
G. Saki
Physiology Research Center, Ahvaz Joundishapour University of Medical Science, Ahvaz, Iran
M. Hashemitabar
Department of Anatomical Sciences, Faculty of Medicine, Ahvaz Joundishapour University of Medical Science, Ahvaz, Iran
M. Abdollahi
Physiology Research Center, Ahvaz Joundishapour University of Medical Science, Ahvaz, Iran
S.H. Razie
Physiology Research Center, Ahvaz Joundishapour University of Medical Science, Ahvaz, Iran
Asian Journal of Animal and Veterinary Advances
Year: 2011 | Volume: 6 | Issue: 7 | Page No.: 697-705







ABSTRACT
The aim of this study was to evaluate the effects of Vero cells on early embryonic cleavage rate and overcome cellblock of mice embryos in vitro. Female mice were super-ovulated by Intra-peritoneal injection of 5 IU Pregnant mare serum gonadotropine (PMSG) and 5 IU Human Chorionic Gonadotropine (HCG) 48 h later. The super-ovulated female NMRI mice placed individually with NMRI (Noda Medical Research Institute) males of proved fertility. The following morning, the females with positive vaginal plug were killed and cumulus-enclosed single cell embryos (2PN) were recovered. Two pronuclear (n=170) embryos were divided randomly into 2 groups: (1) co culture with vero cells (2) culture in simple culture medium. The rate of the development and the morphological appearance of mouse embryos in two groups were recorded daily for 120 h after retrieval in each system using inverted microscope. On day 5 of development the results showed that embryos cultured on vero cells had a significantly higher blastocyst and hatching formation rate than those in simple culture medium alone (p<0.05). It is concluded that Vero cells may improve mouse embryo development partly by increasing blastocyst formation, hatching blastocyst rate.
PDF Abstract XML References Citation
Received: December 17, 2010;
Accepted: February 10, 2011;
Published: June 15, 2011
How to cite this article
G. Saki, M. Hashemitabar, M. Abdollahi and S.H. Razie, 2011. Comparison of Mouse Embryo Cleavage Rate from One Cell Stage to Blastocyst Stage in Co-culture with Vero Cells and Without Co-culture. Asian Journal of Animal and Veterinary Advances, 6: 697-705.
DOI: 10.3923/ajava.2011.697.705
URL: https://scialert.net/abstract/?doi=ajava.2011.697.705
DOI: 10.3923/ajava.2011.697.705
URL: https://scialert.net/abstract/?doi=ajava.2011.697.705
INTRODUCTION
Despite the great advances that recently made in vitro fertilization procedures still, there is a high percentage of fetal loss before and other the transfer. Quality of the transferred embryos is the major factor that affects success of IVF (in vitro fertilization) (Edwards, 2007; Cetin et al., 2010). So it was suggested that to improve the quality, the embryos with high potential of growth have screened by longer period of culture. Thus embryos to be cultured in vitro still blastocyst stage and then transferred to the uterus (Lane and Gardner, 1997). Also studies show that the transfer time for human embryos should be in the same time with endometrial acceptance (Grewal et al., 2008; Mardon et al., 2007; Rebollar-Lazaro and Matson, 2010). Embryos co-culture till blastocyst stage and then transferring them provides this synchronization. Embryos in stages before implantation in most species of animals cannot develop to blastocyst in vitro situation and their growing will be stopped in a special stage. It used to be said that embryos growth stopping is because of the in vitro conditions and for any specie occurs at a special time. For example the growth of mouse and hamster s embryos in stage 2 cells of growth, cow in stage 4, pig in stage 4, cat, goat and sheep in stage 8 to 16 stops (Tesarik, 1989; Schramm et al., 2003; Banwell and Thompson, 2008). Many researches have been done to prevent fetal growth stopping in the libratory. It seems that this stopping is because of adverse (or suboptimal) conditions of culture and accurse in genomic activation stage or in embryos transition stage from tube to uterus (Joo et al., 2001). Co-cultural systems effectively prevent fetal growth block in vitro (Feng et al., 1994; Hajializadeh et al., 2008; Renroad, Nematollahi-Mahani et al., 2009). Researchers found that in human and different species of mammals, embryo toxic causes fetal block in different fetal growth stages in laboratory (Weichselbaum et al., 2002). Xie et al. (2005) found that blocking happens in genomic activation stage. Also, evidence showed that co-cultural system overcomes on the block. That this requires non-specific factors secreted from colleague cells such as glycoprotein's that make growth and embryo transition from block stage easy (Feng et al., 1996; Peura et al., 2010; Guerin and Nicollet, 1997). Bongso et al. (1993) described positive and negative aspects of co-cultural functions on embryos. One of negative aspects that referred to non-co-cultural situation and co-cultural can disappeared it, is presence of hypoxanthine. That is cause of block in mouse embryos. The main goal of creating co-cultural systems for human embryos is imitate paths that embryo experiences naturally in the uterus or fallopian tubes. Weichselbaum et al. (2002) reported that human embryos co-culture with epithelial cells of Fallopian tube is pula in vitro to protect cleavage deviations and human fetus growth causes decrease cleavage abnormalities, increase embryos qualities, production of same size blastomers and minimum of fragmentation in compared with normal. The coculture of human embryo with single longer of different cells has described. But obtain human samples is difficult and also use of human samples because of possible viral and bacterial infections is discussed. Animals have the same potential to infection but they provide a very accessible source of cells to co-culture. For the first time in 1990, Menezo used of immortal cell line with no genital origin call vero cells that their origin was epithelium of green African monkey's kidney (Menezo et al., 1990). Because kidney and genital organ have a same embryonic origin therefore vero cells and cells derived from genital organ have mesoderm origin. Vero cells are suitable kind of animal cells to use in coculturing with human embryos because due to their application in produce vaccines they will be well controlled against infections and viruses. Menezo et al. (1990) found that in coculture of human embryos with vero cells more than fifty percents of them reach to blastocyst stage and so lots of them pass from block stage. Coculture of vero cells with B2 environment system has better results according to amount of blastocyst formation. It seems that this system has a good growth potential (sixty percent in compare with three percent in control group). According to this observation, co-cultural system improves human embryos growth and prevents of their degeneration. Mouse and cow embryos were used to evaluate the effect of vero cells on embryos growth. Ouhibi et al. (1990) observed that vero cells overcome to cellular block of mouse embryo. To determine whether improvement of embryo growth is a result of deleting of waste materials or is because of secretion of amblyopic factors. Lai et al. (1992) have observed mouse embryos growth in co-cultural environment and non co-cultural environments. Results showed that vero cells cause mouse embryos s growth improvement in compare with the control group and this probably because of deleting the growth blocker materials. Myers et al. (1994) found that embryos growth in an environment full of vero cells has the same increase with environment with single layer of vero cells. Chen et al. (1994) also observed that vero cells have overcome to cellular block in mouse embryos in blastocyst formation duration and also cause improve of exiting of zona cells in blastocyst. Feng et al. (1996) observed the growth of embryo in different co-cultural systems including single layer of human granulosa, Bovine Oviduct Epithelial Cells (BOEC) and Bovine Uterine Epithelial Cell (BUEC). Rexroad and Powell (1988) observed that the Ham's F10 culture environment in spite of its beneficial effects on fetal growth causes death in somatic cleavage cells. Other researches showed that the DMEM medium is benefit for cells but it cannot support fetal growth well (Leppens and Sakkas, 1995). Huang et al. (2005) cultured mouse single cell embryos in different drops of KSOM medium (by adding glucose, amino acids, BSA) for five days. With attention to the previous studies in this field, it decided to make more researches and studies in this field and with use of different medium. In this study the KSOM medium was used. The advantage of KSOM culture environment in support of embryo s growth and pass of block cell stage is because of presence of low amounts of glucose in its compounds, so that embryos in this environment probably pass of block cell stage. The significant point is that glucose as energy substrate can not support development of mouse embryos till 4-8 cellular stage (Leese and Barton, 1984; Lane and Gardner, 2000). Presence of glucose in previous (early) stage of culture causes delay or even prevents of mammals embryos genesis in the culture environment (Lane and Gardner, 1997). In this study, the co- culture system with Vero cells was established to study its effect on early embryonic cleavage rate and overcome cell block of mice embryos in vitro.
MATERIALS AND METHODS
This experimental study was performed in physiology research center of Ahvaz Jondishapour University of Medical Sciences (AJUMS) from September 2009 to April 2010. Single layer of vero cells production: To produce single layer, after growth and proliferation of vero cells in flask, culture environment pilled on the surface of the cells and 5 cc of 5% trypsin solution in phosphate buffer in addition to EDTA 2 mg mL-1 were added to flask. Flask contains trypsin that shock continuously in 37°C temperature. After separating the cells from flask bottom and their floating, 10cc of DMEM + 10% FBS culture environment added to it till to prevent of more tripsin activity and damage cells. Solution contains vero cells transferred to test strill tubes and centrifuged with 400 g acceleration for five minutes. After pilling liquid on cells one of two culture environments added to it again and cells has been floated with the pipe. To count cells the neobar lam was used. Petri dish with 30 mm diameter was placed in it 100 μL drops of solution contains cell and a thin layer of paraffin oil covered the surface of it then Petri dish transferred to incubator under condition of 37°C and 5% CO2. Usually after two days, about 80% cover the floor of the drop.
Animals: This study was conducted from March 2009 to November 2009. A total 30 adult NMRI (Noda Medical Research Institute) female mice 8-10 weeks of age, weighing 55±6.6 g were purchased from Laboratory Animals Care and Breeding Center of Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran. Female mouse were super-ovulated by i.p. injection of 5 IU Pregnant mare serum gonadotropine (PMSG) and 5 IU human chorionic gonadotropine (HMG) 48 h later and then placed in male mouse's cage separately. Next day mouse with vaginal plug to obtain two pronucleous embryos were separated (Celis and Celis, 2002). Female mouse with positive vaginal plug was killed by cervical dislocation method and single cell embryos from the end of uterine tubes have flashed in M2 culture medium by using of 30G niddle. After removing the embryos from the uterine tubes, by use pipette, a few amount of hyaluronidase solution (1 mg mL-1) was added to medium in order to removed the cumulus cells around the zygotes and it repeated for many times. Then embryos transferred to M2 environment 50 μL drops that were under mineral oil. The final step is repeated several times to be ensure that medium be cleaned from the hyaluronidase and waste cells. After that time the two-pronuclear embryos cultured in 50 μL drops of KSOM medium that covered by mineral oil for 120 h. In that time the count of embryo who reached to two cells stage, four cells stage and pass of block cell stage were calculated.
Statistic methods: Cleavage division's speed and compare it with any group has been done with use of single side Fisher's ANOVA (Farouk and Vlad, 2008). Compare of number of the embryos who reach to other stages in each group has been done by non parametric Mann-Whitney test (Lai et al., 1992).
RESULTS
In this study, 30 NMRI female mice super-ovulated by i.p. injection of 5 IU PMSG and 5 IU HCG 48 h later, 170 single cell embryos (two pronuclear) obtained and randomly divided into two equal groups. 85 of embryos were cultured in simple medium and another 85 embryos cultured with vero cells. Cleavage division’s speed and reaching to two and four cells stage was evaluated by invert microscope at 24 and 48, 72, 96 and 120 h after the starting culture.
Table 1 shows that from total 85 embryos who cultured in simple medium until 24 h, 83 embryos (97.6%) reached to 2 cells stage but only 2(2.4%) remained in 1 cell stage and 82 of 85 embryos (96.5%) reached to 2 cells stage and 3 of 85 embryos (3.5%) remained in 1 cell stage when embryos cultured with vero cells for 24 h. Statistical analysis showed that the difference between two type of culture in cleavage division’s speed and reaching to two cell stage was not significant (p>0.05). In second days (48 h after cultured) in simple medium, 2 of 85 (2.4%) remained in same stage, 41 of 85 (48.2%) 42 of 85 embryos (49.4%) reached to 2 and 4 cells stage respectively but in co-cultured with vero cells group 1 (1.2%), 29 (34.1%) and 55 of 85 embryos (64.7%), remained in 1 cell stage and reached to 2 and 4 cells stage, respectively. Statistical analysis showed that disappearing of cellular block and reaching to 4 cell stage in embryos cultured with vero cells was increased significantly (p =0.04).
Data of Table 2 show that from total 85 embryos that cultured with Vero cell until 72 h, 5 embryos (5.88 %) reached to blastocyst stage but the blastocyst formation rate in another culture medium was zero. Statistical analysis showed that the difference between two type of culture in cleavage division’s speed and reaching to blastocyst stage was significant (p = 0.03).
| Table 1: | Cleavage division speed of NMRI mouse embryos cultured in simple and with vero cells until 48 h after culture starting |
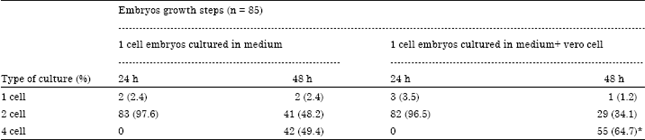 | |
| Values in parenthesis are percentages. Significant difference at p<0.05 | |
| Table 2: | Cleavage divisions speed of NMRI mouse embryos cultured in simple and with Vero cells until 96 h after culture starting |
 | |
| Values in parenthesis are percentage. *Significant difference at p<0.05 | |
| Table 3: | Cleavage divisions speed of NMRI mouse embryos cultured in simple and with Vero cells until 120 h after culture starting |
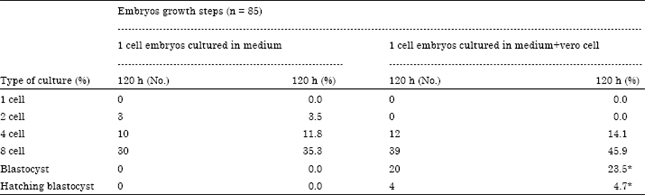 | |
| *Significant difference at p<0.05 | |
Present results in Table 3 showed that from total 85 embryos that cultured with Vero cell until 120 h, 20 embryos (23.5%) and 4 (4.7%) reached to blastocyst and blastocyst hatching stage respectively but all embryos who cultured in simple medium failed to reaching the blastocyst or hatching blastocyst stages. Statistical analysis showed that the difference between two type of culture in cleavage division’s speed and reaching to blastocyst and hatching blastocyst stage was significant (p = 0.001).
DISCUSSION
Embryos in stages before implantation in most species of mammals can not develop to blastocyst in vitro situation and their growing will be stopped in a special stage. It used to be said that embryos growth stopping is because of the in vitro conditions and for any specie accrues at a special time. For example the growth of mouse and hamsters embryos in stage 2 cells of growth, cow in stage 4, pig in stage 4, cat, goat and sheep in 8 to 16 stop (Tesarik, 1989; Banwell and Thompson 2008). Many researches have been done to prevent fetal growth stopping in the laboratory. It seems that this stopping is because of adverse conditions of culture and accurse in genomic activation stage or in embryo transition stage from pipe to uterus (Joo et al., 2001; Feng et al., 1994; Nematollahi-Mahani et al., 2009). Co-cultural systems effectively prevent fetal growth block in vitro (Feng et al., 1994; Nematollahi-Mahani et al., 2009). In the lots of co-cultural systems, the used basal medium is a simple ionic culture environment that is enough to embryos growth but it can be a danger for cleavage cells function. In other researches, the complex culture environment has been used that was designed for cleavage cells and was not enough to embryo s growth. The main problem is selecting the basal environment. Rexroad and Powell (1988) observed that the Ham's F10 culture environment is spite of its beneficial effects on fetal growth causes death in somatic colleague cells. Other research showed that the DMEM environment is benefit for cells. But it can't support fetal growth well (Leppens and Sakkas, 1995). In this research embryo in two different cultures, culture with vero cells and without them cultured to 4 cells stage (48 h after flashing) in KSOM culture environment. In co-cultural system with vero cells, 64.7% of embryos passed of block cell stage and in system without co-culture, 49.4% of embryos passed of block cell stage and reached to 4 cell steps. These data is reinforced the study of Weichselbaum et al. (2002). One of the most important of embryo needs before implantation is energy. Studies showed that private is necessary to prepare energy for zygote cell and its previous divisions (Brown and Whittingham, 1991). In the next stages presence of lactate is necessary and then glucose as an energy source is considered. Thus embryo till blastocyst stage doesn’t use of glycolyse to prepare energy and presence of glucose in the early steps of culturing causes delay in or even prevents of mammals embryos development in medium (Lane and Gardner, 1997; Brown and Whittingham, 1991; Suarez et al., 2005). Nutritional model of human embryos is very similar to the mouse embryos. Mouse oocyte and zygote consume a small amount of glucose than pyruvate but during the compression time glucose is taken more than pyruvate. So mouse ovule and zygote need pyruvate as the source of energy. Since two cells stage, lactate can be used at the same time with pyruvate. The significant point is that glucose as the pure energy substrate can not support the mouse embryo till 4 to 8 cells stage (Leese and Barton., 1984). Presence of glucose in the primary stages of culturing causes delay in development of mammals embryos in culture environment or even prevent of it (Gardner and Lane, 1997). So that advantage of KSOM environment in protection of fetal growth and it passing of cellular block stage is due to low amount of glucose in its compound, thus embryos in this environment pass of cellular block stage easily. Bongso et al. (1993, 1991) offered two possible mechanisms to co-cultural systems function: one, positive role that contains secretion of embryotrophic factors, substrates, growth factors and sytokinasis and another one, negative role that contains disposal of hazardous compounds, ammonium and free radicals use of activated metabolism of colleague cells. One of negative aspects that relives to the non co-cultural conditions and co-cultural conditions attribute them is presence of hypoxanthine that is known as the cause of block in mouse embryo. Embryo culturing of lots of pre implantation mammal’s embryos in a range of cultures including simple to improved cultures has been done. Honda et al. (1994) observed in mouse embryos coculturing with peritoneal macrophages that embryos passed of two cells block stage. Results showed that peritoneal macrophages have beneficial effect on the growth of mouse embryos in vitro. Menezo for the first time in 1990 used of immortal cell-line with non-genital origin called vero cells that their origin was the epithelium of green African monkey's kidney. Since kidney and genital organ have a same embryonic origin (mesoderm) there for vero cells derived from genital organ have mesoderm origin. Vero cells are suitable kind of animal cells to use in coculturing with human embryos because due to their application in produce vaccines, they will be well controlled against infections and viruses. Menezo et al. (1990) found that in human embryos coculturing with vero cells more than 50% of them reach to blastocyst stage that at least of blastocysts (13.25%) will reach to hatching stage. Our present showed that cultured of embryo with vero cell causes blastocyst formation and also hatching blastocyst significantly increases. This results in agreement with previous reports (Farouk and Vlad, 2008; Menezo et al., 1990). According to these observations co-cultural system, improves growth of human embryos and prevents of embryos degeneration. Also beneficial effects of coculturing are not just because of genital origin of cells but more because of their epithelial origin. Menezo et al. (1990) expressed that use of co-cultural systems and their success are depend on many factors that their most important are:
| • | Cultivated embryos specie |
| • | Type of co-culture environment as the basal environment |
| • | Type of cell lines that used as the colleague cells layer |
Mouse and cow embryos were used to evaluate vero cells effects on growth of embryos. Lai et al. (1992) showed that Vero cells cause improves growth of mouse embryo in compare with control group. Myers et al. (1994) found that growth of embryos in an environment full of vero cells have a same increasing in compare with use of single layer of vero cells. Chen et al. (1994) observed growth block of mouse embryos in HTF environment without protein. They observed that mouse embryos in HTF co-cultural environment with serum and obtained peptides of vero cells environment overcome the cellular block. They concluded that special factors secreted from vero cells are critical in mouse embryos passing of block stage. Ouhibi et al. (1990) has done the coculturing of mouse embryos on single layer of different cells including cow's oviduct cells, MDBK cells and vero cells. In co-culturing with vero cells single cell embryos did not pass cellular block stage. Initial primary study on metabolic changes caused by colleague cells showed that there were no relationship between decrease glucose rate and increase lactate and growth of embryos.
CONCLUSION
The results of present study show that co-culturing of the mouse embryos in KSOM basal environment and vero cells causes improvement in growth of mouse embryos by pass of cellular block stage and also the Vero cells may improved mouse embryo development partly by increasing blastocyst formation, hatching blastocyst rate.
ACKNOWLEDGMENT
This article is extracted from the M.Sc thesis for Abdollahi M. Special thanks to Ahvaz Joundishapour University of medical sciences for the financial support.
REFERENCES
- Banwell, K.M. and J.G. Thompson, 2008. In vitro maturation of mammalian oocytes: Outcomes and consequences. Semin. Reprod. Med., 26: 162-174.
CrossRefPubMedDirect Link - Bongso, A., C.Y. Fong, S.C. Ng and S. Ratnam, 1993. The search for improved in vitro systems should not be ignored: Embryo co-culture may be one of them. Hum. Reprod., 8: 1155-1160.
PubMed - Bongso, A., S.C. Ng, C.Y. Fong and S. Ratnam, 1991. Cocultures: A new lead in embryo quality improvement for assisted reproduction. Fertil. Steril., 56: 179-191.
PubMed - Brown, J.J. and D.G. Whittingham, 1991. The roles of pyruvate, lactate and glucose during preimplantation development of embryos from F1 hybrid mice in vitro. Development, 112: 99-105.
PubMed - Chen, H.F., H.N. Ho, S.C. Huang, T.Y. Lee and Y.S. Yang, 1994. Peptides extracted from Vero cell cultures overcome the blastocyst block of mouse embryos in a serum-free medium. J. Assist. Reprod. Genet., 11: 165-171.
PubMed - Edwards, R.G., 2007. Human implantation: The last barrier in assisted reproduction technologies. Reprod. Biomed., 1: 5-22.
PubMed - Farouk, F.N.O. and M. Vlad, 2008. In vitro development of mouse pronuclear embryos to blastocysts in sequential media with and without co-culture of autologous cumulus cells. J. Reprod. Dev., 54: 385-390.
PubMed - Feng, H.L., Q.Z. Yang, Q.Y. Sun, P.C. Qin and J.M. Liu, 1994. Development of early bovine embryos in different culture systems. Vet. Rec., 135: 304-306.
PubMed - Feng, H.L., X.H. Wen and S.C. Presser, 1996. Effect of different co-culture systems in early human embryo development. Hum. Reprod., 11: 1525-1528.
PubMed - Gardner, D.K. and M. Lane, 1997. Culture and selection of viable blastocysts: A feasible proposition for human IVF. Hum. Reprod., 3: 367-382.
PubMed - Grewal, S., J.G. Carver, A.J. Ridley and H.J. Mardon, 2008. Implantation of the human embryo requires Rac1-dependent endometrial stromal cell migration. Proc. Natl. Acad. Sci. USA., 105: 16189-16194.
PubMed - Guerin, J.F. and B. Nicollet, 1997. Interest of co-cultures for embryos obtained by in vitro fertilization: A French collaborative study. Hum. Reprod., 12: 1043-1046.
PubMed - Hajializadeh, N., H. Babaei, S. N. Nematollahi-Mahani and S. Azizollahi, 2008. The development of mouse early embryos in vitro in fibroblasts and cumulus cells co-cultures supplemented with retinoic acid. Iran. J. Vet. Res., 9: 1-8.
Direct Link - Honda, R., K. Matsuura, Y. Fukumatsu, T. Kawano and H. Okamura, 1994. In vitro enhancement ofmouse embryonic development by co-culture with peritoneal macrophages. Hum. Reprod., 9: 692-696.
PubMed - Huang, W.J., J. Yuan, X. Deng and J. An, 2005. Effects of modification of the nutritionalcomponents in potassium simplex optimized medium on preimplantation developmentand cleavage of mouse embryos in vitro. Di Yi Jun Yi Da Xue Xue Bao., 25: 256-261.
PubMed - Joo, B.S., M.K. Kim, Y.J. Na, H.S. Moon, K.S. Lee and H.D. Kim, 2001. The mechanism of action of coculture on embryo development in the mouse model: Direct embryo-to-cell contactand the removal of deleterious components. Fertil. Steril., 75: 193-199.
PubMed - Lai, Y.M., D.E. Stein, Y.K. Soong, Y.X. Tang and J. Grifo et al., 1992. Evaluation of Vero cell co-culture system for mouse embryos in various media. Hum Reprod., 7: 276-280.
PubMed - Leese, H.J. and A.M. Barton, 1984. Pyruvate and glucose uptake by mouse ova and pre-implantation embryos. J. Reprod. Fert., 72: 9-13.
PubMed - Lane, M. and D.K. Gardner, 2000. Lactate regulates pyruvate uptake and metabolism in the preimplantation mouse embryo. Biol. Reprod., 62: 16-22.
PubMed - Lane, M. and D.K. Gardner, 1997. Differential regulation of mouse embryo development and viability by amino acids. J. Reprod. Fertil., 109: 153-164.
PubMed - Leppens, G. and D. Sakkas, 1995. Differential effect of epithelial cell-conditioned medium fractions on preimplantation mouse embryo development. Hum. Reprod., 10: 1178-1183.
PubMed - Mardon, H., S. Grewal and K. Mills, 2007. Experimental models for investigating implantation of the human embryo. Semin. Reprod. Med., 25: 410-417.
PubMed - Menezo, Y.J.R., J.F. Guerin and J.C. Czyba, 1990. Improvement of human early embryo development in vitro by coculture on monolayers of Vero cells. Biol. Reprod., 42: 301-306.
Direct Link - Myers, M.W., J.R. Broussard, Y. Menezo, S.G. Prough, J. Blackwell, R.A. Godke and J.K. Thibodeaux, 1994. Established cell lines and their conditioned media support bovine embryo development during in vitro culture. Hum. Reprod., 9: 1927-1931.
PubMed - Nematollahi-Mahani, S.N., A. Nematollahi-Mahani, G. Moshkdanian, Z. Shahidzadehyazdi and F. Labibi, 2009. The role of co-culture systems on developmental competence of preimplantation mouse embryos against pH fluctuations. J. Assist. Reprod. Genet., 26: 597-604.
PubMed - Ouhibi, N., J. Hamidi, J. Guillaud and Y. Menezo, 1990. Co-culture of 1-cell mouse embryos on different cell supports. Hum. Reprod., 5: 737-743.
PubMed - Peura. T.T., J. Schaft and T. Stojanov, 2010. Derivation of human embryonic stem cell lines from vitrified human embryos. Methods Mol. Biol., 584: 21-54.
PubMed - Rexroad, C.E. and A.M. Jr, Powell, 1988. Co-culture of ovine eggs with oviductal cells and trophoblastic vesicles. Theriogenology, 29: 387-397.
PubMed - Rebollar-Lazaro, I. and P. Matson, 2010. The culture of human cleavage stage embryos alone or in groups: Effect upon blastocyst utilization rates and implantation. Reprod. Biol., 10: 227-234.
PubMed - Schramm, R.D., A.M. Paprocki and C.A. VandeVoort, 2003. Causes of developmental failure of in vitro matured rhesus monkey oocytes: Impairments in embryonic genome activation. Hum. Reprod., 18: 826-833.
PubMed - Suarez, S.S., D.F. Wolf and S. Meizl, 2005. Induction of the acrosome reaction in human spermatozoan by a fraction of human follicular fluid. Gamemrte. Res., 14: 107-111.
CrossRef - Tesarik, J., 1989. Involvement of oocyte-coded message in cell differentiation controlof early human embryos. Development, 105: 317-322.
PubMed - Weichselbaum, A., Y. Paltieli, R. Philosoph, B. Rivnay, R Coleman, M.M. Seibel and S. Bar-Ami, 2002. Improved development of very-poor-quality human preembryos by coculture with human fallopian ampullary cells. J. Assist. Reprod. Genet., 19: 7-13.
PubMed - Xie, Y., T. Sun, Q.T. Wang, Y. Wang, F. Wang, E. Puscheck and D.A. Rappolee, 2005. Acquisition of essential somatic cell cycle regulatory protein expression and implied activity occurs at the second to third cell division in mouse preimplantation embryos. Febs Lett., 579: 398-408.
PubMed