
Ying He
Hebei Key Laboratory of Preventive Veterinary, Department of Animal Science, Hebei Normal University of Science and Technology, Qinhuangdao 066600, China
Fei Zhong
Department of Basic Veterinary Medicine, College of Animal Science and Technology, Agricultural University of Hebei, Baoding 071001, China
Wangbin Cao
Hebei Key Laboratory of Preventive Veterinary, Department of Animal Science, Hebei Normal University of Science and Technology, Qinhuangdao 066600, China
Manfu Zhang
Laboratory for Animal Molecular Virology, College of Biological Sciences, China Agricultural University, Beijing 100094, China
Asian Journal of Animal and Veterinary Advances
Year: 2012 | Volume: 7 | Issue: 3 | Page No.: 235-242







ABSTRACT
Canine parvovirus type 2 (CPV-2) has emerged as a pathogen of new acute infectious disease in dogs. It affects primarily young animals where it can cause haemorrhagic enteritis and myocarditis. Receptor binding is a key step in the life cycles of all animal viruses. It has been reported that the specific binding to transferrin receptor (TfR) on cell membranes plays an important role in tissue tropism and host ranges of CPV-2. In this study, the entry and the subcellular localization of viral particles and TfR in the cells of FK81 and MDCK were analyzed; the distribution of TfR and infectious ability of CPV-2 were determined in the cells of various primary tissue cultures from dog fetus. We observed complicated temporal patterns of CPV-2 virus particle trafficking during post-incubation with cells of FK81 and MDCK. The virus particles were located in the cytoplasm 4-8 h after inoculation, they appeared in nuclei at 12-16 h of post-incubation, then reappeared in cytoplasm at 24 h again; they distributed all over the cellular organelles after 32 h incubation. The virus and the transferrin were co-located in the perinuclear cytoplasm after 2 h of incubation with the FK81 cells; addition of excessive transferrin decreased the rate of infection of CPV-2, but did not block virus entry. We found that CPV-2 replicated in primary culture cells of the liver, kidney, cardiac muscle, spleen and intestinal epithelia but not of the lung and internalization of transferrin receptor in primary cells was related to the culture tropism of CPV-2. These results confirmed that the TfR is critical for CPV-2 infection with both cell lines and primary tissue cells from fetes.
PDF Abstract XML References Citation
Received: August 02, 2011;
Accepted: November 30, 2011;
Published: January 14, 2012
How to cite this article
Ying He, Fei Zhong, Wangbin Cao and Manfu Zhang, 2012. The Subcellular Localization and the Tissue Tropism of Canine Parvovirus based on the Co-localization of Transferrin Receptors. Asian Journal of Animal and Veterinary Advances, 7: 235-242.
DOI: 10.3923/ajava.2012.235.242
URL: https://scialert.net/abstract/?doi=ajava.2012.235.242
DOI: 10.3923/ajava.2012.235.242
URL: https://scialert.net/abstract/?doi=ajava.2012.235.242
INTRODUCTION
Canine parvovirus (CPV-2) is a member of the autonomous parvovirus genus in the Parvoviridae family, primarily affecting young animals where it can cause haemorrhagic enteritis and myocarditis. The virus is believed to be a host range variant of a feline parvovirus that acquired the ability to infect dogs through changes in its capsid proteins (Hueffer et al., 2003a). CPVs have a single stranded DNA genome of 5 kb in size. The viruses replicate the genome in the host nuclei using the host replication machinery. Replicated genome is transported into plasma where the DNA content is packaged with the capsid proteins. The packaged virions contain the DNA genome and multiple copies of two capsid proteins VP1 and VP2.
Receptor binding is a key step in the infectious cycles of all animal viruses and in many cases the virus-receptor interactions determine both the host susceptibility and tissue tropism (Smith and Helenius, 2004; Spear, 2004). CPV-2 enters into its host cells via transferrin receptors and clathrin-mediated endocytosis (Parker and Parrish, 2000; Parker et al., 2001). Studies of the virus-cell interactions and intracellular trafficking of canine and feline parvoviruses have shown that the specific interactions between viral capsids and host transferrin receptors are critical to infection (Hueffer et al., 2003b, 2004; Palermo et al., 2003, 2006). Goodman et al. (2010) demonstrated further that most substitutions of TfR had little effect on virus binding and uptake except mutations of several sites in the apical domain of the receptor that either prevented binding to the capsids or reduced the affinity of receptor binding to various degrees. Glycans within the virus binding face of the apical domain also contributed to capsid binding. Those studies were conducted in cells of cell lines, the relevance of the observations to the in vivo infection is not clear.
To understand the relationship between CPV entry and its tissue tropism, we examined the viral entry and the subcellular localization of virons and TfR in the cells of FK81 and MDCK and examined the reproduction of CPV-2 in primary culture cells of liver, kidney, cardiac muscle, spleen, lung and intestinal epithelia; we also determined the locations of transferrin receptor in those primary cells. Our results confirmed that the TfR is critical for the infection of CPV-2 in both cell lines and primary fetal tissue cells.
MATERIALS AND METHODS
Virus, plasmids and cells: Feline FK81 cells and canine MDCK cells were purchased from “The Clinical Central of the Agricultural Ministry of China”. Three newborn canines were sacrificed and the heart, liver, kidney, spleen, lung and duodenum were put into PBS on ice under sterile condition. The primary culture cells were prepared as regular methods. All cells were maintained in Dulbecco’s modified Eagle’s medium with 10% fetal calf serum in 5% CO2 at 37°C. The B2004 strain of canine parvovirus was from our laboratory stock, which was isolated from the stool of a sick dog in Beijing (He et al., 2009). Its sequence (accession No. EF011664) indicated that B2004 is a member of the widely distributed CPV-2a subclade (He et al., 2009). The virus was propagated in feline kidney cell line (FK81) and harvested 5 days post infection. Cell debris was pellet by centrifugation and the supernatant containing virions was stored in a -70°C freezer until use. The CPV suspensions were titrated using 50% tissue culture infective dose (TCID50) assay (LaBarre and Lowy, 2001). CPV monoclonal antibodies mAbc2 (against VP2 of virus capsid) was kindly provided by Prof. Sun (The Clinic Central of the Agricultural Ministry, China).
Iron loading and labeling of canine transferrin: Canine transferrin (Sigma, St. Louis, Mo.) was iron loaded and labeled as described by Hueffer et al. (2003a). Briefly, transferrin (5 mg mL-1) was incubated for 30 min at 37°C with a solution containing 240 nM FeCl3, 2 mM nitrilotriacetic acid in 250 mM Tris-HCl (pH 8.0) and 10 μM NaHCO3. Iron-loaded transferrin was separated from free iron by a PD-10 column in PBS. For labeling, 2 mg of iron-loaded transferrin was dialyzed against 50 mM borate buffer (pH 9.0), incubated with 0.1 mg of rhodamine (Dupont NEN, Boston, Mass.) for 1 h at 4°C. The labeled transferrin was separated from free dye by a Sephadex G25 column in PBS.
Infection assays: Cells were seeded on 24 well plate, incubated for 1 h at 37°C, then were inoculated with CPV (103.65 TCID50 per well) and were incubated for 12 h after inoculation. The infected cells were continually incubated at 37°C for 48 h in fresh growth medium. The cells were fixed with 4% paraformaldehyde, permeabilized with 0.1% Triton X-100 in PBS with 1% bovine serum albumin. Infected cells were detected by immunostaining with mAbc2 as primary antibody and goat anti-mice IgG-FITC as secondary antibody. The infection rate was determined by microscopy as the percentage of green-stained cells in total 200 cells counted in each sight field and averaged over five sight fields.
Microscopic analysis of localization of the virus and the transferrin: Cells seeded on cover slip and incubated for 12 h, then were inoculated with unlabeled virus particles (103.65 TCID50 per well) and incubated at 37°C for various time as indicated. The cells were washed and fixed with 4% paraformaldehyde in PBS. For antibody staining, the cells were permeabilized with 0.1% Triton X-100 in PBS and 1% bovine serum albumin before incubation with mAbc2 (specific identified VP2) as primary antibody and FITC conjugated goat anti-mouse IgG as secondary antibody to detect virus. The immunostained cells were examined with a laser scanning fluorescence microscope (LSM510, Zeiss). To examine distribution of canine transferrin in the cells of primary tissue culture or cell lines, the cells seeded on cover slip incubated with labeled transferrin for 2 h, washed in cold PBS or the cells incubated with both labeled transferrin and viral particles for 0.5-2 h, then cells were immunostained and confocal images were obtained using a Zeiss LSM510 confocal microscope and images were analyzed using Zeiss software.
RESULTS
Step sequences of CPV-2a entry and intracellular trafficking were similar for FK81 and MDCK cell lines: The virus particle distribution as detected by mAbc2 (specific to VP2) in cells after inoculation 4-48 h was shown in Fig. 1. The viral uptake and intracellular trafficking occur following certain path (Harbison et al., 2009). Present result demonstrated that, the viral particles were initially attached on the surface of the cells, then entered into the cytoplasm and stayed there for 8-12 h, then transferred into nuclei where the genome was presumably replicated as evidenced that many viral particles appeared in nuclei after incubation for 16 h. After 24 h incubation, viral particles appeared in cytoplasm, in consistence with assembly of new viruses in cytoplasm. The viral particles were distributed all over the cell after 32 h incubation. This temporal pattern of viral distribution was shared by both FK81 and MDCK cell lines.
CPV-2a capsid and TfR were colocolized subcellularly in F81 cell: To determine the interaction between CPV-2a and TfR, the FK81 cells were incubated with CPV particles and rhodamine-labeled transferrin for a time as indicated, then stained by mAbc2 and FITC conjugated goat anti-mouse IgG and examined with a laser scanning fluorescence microscope. Figure 2 showed that the virus and transferrin located in the same compartments in cytoplasm, particularly presented around perinuclear cytoplasm after 2 h incubation.
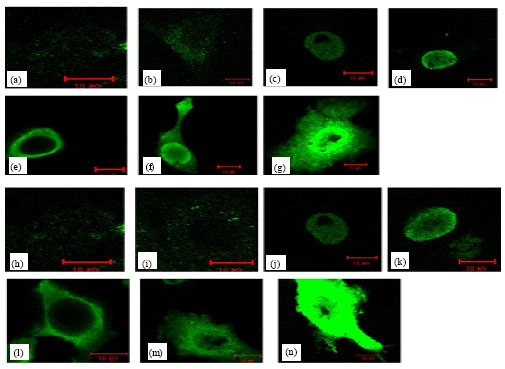 | |
| Fig. 1(a-n): | Confocal images of cultured cells after inoculation with CPV at times (h) as indicated in 10 μm scale. The green shows the CPV particles stained by CPV Mab and FITC conjugated goat anti-mouse IgG. (a-g) Images of MDCK cells at 4, 8, 12, 16, 24, 32, 48 h and (f-m) Images of FK81 cells at 4, 8, 12, 16, 24, 32, 48 h |
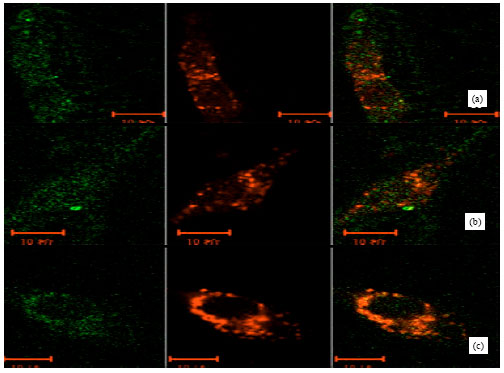 | |
| Fig. 2(a-c): | Confocal images of FK81 cell incubated with both CPV particles and transferrin for times as indicated in 10 μm scale. The red shows the Tf labeled with rhodamine; the green shows the CPV particles stained by CPV Mab and FITC conjugated goat anti-mouse IgG. (a) 0.5 hm (b) 1 h and (c) 2 h |
| Table 1: | Infection percentage of different tissue cultures inoculated with CPV after 48 h |
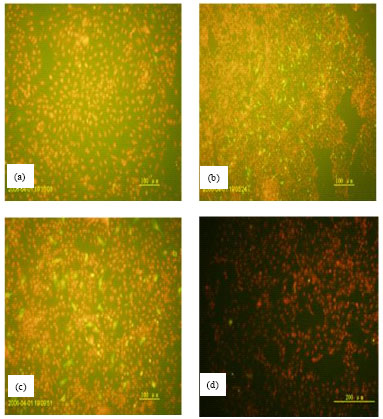 | |
| Fig. 3(a-d): | Detection of the MDCK cell line infected by CPV 48 h after inoculation (100 μm scale) treated with (a) CPV antibody, (b) Nothing, (c) Transferrin and (d) Negative control (Normal cell). Red shows the nuclear DNA of the cells stained by PI, green shows the CPV particles immuno-stained by CPV Mab and FITC conjugated anti-mouse IgG |
This result suggested that for entering into cells, CPV particles and transferrin molecules were likely bound to the same receptors that were TfR. Addition of excessive transferrin decreased but not blocked the infection of CPV. In contrast, viral infection and reproduction were interrupted by adding of mAbc2 (1:100 dilution) against CPV (Fig. 3).
Susceptibilities to CPV-2a were different for various primary tissue culture cells: To test the culture tropism of CPV-2a, the primary culture cells of liver, kidney, cardiac muscle, lung, spleen and intestinal epithelia were infected with virus, the result showed that CPV reproduced in those cells except lung cell and the infection rates varied for cell origins (Table 1). To our knowledge, this is the first study to demonstrate the susceptibilities of primary cells of dog to CPV.
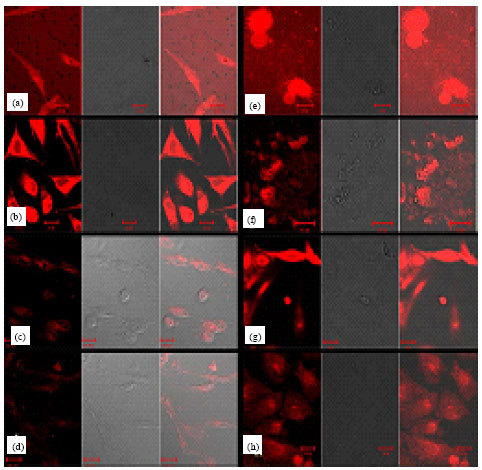 | |
| Fig. 4(a-h): | Confocal images of four category cells incubated with rhodamine-labeled transferrin in 20 μm scale. (a) Cardiac cell, (b) Nephritic cell (c) Lung cell, (d) MDCK cell, (e) Spleen cell, (f) Intestinal epithelia, (g) FK81 cell and (h) Hepatic cell |
The presence of transferrin receptor in primary cells was relative to the tissue culture tropism of CPV: To determine the role of the TfR in tissue tropism of virus, primary culture cells of various tissues were incubated with rhodamine-Tf for 2 h, examined with a laser scanning fluorescence microscope. Substantial amount of TfR was found in tissue culture cells of kidney, cardiac muscle, spleen and intestinal epithelia but little in lung cells as shown in Fig. 4, in accordance with the susceptibility assay in 3.3. This result demonstrated for the first time that the TfR was critical to viral infection of primary tissue cells.
DISCUSSION
Viral infection of animal cells involves sequential steps that deliver the virus and its genome from the cell surface to the compartment where replication can occur. The binding to TfRs on the surface of host cell was the crucial step for CPV infection. TfR binding appears to provide a structural interaction necessary for infection as replacement of the TfR ectodomain with binding domains of antiviral antibodies allowed attachment and uptake of virus but not infection (Hueffer et al., 2003a, b). We showed that after incubation with FK81 or MDCK cell for 2 h, both the CPV particles and TfR were located in the cytoplasm surrounding nuclei and that there is close relationship between abundance of TfRs in primary cells and the ability of CPV to infect those cells. This is the first study to elucidate the mechanism of CPV tissue tropism to primary tissue culture based on the TfR localization. Our results showed that the CPV and Tf likely used same receptor (TfR) to enter the cell and the presence of transferrin receptor in primary cells was relative to the tissue culture tropism of CPV.
In the normal pathway of TfR-mediated entry, the TfR-Tf complex enters the early endosome, from which some of the complex is rapidly recycled to the cell surface while most passes to the perinuclear recycling endosome. From there it recycles to the cell surface where the iron-free apo-Tf is released at neutral pH. The rate of uptake and the efficiency of TfR recycling depend on the form of the ligand and more than 97% of monomeric Tf recycles to the cell surface within 10 to 30 min (Ponka and Lok, 1999; Van Dam and Stoorvogel, 2002). However, cross-linking TfRs with oligomeric Tf or antibodies causes the complexes to be retained within endosomes for longer time and a higher proportion is trafficked to late endosomes and lysosomes for degradation (Marsh and Helenius, 2006; Govindasamy et al., 2003). Present result reveals that addition of Tf decreases the viral infection but did not block the entry.
TfR expression is tightly regulated and it is highly expressed in dividing cells with high iron needs. High expression of TfR would favor the binding of viruses. The presence of TfR in various tissue cells should be relevant to the entrance and reproduction of virus. Present results indicated that this is indeed to be the case and we demonstrated that TfR played importance role in infection of CPV. Our result showed that CPV reproduced in the primary culture cells of liver, kidney, cardiac muscle, spleen and intestinal epithelia except lung cell and the infection rates of those cells are correlate to the distribution of TfR on the cells. It may clue to some information on the tissue tropism in natural cases, which answer for haemorrhagic enteritis and myocarditis of puppy. The results provided a foundation for further biophysical studies and for antiviral drug search and design based on the TfR.
ACKNOWLEDGMENTS
The study was supported by the PhD Grand of Hebei Normal University of Science and Technology. The authors would like to thank Prof. Sun for his excellent assistance, kindly providing us with the anti-CPV Mab used in the study. We would also like to thank Prof. Jianying Wang (Inner Mongolia University of Science and Technology) for critically reading the manuscript.
REFERENCES
- Govindasamy, L., K. Hueffer, C.R. Parrish and M. Agbandje-McKenna, 2003. Structures of host range-controlling regions of the capsids of canine and feline parvoviruses and mutants. J. Virol., 77: 12211-12221.
CrossRefDirect Link - Hueffer, K., J.S.L. Parker, W.S. Weichert, R.E. Geisel, J.Y. Sgro and C.R. Parrish, 2003. The natural host range shift and subsequent evolution of canine parvovirus resulted from virus-specific binding to the canine transferrin receptor. J. Virol., 77: 1718-1726.
CrossRefDirect Link - Hueffer, K., L. Govindasamy, M. Agbandje-McKenna and C.R. Parrish, 2003. Combinations of two capsid regions controlling canine host range determine canine transferrin receptor binding by canine and feline Parvoviruses. J. Virol., 77: 10099-10105.
CrossRefDirect Link - Hueffer, K., L.M. Palermo and C.R. Parrish, 2004. Parvovirus infection of cells by using variants of the feline transferrin receptor altering clathrin-mediated endocytosis, membrane domain localization and capsid-binding domains. J. Virol., 78: 5601-5611.
CrossRefDirect Link - Palermo, L.M., K. Hueffer and C.R. Parrish, 2003. Residues in the apical domain of the feline and canine transferrin receptors control host-specific binding and cell infection of canine and feline Parvoviruses. J. Virol., 77: 8915-8923.
PubMedDirect Link - Palermo, L.M., S.L. Hafenstein and C.R. Parrish, 2006. Purified feline and canine transferrin receptors reveal complex interactions with the capsids of canine and feline parvoviruses that correspond to their host ranges. J. Virol., 80: 8482-8492.
PubMed - Parker, J.S.L. and C.R. Parrish, 2000. Cellular uptake and infection by canine parvovirus involves rapid dynamin-regulated clathrin-mediated endocytosis, followed by slower intracellular trafficking. J. Virol., 74: 1919-1930.
Direct Link