
Yiming Sulaiman
College of Animal Science and Technology, China Agricultural University, Beijing 100193, China
Changxin Wu
College of Animal Science and Technology, China Agricultural University, Beijing 100193, China
Chunjiang Zhao
College of Animal Science and Technology, China Agricultural University, Beijing 100193, China
Asian Journal of Animal and Veterinary Advances
Year: 2011 | Volume: 6 | Issue: 1 | Page No.: 71-79







ABSTRACT
To study genetic information of the local sheep populations of Northwestern China and their phylogenetic positions in relation to other Asian populations and European sheep, the genetic diversity and phylogeny evolution of 223 individuals from 19 indigenous sheep populations in Northwestern China and one Western breed, Texel were analyzed with entire sequences of mitochondria DNA control region. The length of the sequences was considerably variable between 1103 and 1256 bp. Phylogenetic analysis indicated that 3 mtDNA lineages A, B and C were found in all sampled populations, except for the absence of lineage C in two populations. Phylogenetic tree and median joining network were constructed with the sequenced mtDNA control region and both the phylogenetic tree and network demonstrated that there were three independent clusters and some sequences deviated from cluster A and C. Neighbor-joining tree constructed based on genetic distances showed a multi-maternal originality of the nineteen Xinjiang local sheep populations, which were clustered with the sheep groups in other parts of China, Central Asia, Turkey and India. The high genetic diversity of Xinjiang local sheep populations suggests that they may have distinct maternal origins and more complicated phylogenetic history than the other Chinese sheep breeds.
PDF Abstract XML References Citation
Received: March 28, 2010;
Accepted: May 06, 2010;
Published: July 10, 2010
How to cite this article
Yiming Sulaiman, Changxin Wu and Chunjiang Zhao, 2011. Phylogeny of 19 Indigenous Sheep Populations in Northwestern China Inferred from Mitochondrial DNA Control Region. Asian Journal of Animal and Veterinary Advances, 6: 71-79.
DOI: 10.3923/ajava.2011.71.79
URL: https://scialert.net/abstract/?doi=ajava.2011.71.79
DOI: 10.3923/ajava.2011.71.79
URL: https://scialert.net/abstract/?doi=ajava.2011.71.79
INTRODUCTION
Mitochondrial DNA (mtDNA) is an important material for phylogenetic inference and has been widely used to explore genetic diversities among various groups of organisms. MtDNA data from wild and domestic sheep revealed that there were no contributions from urial and argali species to domestic sheep (Hindleder et al., 1998, 2002) and 3 maternal lineages A, B and C were identified in modern domestic sheep breeds sampled from different geographical regions of the world (Hiendleder et al., 1998, 2002; Meadows et al., 2007; Wood and Phua, 1996; Guo et al., 2005; Pedrosa et al., 2005; Tsunada et al., 2010).
Xinjiang Uyghur Autonomous Region in the northwestern China spanning over 1.6 million km2, which constitutes one-sixth of China's total territory, as a hub of Silk Road, playing an important role to linking Middle East and Europe and still preserves a multi-ethnic, multi-linguistic and multi-religious features up to now. The area is a melting pot for not only different cultures, but also livestock breeds. The Uyghur, Kazakh, Kyrgyz, Mongolian, Tajik and the other ethnic groups in this region all have long nomadic history and they raised and preserved their livestock breeds under different cultures, geographical isolation and self-sufficient production systems. Up to now, Xinjiang still has a large number of indigenous sheep populations.
The numbers of sheep breeds in Xinjiang are different in different sources (ACS, 1964; Yu et al., 1992; Tu et al., 1989; Chen and Xu, 2004). Some sheep breeds still have not been registered officially due to the lack of genetic information and study. Genetic information about local sheep breeds of Xinjiang and their varieties and its phylogenetic position in relation to neighbor Central Asian Republics and the other areas of China have not been clearly demonstrated.
The aim of this research is to study the genetic diversity and the phylogeny of indigenous sheep populations domesticated in Xinjiang. In the present study, we compared our data with the available sheep sequences in Genbank database (Guo et al., 2005; Hiendleder, 1998; Hiendleder et al., 1998, 2002; Pedrosa et al., 2005) and solid evidences achieved from the mtDNA analysis showed that the indigenous sheep populations in Xinjiang have multi-maternal origins.
MATERIALS AND METHODS
Sample collection and genomic DNA extraction: This research project was conducted from October 2008 to December 2009 in Beijing, including sampling from Xinjiang local sheep from June 2008 to October 2008 and January 2009 to March 2009.
Ear tissues of nineteen sheep populations were collected from small and remote villages in Xinjiang and blood samples of Texel, a European breed, were collected from a farm in Beijing. Samples were randomly collected from each population and the geographical distribution and the pedigree information provided by the owners and local farmers were also taken into consideration. Ear tissues were collected and stored at -20°C in 75% ethanol before DNA extraction. Total Genomic DNA was extracted from ear tissue and blood by a standard phenol-chloroform extraction method with minor modification (Sambrook et al., 1989). Geographical distribution of the sampled populations was shown in Fig. 1.
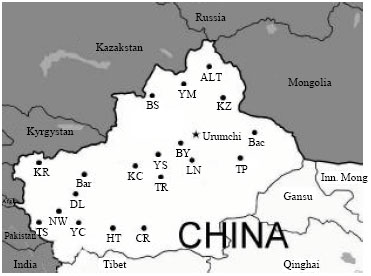 | |
| Fig. 1: | Geographical distribution of the nineteen studied sheep populations in Xinjiang. Full names of the populations were given in Table 2. The map was copied from Wikipedia (http://www.en.wikipedia.org/wiki/File:China_Xinjiang.svg) with modification |
DNA amplification and sequencing: To select individuals from each population for mtDNA sequencing, the two pairs of primers and SSCP analysis described previously were used (Guo et al., 2005). The PCR amplification for SSCP analysis was carried out in a 25 μL reaction mixture containing 20-100 ng DNA template, 1.0 μM of each primer, 200 μmol of dNTPs, 1x PCR buffer (including 1.5 mM MgCl2 and one unit of Taq DNA polymerase (Huitian Dongfang Co., Beijing, China). The PCR conditions consisted of an initial denaturation at 94°C for 5 min, followed by 33 cycles at 94°C for 30 sec, 58°C for 30 sec, 72°C for 45 sec and a final extension of 10 min at 72°C. The PCR Amplification was performed in an Effondorf Master Gradient Programmable Thermal Controller (Effondorf Inc., Germany).
The SSCP vertical electrophoresis was performed at 10V cm-1 in 7 min at first and following 5V cm-1 at RT (room temperature) for 14 h, using the DYY-6C power system (Beijing 61 Factory, Beijing, China). The gel was silver stained and different lineages visually determined and scored (Guo et al., 2005). The DNA Samples for sequencing entire mitochondrial control region were randomly selected from each lineage and amplified with the primers described previously (Hiendleder et al., 2002). The purified products were bidirectly sequenced using BigDye™ Terminator v3.1 Ready Reaction Cycle Sequencing Kit (Applied Biosystems) on ABI PRISM 3700 DNA Analyzer. Two following internal primers were used for sequencing in present study.
SGM-C378 (5’-ATGCGTATCCTGTCCATTAGA-3’) and SGM-C379 (5’-TGAAGAAAGAAC CAGATGCC-3’).
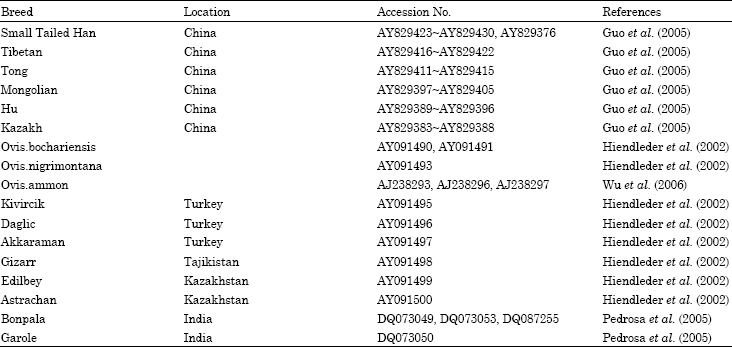
Data analysis: Data analyzed in the present study comprised the sequences of entire mtDNA control region of 223 from the nineteen sheep populations in Xinjiang and one western breed, Texel and other 59 known sheep sequences obtained from Genbank, including the breeds in China, Turkey, Tajikistan, Kazakhstan, India and wild sheep like Ovis ammon nigrimontana, Ovis vignei bochariensis, Ovis ammon (argali). The breed names of sequences retrieved from Genbank, their location and Genbank access number are shown in Table 1. All sequences were manually edited using the Bioedit package (Hall, 1999) and aligned by using the Clustal W program (Thompson et al., 1994).
| Table 1: | Sequences obtained from Genbank |
 | |
Phylogenetic and molecular evolutionary analyses were conducted using MEGA version 4 (Tamura et al., 2007), with a Kimura 2-parameter (transition only) model and a bootstrap (1000 replications) test. A pairwise distance matrix between mtDNA haplotypes was calculated and a Neighbor Joining (NJ) tree was constructed based on the nucleotide p-distances and other parameters were set at the default values.
The Median Joining (MJ) networks (Bandelt et al., 1999) were drawn using the program Network 4.1.0.9 (http://www.fluxus-engineering.com) to investigate the possible relationships among haplotypes. Nucleotide and haplotype diversities were computed using DnaSP 5.10.00 (Librado and Rozas, 2009). Fu’s Fs statistics (Fu, 1997) and Mismatch distribution, were computed using Arlequin3.11 (http://www.cmpg.unibe.ch/software/arlequin3/).
RESULTS AND DISCUSSION
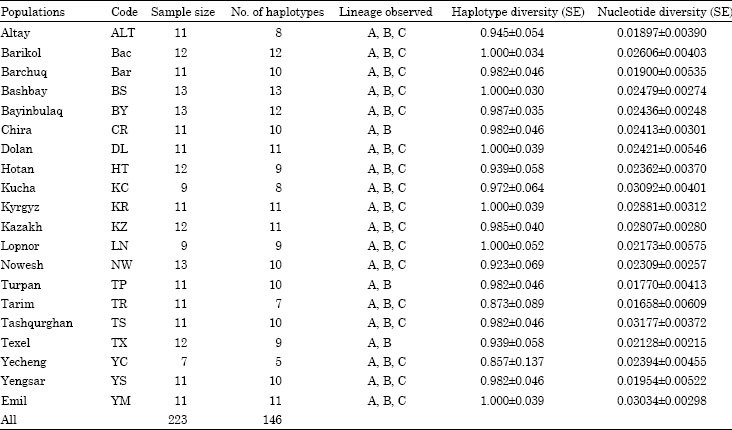
Two hundred and twenty three mtDNA samples from the nineteen local breeds in Xinjiang and one Western breed were sequenced. The length of the sequences in Xinjiang sheep was considerably variable between 1103 and 1256 bp. The sample information and number of haplotypes, haplotypes diversity, observed lineage and nucleotide diversity of each population were shown in Table 2.
We constructed a Neighbor-joining phylogenetic tree (Saitou and Nei, 1987) of the 19 Xinjiang local populations with Hu, Tong, Han, Mongolian, Tibetan, Kivircik, Daglic, Akkaraman, Gizarr, Edilbey, Astrachan, Bonpala, Garole, Ovis ammon nigrimontana, Ovis vignei bochariensis and Ovis ammon argali which retrieved from Genbank. The results of the phylogenetic analysis showed the same three clades, which were reconciled with haplotypes A, B and C. The NJ tree constructed by Mega 4.0 was shown in Fig. 2.
| Table 2: | The sample information and some diversity indices of the studied population |
 | |
| SE: Standard error | |
 | |
| Fig. 2: | Phylogenetic relationships of Xinjiang local sheep breeds. Sequences of Hu, Tong, Han, Mongolian, Tibetan, Kivircik, Daglic, Akkaraman, Gizarr, Edilbey, Astrachan, Bonpala, Garole, Ovis ammon nigrimontana, Ovis vignei bochariensis and Ovis ammon argali were retrieved from Genbank |
All of the sheep populations in Xinjiang were clustered with other sheep breeds of China, such as Mongolian Sheep, Tibetan Sheep and also the breeds in other countries mainly around Silk Road, such as Kivircik, Daglic, Akkaraman, Gizarr, Edilbey, Astrachan, Bonpala and Garole, which correspond with their geographical distribution. But surprisingly, the Tarim population was not clustered with the sheep breeds in Xinjiang, but with Gizarr, Astrachan, Bonpala and Garole;The Emil population was clustered with Tibetan Sheep at first and then clustered with Edilbey, Daglic, Kivircik, Daglic and Akkaraman, despite of long geographical distances between them. Fu’s Fs statistics (Fu, 1997) gives a very significant negative value when a population expansion occurs. Based on the results of the Fu’s Fs statistic and mismatch distributions (data not shown) from Arlequin, the probability of observing a random neutral sample with a number of alleles is similar than the observed value. The Fs values for haplotypes A, B and C were -76.282 (p<0.000), -22.510 (p<0.000) and -9.334 (p<0.00600), respectively, which suggested that lineages A, B and C depart significantly from the neutral model and have a quite different demographical history. A network analysis was performed to define the genetic structure between the lineages. The results showed that the 3 clusters correspond with the phylogenetic clades (Fig. 3).
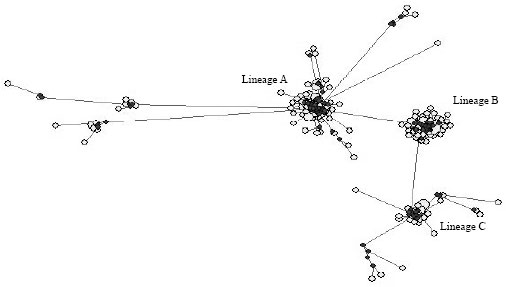 | |
| Fig. 3: | Network of three lineages in nineteen sheep populations in Xinjiang. The hollow circles are 223 samples and the solid ones indicate median vectors |
| Table 3: | Parameters of mtDNA variability within the three clusters in Xinjiang local sheep |
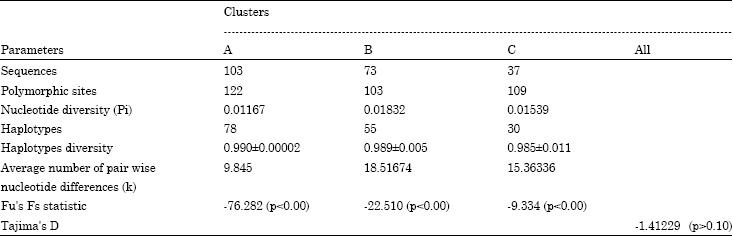 | |
Each clade was clearly isolated, but there was some deviation in clade A and clade C, showing more than one common maternal ancestor in the two clades. Parameters of mtDNA variability within each cluster were given in Table 3.
To study the origins of Xinjiang indigenous sheep populations, 223 entire D-loop regions of different Xinjiang sheep populations were sequenced and a phylogeny tree was constructed with the sequence data. We conducted a comprehensive phylogenetic analysis of the nineteen Xinjiang local sheep breeds to obtain information about the genetic origins and evolutionary history compared with other sheep breeds/populations of different regions of China and other countries.
The length of the sequences varies between 1103 and 1256 bp. The difference of length of the sequences was caused by 3-5 copies of a 75 bp tandem repeated sequence. In earlier studies, three maternal lineages A (Asian type), B (European type) and C (Central Asian and Middle East type) were identified in modern domestic sheep breeds sampled from different geographical regions of the world (Hiendleder et al., 1998, 2002; Meadows et al., 2007; Wood and Phua, 1996; Guo et al., 2005; Pedrosa et al., 2005; Tsunada et al., 2010; Chang, 2009) and a small number of individuals of maternal lineage D (Caucasian type) also were reported before. In the present study, all of lineage A, B and C were found in our studied populations. There were some studies on phylogenetic analysis carried out about the local sheep breeds of Xinjiang (Wang et al., 2007; Yang and Zhao, 2002; Tang et al., 2009; Tsunada et al., 2010; Guo et al., 2005) and the results of them suggested that Xinjiang sheep populations are mainly maternally originated from and influenced by Kazakh and Mongolian sheep. However, there were few studies about the origins and phylogeny of the many unregistered Xinjiang sheep populations. Sheep breeds in China are mainly categorized into Mongolian, Tibetan and Kazakh and Yunnan sheep groups based on archaeological, morphological, historical and available molecular genetic information (Chang, 2009). Previous studies showed that some Xinjiang local sheep populations share the same haplotypes with other sheep breeds of China and local sheep populations in Xinjiang belong to Kazakh sheep groups and have close genetic relationship with Mongolian Sheep (Yu et al., 1992; Tu et al., 1989; Chen and Xu, 2004; Chang, 2009), but in present study, there are some exceptional cases. It is very interesting that the Tarim population was not clustered with other sheep populations or breeds in China, but with Gizarr, Astrachan, Bonpala and Garole; The Emil population was clustered with Tibetan at first and then clustered with Edilbey, Daglic, Kivircik, Daglic and Akkaraman, despite the thousands kilometer distance between them. Some individuals are deviated from clade A and C. Unlike lineage B(European type), lineage A and C had more complex phylogenetic branches, which indicates the two lineages might be derived from a number of founders instead of a single common ancestor. Xinjiang local sheep breeds may have more complicated phylogenetic history than other Chinese breeds. The populations like Tarim, Yengsar, Barchuq and Lopnor shows less genetic distance from Kazakh and Mongolian sheep breeds and were clustered with Tajikistan and Indian sheep’s.
Most of Xinjiang sheep populations were closed to Asian types. However, one population showed genetic links with the Turkish and Indian sheep breeds. The samples used in the study were mostly distributed in remote and undeveloped areas and hardly influenced by foreign sheep breeds, so they could represent the genetic situations of these domestic sheep populations. But due to the limited sequencing number of each population, it could not be confirmed whether the populations had migrated from Turkish and Indian sheep breeds or Turkish and Indian sheep breeds originated from Tarim.
In the present study, some sheep populations from the same region were clustered into different subgroups, like Tarim, Lopnor and Kucha which were from South Xinjiang, but our results showed that they belonged to different subgroups, respectively. On the other hand, some populations from different areas were clustered to the same subgroups which suggested that there might be some gene flows between sheep populations. Although, this region was separated by mountains, large deserts, relatively isolated oasis and long geographical distances, gene flows probably existed between the different populations in these regions along with Silk Road.
Historical and archaeological events indicated that Europoid people lived in Xinjiang from pre-historic times. In ancient time, Xinjiang territory was usually divided by many local oasis rulers like Huns, Turkic and Mongoloid or warring empires 'chasing grass and water' (Millward, 2007). Xinjiang were sweep away by imperators ruling over pastoralist and agrarian areas from Iran to China. Parts of Xinjiang were often ruled together with lands which now are now the Central Asian Republics. Later the dissolution of Turk empire because of wars, diseases and natural disaster led to one of history's great movement of Turkic-speaking tribes into Xinjiang and across Central Eurasia. As a result of the movement, there are now several Turkic language-spoken ethnic groups in Xinjiang including Turkish, Uyghur, Uzbek, Kazakh, Kyrgyz and others. The movement also led to migration and integrations of livestock populations. For thousands of years, there were active commodities exchanges along the famous Silk Road. Sheep, as a main livestock in these regions, might also be under the exchange. The above information and historical events may provide some clues for us to understand why the two Xinjiang local sheep populations were genetically closed to Turkish and Indian sheep breeds.
Present results show that genetic relationships of the two populations with Mongolian, Tibetan and Kazakh breeds are not close and the origin and migration history of Tarim and Hotan populations in South Xinjiang were independent from the other populations in the same areas. The high genetic diversity of Xinjiang local sheep populations implies that they may have distinctive origins. According to NJ phylogenetic tree and genetic distances, Xinjiang local sheep populations may be classified into four subgroups: (1) Altay, Turpan,Tarim, Yengsar, Barchuq and Lopnor. (2) Emil, Chira, Barikol, Bashbay, Tashqurghan, Kyrgyz, Nowesh, Bayinbulaq and Kazakh. (3) Yecheng, Kucha and Hotan. (4) Dolan. The first and third subgroups might be derived from a number of founders instead of a common ancestor.
This study provided molecular evidences of Xinjiang local sheep of genetic diversities, which showed that Xinjiang local sheep populations may have multiple maternal origins, which are not only related to Kazak, Mongolian and Tibetan origin, but also to other two ancestors of Hotan and Tarim.
The results would be also helpful for us to understand the diversity and origins of Central Asian and Middle East domestic sheep. Whether Hotan and Tarim are derived from two separate maternal origins needs to enunciate with more evidences from the further studies on phylogeny of sheep populations in Afghanistan, Pakistan, Iran and Central Asian Republics.
ACKNOWLEDGMENTS
The research was supported by the National Basic Research Program of China (973 Project) (code: 2006CB102100).
REFERENCES
- Bandelt, H.J., P. Forster and A. Rohl, 1999. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol., 16: 37-48.
PubMedDirect Link - Fu, Y.X., 1997. Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics, 147: 915-925.
PubMedDirect Link - Guo, J., L.X. Du, Y.H. Ma, W.J. Guan and H.B. Li et al., 2005. A novel maternal lineage revealed in sheep (Ovis aries). Anim. Genet., 36: 331-336.
PubMed - Hall, T.A., 1999. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acid Symp. Ser., 41: 95-98.
Direct Link - Hiendleder, S., 1998. A low rate of replacement substitutions in two major Ovis aries mitochondrial genomes. Anim. Genet., 29: 116-122.
PubMed - Hiendleder, S., K. Mainz, Y. Plante and H. Lewalski, 1998. Analysis of mitochondrial DNA indicates that domestic sheep are derived from two different ancestral maternal sources: No evidence for contributions from urial and argali sheep. J. Heredity, 89: 113-120.
CrossRefDirect Link - Hiendleder, S., B. Kaupe, R. Wassmuth and A. Janke, 2002. Molecular analysis of wild and domestic sheep questions current nomenclature and provides evidence for domestication from two different subspecies. Proc. Biol. Sci., 269: 893-904.
CrossRefDirect Link - Tsunada, K., H. Chang , G.B. Chang ,W. Sun and T.S.H. Dorji et al., 2010. Phylogeny of local sheep breeds in East Asia, focusing on the bayanbulak sheep in China and the sipsu sheep in Bhutan. Biochem. Genet., 48: 1-12.
CrossRef - Librado, P. and J. Rozas, 2009. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics, 25: 1451-1452.
CrossRefPubMedDirect Link - Meadows, J.R.S., I. Cemal, O. Karaca, E. Gootwine and J.W. Kijas, 2007. Five ovine mitochondrial lineages identified from sheep breeds of the near East. Genetics, 175: 1371-1379.
CrossRef - Pedrosa, S., M. Uzun, J.J. Arranz, B. Gutierrezz-Gil, F.S. Primitivo and Y. Bayon, 2005. Evidence of three maternal lineages in near eastern sheep supporting multiple domestication events. Proc. R. Soc. B, 272: 2211-2217.
CrossRefDirect Link - Saitou, N. and M. Nei, 1987. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol., 4: 406-425.
CrossRefPubMedDirect Link - Tamura, K., J. Dudley, M. Nei and S. Kumar, 2007. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol., 24: 1596-1599.
CrossRefPubMedDirect Link - Tang, C.W., W.T. Xing, L. Shi, X. Yu and W.J. Liu et al., 2009. Microsatellite analysis of the genetic polymorphism in 13 sheep breeds and 1 hybrid group in Xinjiang. Chinese Vet. J., 36: 73-78.
Direct Link - Thompson, J.D., D.G. Higgins and T.J. Gibson, 1994. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res., 22: 4673-4680.
CrossRefPubMedDirect Link - Wang, X., Y.H. Ma, H. Chen and W.J. Guan, 2007. Genetic and phylogenetic studies of Chinese native sheep breeds (Ovis Aries) based on mtDNA D-loop sequences. Small Ruminant Res., 72: 232-236.
CrossRef - Yang, X.J. and Y.Zh. Zhao, 2002. Review on the genetics diversity of Tibetan sheep. J. Gansu Agric. Univ., 37: 395-399.
Direct Link