P.J. Casey, 1997. The effects of cryopreservation on the morphometric dimensions of caprine sperm heads. Anim. Reprod. Sci., 49: 37-43.
PubMed - Griggers, S., D.L. Paccamonti, R.A. Thompson and B.E. Eilts, 2001. The effects of pH, osmlarity and urine contamination on equine spermatozoal motility. Theriogenology, 56: 613-622.
PubMed - Hackenbrock, C.R., 1972. Energy linked ultrastructural transformations in isolated liver mitochondria and mitoplasts. J. Cell. Biol., 53: 450-465.
PubMed - Hollinshead, F.K., J.K. O'Brien, W.M.C. Maxwell and G. Evans, 2002. Production of lambs of predetermined sex after the insemination of ewes with low numbers of frozen-thawed sorted X-or Y-chromosome bearing spermatozoa. Reprod. Fertil. Dev., 14: 503-508.
PubMed - Hu, J.H., L.Q. Wang and Q.W. Li, 2002. Study on the temperature curve in semen freezing of the small-tail sheep. J. Northwest Sci-Tech Univ Agri For., 30: 85-88.
Direct Link - Jones, R.C. and I.C.A. Martin, 1973. The effects of dilution, egg yolk and cooling to 5 degrees C on the ultrastructure of ram spermatozoa. J. Reprod. Fertil., 35: 311-320.
PubMed - Jones, R.C. and D.L. Stewart, 1979. The effects of cooling to 5 degrees C and freezing and thawing on the ultrastructure of bull spermatozoa. J. Reprod. Fertil., 56: 233-238.
PubMed - Kasai, T., K. Ogawa, K. Mizuno, S. Nagai and Y. Uchida et al., 2002. Relationship between sperm mitochondrial membrane potential, sperm motility and fertility potential. Asian. J. Androl., 4: 97-103.
PubMed - Lightfoot, R.J. and S. Salamon, 1970. Fertility of ram spermatozoa frozen by pellet method. I. Transport and viability of spermatozoa within the genital tract of the ewe. J. Reprod. Fertil., 22: 385-398.
PubMed - Lindemann, C.B., M. Fisher and M. Lipton, 1982. A comparative study of the effects of freezing and frozen storage on intact and demembranated bull spermatozoa. Cryobiology, 19: 20-28.
PubMed - Lindsey, A.C., J.L. Schenk, J. K.Graham, J.E. Bruemmer and E.L. Squires, 2002. Hysteroscopic insemination of low numbers of flow sorted fresh and frozen thawed stallion spermatozoa. Equine. Vet. J., 34: 121-127.
CrossRef - Maxwell, W.M. and L.A. Johnson, 1999. Physiology of spermatozoa at high dilution rates: The influence of seminal plasma. Theriogenology, 52: 1353-1362.
CrossRefPubMedDirect Link - Marti, E., J.I. Marti, T. Muino-Blanco and J.A. Cebrian-Perez, 2008. Effect of the cryopreservation process on the activity and immunolocalization of antioxidant enzymes in ram spermatozoa. J. Androl., 29: 459-467.
PubMedDirect Link - Martinez-Pastor, F., A. Johannisson, J. Gil, M. Kaabi, L. Anel, P. Paz and H. Rodriguez-Martinez, 2004. Use of chromatin stability assay, mitochondrial stain JC-1 and fluorometric assessment of plasma membrane to evaluate frozen-thawed ram semen. Anim. Reprod. Sci., 84: 121-133.
CrossRef - Paulenz, H., E. Kommisrud and P.O. Hofmo, 2000. Effect of long-term storage at different temperatures on the quality of liquid boar semen. Reprod. Domestic Anim., 35: 83-87.
CrossRef - Paulenz, H., L. Soderquist, R. Perez-Pe and K.A. Berg, 2002. Effect of different extenders and storage temperatures on sperm viability of liquid ram semen. Theriogenology, 57: 823-836.
Direct Link - Perez, L.J., A. Valcarcel, M.A. de las Heras, D.F. Moses and H. Baldassarre, 1996. In vitro capacitation and induction of acrosomal exocytosis in ram spermatozoa as assessed by the chlortetracycline assay. Theriogenology, 45: 1037-1046.
PubMed - Peris, S.I., A. Morrier, M. Dufour and J.L. Bailey, 2004. Cryopreservation of ram semen facilitates sperm DNA damage: Relationship between sperm andrological parameters and the sperm chromatin structure assay. J. Androl., 25: 224-233.
PubMed - Purdy, P.H., 2006. A review on goat sperm cryopreservation. Small Rumin. Res., 63: 215-225.
CrossRefDirect Link - Roca, J., J.A. Carrizosa, I. Campos, A. Lafuente, J.M. Vazquez and E. Martinez, 1997. Viability and fertility of unwashed Murciano-Granadina goat spermatozoa diluted in tris-egg yolk extender and stored at 5°C. Small Rumin. Res., 25: 147-153.
Direct Link - Salamon, S. and W.M.C. Maxwell, 1995. Frozen storage of ram semen I. Processing, freezing, thawing and fertility after cervical insemination. Anim. Reprod. Sci., 37: 185-249.
CrossRef - Salamon, S. and W.M.C. Maxwell, 1995. Frozen storage of ram semen II. Causes of low fertility after cervical insemination and methods of improvement. Anim. Reprod. Sci., 38: 1-36.
CrossRef - Salamon, S. and W.M.C. Maxwell, 2000. Storage of ram semen. Anim. Reprod. Sci., 62: 77-111.
PubMedDirect Link - Simpson, A.M., M.A. Swan and I.G. White, 1986. Action of phosphatidylcholine in protecting ram sperm from cold shock. Gamete Res., 15: 43-56.
CrossRef - Thomas, C.A., D.L. Garner, J.M. Dejarnette and C.E. Marshall, 1998. Effect of cryopreservation on bovine sperm organelle function and viability as determined by flow cytometry. Bio. Reprod., 58: 786-793.
PubMed - Vishwanath, R. and P. Shannon, 1997. Do sperm cells age? A review of the physiological changes in sperm during storage at ambient temperature. Reprod. Fert. Dev., 9: 321-331.
PubMed - Watson, P.F., 1995. Recent developments and concepts in the cryopreservation of spermatozoa and the assessment of their post-thawing function. Reprod. Fertil. Dev., 7: 871-891.
CrossRefPubMedDirect Link - White, I.G., 1993. Lipids and calcium uptake of sperm in relation to cold shock and preservation: A review. Reprod. Fertil. Dev., 5: 639-658.
PubMed