
Dan Du
College of Animal Science and Technology, China Agricultural University, Beijing 100193, China
Chunjiang Zhao
College of Animal Science and Technology, China Agricultural University, Beijing 100193, China
Hao Zhang
College of Animal Science and Technology, China Agricultural University, Beijing 100193, China
Guocai Han
College of Animal Science and Technology, China Agricultural University, Beijing 100193, China
Asian Journal of Animal and Veterinary Advances
Year: 2011 | Volume: 6 | Issue: 6 | Page No.: 564-571







ABSTRACT
Tibetan Horse lives in Qinghai-Tibet Plateau of South-West China, the controversy on whether Tibetan Horse should be classified into Southwest Horse Type or not has existed in China for a long time as there are four other type horses in the country. In the study, the genetic diversity and structure of Tibetan Horse were investigated using 14 microsatellite markers and 153 animals belonging to three Tibetan Horse subpopulations in Xigaze, Qamdo and Nagqu, respectively and Mongolian Horse and Ningqiang Pony which served as reference groups to Tibetan Horse. The results showed that the mean Number of Allele (MNA) across the 14 loci in Tibetan Horse was 13.1 and the average observed heterozygosity (0.710) and PIC (0.752) were slightly higher than those (0.692 and 0.773, respectively) of Mongolian Horse. Although, there was abundant genetic variation, genetic differentiation between horse populations was low, with only 2.1% of the total genetic variance among populations. Qamdo, Xigaze and Mongolian Horse had close relationship and Nagqu is distinctly separated from the other two Tibetan Horse subpopulations and referenced breeds in the clusters. The results indicated the genetic relationship of Tibetan Horse were closer to Mongolian Horse rather than to Ningqiang Pony, a typical breed in Southwest Horse Type, which suggests that Tibetan Horse was a unique horse breed which could not be classified into Southwest Horse Type.
PDF Abstract XML References Citation
Received: July 20, 2010;
Accepted: October 17, 2010;
Published: November 24, 2010
How to cite this article
Dan Du, Chunjiang Zhao, Hao Zhang and Guocai Han, 2011. Genetic Diversity of Tibetan Horse and its Relationships with Mongolian Horse and Ningqiang Pony Assessed by Microsatellite Polymorphism. Asian Journal of Animal and Veterinary Advances, 6: 564-571.
DOI: 10.3923/ajava.2011.564.571
URL: https://scialert.net/abstract/?doi=ajava.2011.564.571
DOI: 10.3923/ajava.2011.564.571
URL: https://scialert.net/abstract/?doi=ajava.2011.564.571
INTRODUCTION
There are five indigenous horse types in China, Tibetan Horse, Mongolian Horse, Southwest Horse, Hequ Horse and Kazakh Horse which were classified based on body conformation, distribution and history (Xie, 1986). Tibetan Horse, distributing around in four Southwest Provinces of China, Tibet, Qinghai, Sichuan and Yunnan, 60% of total live in Tibet. Tibetan Horse is classified into three different subpopulations as follows, Hill Type, accounting for 32% of total Tibetan Horse Population and in Qamdo prefecture; Altiplano Type, occupying 44% of the total and in Nagqu prefecture; Valley Type, making up 18.8% of the total and in Xigaze city (Xie, 1986).
Tibetan Horse is well adapted to high altitude in Qinghai-Tibet Plateau and extremely hardy and disease-resistant. Although, it lives in Southwest China, but some researchers classify it into a unique horse breed, while others believe it should be a breed in Southwest Horse Type together with other horse populations existing in Southwest region (Chang, 1995; Xie, 1986). Finally, the classification of Tibetan Horse remains controversial.
Mongolian Horse mainly distributes in Inner Mongolia region of Northern China, which had played an important role in world’s history. The great expansions of the Mongolian Empire under Attila (5th century) and Genghis Khan (13th century) resulted in the migrations of both its people and horses into its new territory, including Tibet (Sheng and Liu, 2006). Close relationship between the Mongolian native horse and Northern European horse breeds was observed by Bjornstad et al. (2002), thus there was a possible linkage between Mongolian Horse and Tibetan native horse.
The relationship among the three Chinese indigenous horse breeds, Tibetan, Mongolian and Southwest Horse, remains unclear up to now. In this study, Ningqiang Pony was referred to as Southwest Horse breeds. Though there are some reports on Mongolian Horse or Southwest Horse (Li et al., 2005; Sun, 2008), the genetic research on Tibetan Horse is absent. The only report on it was the research conducted for the zoological characteristics and polymorphism of blood proteins so far (Wang, 1996). The research of other livestock (chicken in Yu et al., 2006; sheep in Zhang et al., 2008b; goat in Mahmoudi, 2010) demonstrated that microsatellite is a useful tool for studying the genetic relationship among closely related breeds. In the study, we studied the genetic divergence of Tibetan Horse in Tibet using 14 microsatellite loci recommended by FAO and ISAG, which was the first time to study the three Tibetan horse subpopulations together as far as we know. It may reveal the genetic relationship among Tibetan Horse, North Horse and Southwest Horse. The information generated in this study may further be utilized for studying differentiation and relationship among different Chinese indigenous horse breeds.
MATERIALS AND METHODS
Sampling collection: This research project was fully sponsored by National Basic Research Program of China with grant number 2006CB102100. A total of 239 animals were sampled from Mongolian Horse in 2006, Ningqiang Pony in 2007 and the three Tibetan Horse populations in 2007. Blood or ear skin was randomly collected. The sampled populations and their geological information are shown in Table 1 and Fig. 1, respectively. Horse DNA was isolated from whole blood or ear tissues in Equine blood testing center of China Agricultural University (Beijing) according to a modified phenol-chloroform extraction method as Sambrook and Russell (2002) noted.
Microsatellite markers and genotyping: Fourteen microsatellite markers (AHT4, ASB17, ASB23, HTG7, HMS2, HTG6, HMS6, HTG10, LEX33, ASB2, VHL20, HMS3, HMS7, HTG4) dispersed over 10 different chromosomes were used in the present study. Each of the 14 microsatellites was amplified alone in independent PCR reaction.
| Table 1: | Sampling information of the studied horse breeds |
 | |
 | |
| Fig. 1: | Geographical location of the five horse breeds |
Forward primers were end-labelled with fluorescent dyes (6-FAM, TET or HEX). Genotypes for each marker were determined at State Key Laboratory of Agrobiotechnology (Beijing) using ABI 377 DNA Sequencer (Applied Biosystems) with the internal size standard GeneScanTM-TAMARA350 (Applied Biosystems).
Statistic analysis: Allele frequency, the number of alleles per locus, observed heterozygosity (Ho) and expected heterozygosity (He) for each breed, mean number of alleles (MNA) per locus and data format translation were calculated using Microsatellite Toolkit 3.1(http://animalgenomics.ucd.ie/sdepark/ms-toolkit).
Polymorphic Information Content (PIC) was calculated for each microsatellite locus (Botstein et al., 1980). The exact test for Hardy-Weinberg equilibrium across all loci and subpopulations were performed with GENEPOP (v. 1.2) with their standard deviations using Markov-Chain noted by Raymond and Rousset (1995).
Fixation coefficients (Fst, Fis and Fit) per pair of populations and their statistical significance were computed using the FSTAT program (Goudet, 2001). A hierarchical analysis was performed to examine the population differentiation using analysis of molecular variance (AMOVA) in the Arlequin (Schneider et al., 1997).
| Table 2: | Information on 14 microsatellite loci used in the study |
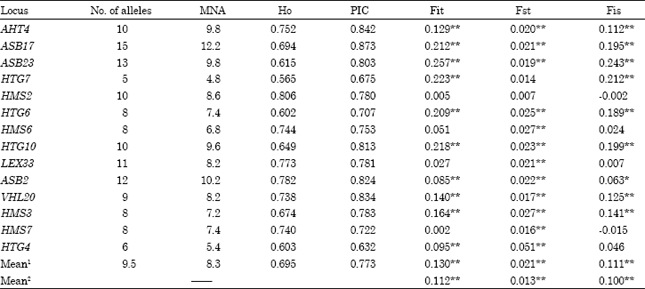 | |
| *p<0.05, **p<0.01. 1For all populations. 2For the 3 Tibetan Horse populations | |
| Table 3: | Diversity information on Tibetan horse populations, Mongolian horse and Ningqiang Pony |
 | |
| PA1: No. of private alleles | |
Principal Component Analysis (PCA) with gene frequency data was performed using the MVSP version 3.1 program (available at http://www.kovcomp.co.uk/mvsp/).
Nei’s genetic distance DA (Nei et al., 1983) and Nei’s standard genetic distance DS (Nei, 1972) between breeds were estimated and used to construct the Unweighted Pair Group Method with Arithmetic (UPGMA) mean dendrograms using DISPAN software (Ota, 1993).
RESULTS
Genetic variability: Totally, 133 alleles were detected across the 14 microsatellite loci (Table 2). Number of alleles per locus varies from 5 (HTG7) to 15 (ASB17). The MNA across 14 loci was 8.3 for the five horse populations. The observed heterozygosity across all loci in the total samples was 0.695 and ranged from 0.565 (HTG7) to 0.806 (HMS2). More information related to diversity across the microsatellite loci is given in Table 2.
The Mean Number of Alleles (MNA) in Tibetan Horse was 13.1, varied between 10.3 in Nagqu Horse and 16.1 in Xigaze Horse (Table 3), which might be explained by the variation in the sample sizes (The sample number for Nagqu Horse and Xigaze Horse were 39 and 61, respectively).
F-statistic: The Fst values for each locus were very close and were shown with Fit and Fis values in Table 2. Highly significant (p<0.01) genetic differentiation (Fst) was detected for all loci. On average, only 2.1% of the variation accounted for total population differentiation and breeds had a 11.1% (p<0.01) deficit of heterozygotes, whereas the total population had a 13% (p<0.01) deficit of heterozygotes.
| Table 4: | Pairwise population differentiation Fst estimates (below the diagonal) between horse breeds and Nei’s standard genetic distance (Ds, above the diagonal) among 5 Chinese local horse population |
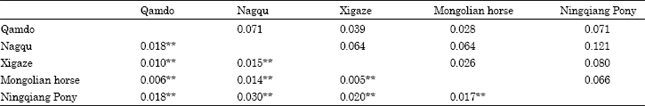 | |
| *p<0.05, **p<0.01 | |
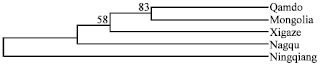 | |
| Fig. 2: | Dendrogram of relationships among 5 Chinese local horse populations using Ds genetic distance and the neighbor-joining method in Dispan programme |
Fst values were showed in Table 4 when breeds were considered in couples. The values of Fst between Tibetan Horse ranged from 1% for the Qamdo-Xigaze pair to 1.8% for the Nagqu-Qamdo pair. The highest Fst was detected between Nagqu and the Ningqiang Pony (3%).
Relationships among horse populations: Values above the diagonal in Table 4 represent the Ds genetic distance, using 14 microsatellite loci, ranged between 0.026 and 0.121 for the five breeds, the results of which were also very similar to ones of Fst. The low genetic distances among the five breeds indicated a close relationship among them.
DA and Ds distance matrices were used to build phylogenetic trees with UPGMA and NJ methods. The UPGMA dendrogram (Fig. 2) shows the relationship between horse populations and the reliability of the obtained tree was examined by 1000 bootstrap replicates. The most robust features of the topology were the cluster (83% support) formed by Qamdo and Mongolian Horse, then the cluster came together with Xigaze. The remaining breeds Ningqiang Pony and Nagqu had separate clusters.
DISCUSSION
All of the microsatellite loci genotyped were highly informative in Tibetan Horse, Mongolian Horse and Ningqiang Pony (PIC>0.6). The average observed heterozygosity in Tibetan Horse was 0.71, which was close with those (0.635-0.707) of Lipizzan horse (Achmann et al., 2004). The average PIC and expected heterozygosity for 14 loci in Mongolian Horse was 0.773 and 0.807, respectively, which was little different from the results (0.761 and 0.809) in Korea horse and Alien horse using the same microsatellite markers (Cho, 2006). Cho reported that Mongolian horse retained the largest amount of genetic variation in all the populations studied. Japanese reported Japanese native horse had lower diversity than Mongolian population and all of the native population were derived from Mongolian horse (Kakoi et al., 2007). Whereas, in the present study, the observed heterozygosity in Mongolian Horse (0.692) was lower than Cho’s report (0.833), but higher than Li’s (0.482). Tibetan Horse had higher genetic diversity than Mongolian Horse and Ningqiang Pony, in which Qamdo population was highest in observed heterozygosity (Ho = 0.762). It can be imagined that Mongolian native horse had decline in genetic diversity because of the policy in pasturing area caused the rapidly decline of Mongolian Horse, which restrains the populations of horse and domestic animals to protect the environment of grass land in Mongolia. The lowest genetic diversity of Ningqiang Pony was in agreement with its endangered status described (Ma et al.,2002; Zhang et al., 2008a; Sun, 2008).
Although, abundant genetic variation was detected, the population differentiation in the studied local horses was not significant. The value of 2.1% of the genetic variation was lower than those from other genetic diversity studies, e.g. 10% for Polish endangered Bilgoraj horses and two common horse breeds (Zabek et al., 2005) and 8% of the Spanish Celtic horse breeds (Canon et al., 2000), but was almost the same with the Fst(0.024) reported in Chinese (Ling et al., 2009). It suggested that our results were consistent with the researches on other Chinese indigenous horse breeds. However, the Fis(0.111) in the study was much higher than the Fis (0.021) in Ling’s. It suggested that the horse breeds in the study has inbreeding in an extent.
The corresponding UPGMA trees clustered the Mongolian native horse together with the Qamdo, the close association between Qamdo and Mongolian native horse support Northern influence on Tibetan Horse. The history records could be helpful to clarify that. Genetic Contribution of Mongolian Horse to Tibetan horse populations is in accordance with Mongolian army’s western aggressive immigration route and the direction of genetic contribution is compatible with human genetic studies (Chen et al., 2006). Chen reported though the contemporary Tibetans inhabit the Southwest China, Tibetans were genetically closer to Northern Han Chinese, Mongolian Chinese rather than to Southern Han Chinese.
Qamdo and Xigaze are both agricultural areas of the Tibetan Horse with comparatively lower altitude, round about 3500 m, where Tibetan Horse may be extensively influenced by Mongolian Horse. Therefore, their higher genetic relationship with Mongolian Horse is not surprising. As average altitude is about 4500 m in Nagqu prefecture, Nagqu Horse was hardly affected by any other horse populations such as Mongolian Horse, which could not adapt to the specific tough ecological condition. That may be an important reason why Nagqu horse is distinct separate with the other two Tibetan Horse populations and referenced breeds.
The results of the present study showed far genetic distance between Tibetan Horse and Ningqiang Pony which represented Southwest Type of the indigenous horses in China. The results are in accordance with previous description by Xie (1986) that Tibetan Horse is an ancient unique horse breed which could not be classified into Southwest Horse.
ACKNOWLEDGMENTS
We acknowledge the assistance of the following people with sample collection: Y. Chamba, H. Zhang, S. Lian, L. Deng. We are also grateful for the encouragement and valuable statistical suggestions provided by Y. Zhang and W..Liu
REFERENCES
- Achmann, R., I. Curik, P. Dovc, T. Kavar and I. Bodo et al., 2004. Microsatellite diversity, population subdivision and gene flow in the Lipizzan horse. Anim. Genet., 35: 285-292.
CrossRefPubMedDirect Link - Bjornstad, G., N.O. Nilsen and K.H. Roed, 2002. Genetic relationship between Mongolian and Norwegian horses. Anim. Genet., 34: 55-58.
PubMed - Botstein, D., R.L. White, M. Skolnick and R.W. Davis, 1980. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet., 32: 314-331.
PubMedDirect Link - Canon, J., M.L. Checa, C. Carleos, J.L. Vega-Pla, M. Vallejo and S. Dunner, 2000. The genetic structure of Spanish Celtic horse breeds inferred from microsatellite data. Anim. Genet., 31: 39-48.
PubMedDirect Link - Cho, G.J., 2006. Genetic relationship among the Korean native and alien horse estimated by microsatellite polymophism. Asian-Aust. J. Anim. Sci., 19: 784-788.
Direct Link - Kakoi, H., T. Tozaki and H. Gawahara, 2007. Molecular analysis using mitochondrial DNA and microsatellites to infer the formation process of Japanese native horse populations. Biochem. Genet., 45: 375-395.
PubMed - Ling, Y.H., Y.J. Cheng, Y.P. Wang, W.J. Guan and J.L. Han et al., 2009. Genetic diversity of 23 Chinese indigenous horse breeds revealed by microsatellite markers. Biodiversity Sci., 17: 240-247.
CrossRef - Nei, M., F. Tajima and Y. Tateno, 1983. Accuracy of estimated phylogenetic trees from molecular data: II. Gene frequency data. J. Mol. Evol., 19: 153-170.
CrossRefPubMedDirect Link - Raymond, M. and F. Rousset, 1995. GENEPOP (version 1.2). Population genetics software for exact tests and ecumenicism. J. Hered., 86: 248-249.
Direct Link - Ya-Bo, Y., W. Jin-Yu, D.M. Mekki, T. Qing-Ping and L. Hui-Fang et al., 2006. Evaluation of genetic diversity and genetic distance between twelve Chinese indigenous chicken breeds based on microsatellite markers. Int. J. Poult. Sci., 5: 550-556.
CrossRefDirect Link - Zabek, T., A. Nogaj, A. Radko, J. Nogaj and E. Stota, 2005. Genetic variation of Polish endangered Bilgoraj horses and two common horse breeds in microsatellite loci. J. Appl. Genet., 46: 299-305.
PubMedDirect Link - Zhang, X.Y., M.L. Zhou, X.H. Zhang and D.J. Wu, 2008. Study on population genetic structure of liangshan semi-wool sheep using microsatellite markers. Pak. J. Biol. Sci., 11: 2423-2427.
CrossRefPubMedDirect Link