
Jinming Huang
Center of Dairy Cattle Research, Shandong Academy of Agricultural Sciences, Jinan, 250131, People�s Republic of China
Li Liu
Center of Dairy Cattle Research, Shandong Academy of Agricultural Sciences, Jinan, 250131, People�s Republic of China
Hongmei Wang
Center of Dairy Cattle Research, Shandong Academy of Agricultural Sciences, Jinan, 250131, People�s Republic of China
Cuixia Zhang
Center of Dairy Cattle Research, Shandong Academy of Agricultural Sciences, Jinan, 250131, People�s Republic of China
Zhihua Ju
Center of Dairy Cattle Research, Shandong Academy of Agricultural Sciences, Jinan, 250131, People�s Republic of China
Changfa Wang
Center of Dairy Cattle Research, Shandong Academy of Agricultural Sciences, Jinan, 250131, People�s Republic of China
Jifeng Zhong
Center of Dairy Cattle Research, Shandong Academy of Agricultural Sciences, Jinan, 250131, People�s Republic of China
Asian Journal of Animal and Veterinary Advances
Year: 2011 | Volume: 6 | Issue: 1 | Page No.: 51-61







ABSTRACT
Toll-like receptor 2 (TLR2), a key component of the innate immune system, plays an important role in the initiation of the inflammatory response to foreign pathogens. This study was carried out to investigate the TLR2 gene expression difference in mammary gland tissues of the cows infected with mastitis and healthy cows and to search for variants of the bovine TLR2 gene for further clarifying the relationship between TLR2 gene and mastitis in cattle. The quantitative real-time polymerase chain reaction (qPCR) revealed that clinical mastitis caused by Staphylococcus aureus could significantly increased the LTR2 mRNA abundance (p<0.05). Seven novel single nucleotide polymorphisms (SNPs) in the 5’ upstream of the TLR2 gene in six cattle breeds (Hereford, Simmental, Limousin, Chinese indigenous Luxi Yellow cattle, Bohai Black cattle and Chinese Holstein cattle) were identified by the direct sequencing method. Four SNPs are located at the putative promoter region and their variants can alter the transcriptional factor binding sites. Genetic diversity showed that the SNP (4bp deletion) in the promoter was unique and two distinct haplotypes were found in Hereford cattle. Six SNPs in the TLR2 gene exon2 and 3’UTR were genotyped by PCR-RFLP and CRS-PCR methods in 398 Chinese Holstein cattle. Case-control study and associated analysis revealed that the cow with the genotype TT (3.15±0.52) in c.+189T>G has significantly lower (p<0.05) somatic cell score (SCS) than that with genotype TG (4.84±0.21) or GG (5.31±0.27) in Chinese Holstein cattle. In conclusion, haplotype analysis and linkage disequilibrium findings showed that the hapoltype TG of two loci (c.+189T>G and c.+631A>G) in the coding region may be used as a tolerance haplotype for the bovine mastitis.
PDF Abstract XML References Citation
Received: February 06, 2010;
Accepted: April 19, 2010;
Published: July 10, 2010
How to cite this article
Jinming Huang, Li Liu, Hongmei Wang, Cuixia Zhang, Zhihua Ju, Changfa Wang and Jifeng Zhong, 2011. Variants and Gene Expression of the TLR2 Gene and Susceptibility to Mastitis in Cattle. Asian Journal of Animal and Veterinary Advances, 6: 51-61.
DOI: 10.3923/ajava.2011.51.61
URL: https://scialert.net/abstract/?doi=ajava.2011.51.61
DOI: 10.3923/ajava.2011.51.61
URL: https://scialert.net/abstract/?doi=ajava.2011.51.61
INTRODUCTION
Mastitis is one of the most frequently occurring and complex diseases of the mammary gland in dairy cows and it causes a great economic loss. The prevalence of mastitis ranges from 10 to 50% (Neelesh, 2007). Epidemiologic analyses show the most prevalent agents associated with this mastitis infection are Staphylococcus aureus (S. aureus), Streptococcus strains and Escherichia coli (E. coli) pathogens (Chaneton et al., 2008).
Toll-like receptors (TLRs), key components of the innate immune system, are responsible for the initiation of the inflammatory response to foreign pathogens such as bacteria, fungi and viruses (Stevens et al., 2008). Toll-like receptor 2 (TLR2) is a member of TLRs and uniquely capable of recognizing the peptidoglycan and lipoteichoic acid from S. aureus and other gram-positive bacteria (Bannerman et al., 2004). Bovine TLR2 can properly transduce signals from S. aureus and E. coli. However, S. aureus fails to both activate NF-kappaB in mammary epithelial cells and to quickly induce the expressions of TNFalpha and interleukin-8 (CXCL8) in the udder (Yang et al., 2008). The TLR2 gene was strongly expressed during mastitis caused by S. aureus (Goldammer et al., 2004) and Strep. uberis (Swanson et al., 2009). The previous reports suggested that TLR2 may play a role in the host response to intramammary infections. Taken together, TLR2 appears to present an attractive candidate susceptibility gene for mastitis.
Bovine TLR2 gene, located to the proximal end of BTA 17, contains two exons and encodes 784 amino acids (White et al., 2003). Polymorphisms in the LTR2 gene encoding receptor associated with the innate immune system are likely to contribute to the overall variation in the resistance or susceptibility to mastitis in dairy cattle. Somatic cell score (SCS) represents a log score of the milk somatic cell count (SCC) and has been genetically correlated with clinical mastitis (Shook and Schutz, 1994). Several studies have identified some ploymorphic sites in the bovine TLR2 gene in the Bos indicus and Bos taurus cattle breeds (Jann et al., 2008; Opsal et al., 2008; Mariotti et al., 2009; Seabury et al., 2010) . Of which, three SNPs of TLR2 gene were localised at positions shaped by positive selection in the ruminant dataset (Leu227Phe, His305Pro, His326Gln) and in domains involved in the recognition of ligands, should be considered as candidate SNPs for immune related traits in cattle (Jann et al., 2008). However, no SNPs were found in Canadian Holstein bulls (Pant et al., 2008). Similarly, no significant association was found between the chromosomal regions surrounding TLR2 and mastitis in Norwegian Red cattle (Opsal et al., 2008). To data, there is no report on the polymorphisms of the bovine TLR2 gene promoter.
Therefore, the objectives of this study were: (1) to determine the TLR2 mRNA expressions in healthy and mastitis infected cow udders; (2) to clone and characterize the bovine TLR2 gene promoter region; (3) to investigate the genetic variations of the promoter of the bovine TLR2 gene in six cattle breeds (Hereford, Simmental, Limousin, Chinese indigenous Luxi Yellow cattle, Bohai Black cattle and Chinese Holstein cattle) and (4) to evaluate the associations between variations within the TLR2 gene with SCS in Chinese Holstein cattle.
MATERIALS AND METHODS
Animals and experiment designs
Experiment 1. Expressions of TLR2 mRNA of mammary tissues in healthy and clinical mastitis cows: Twelve udder tissue samples were collected aseptically from 6 slaughtered cows within 10 min after slaughtering in the slaughter house at Jinan, Shandong Province on February 2009. Base on the clinical symptoms and bacteriological test after culture with blood agar. The glands were categorized as infected by S. aureus and healthy groups. Two samples (infected and healthy) were collected from each cow. The udder tissues were collected from the deepest udder quarter and frozen in liquid nitrogen for RNA analysis. qPCR experiment was carried out from July 2009, to October, 2009, at the laboratory of the College of Animal Science and Technology, China Agricultural University.
Experiment 2. Polymorphisms of the promoter of the TLR2 gene among six cattle breeds: Six cattle breeds (26 Hereford, 30 Simmental, 156 Limousin, 46 Chinese indigenous Luxi Yellow cattle, 42 Bohai Black cattle and 225 Chinese Holstein cattle) were included in this experiment. Samples were obtained from July, 2007 to May, 2009 in Beijing and Shandong Province, China. The promoter region of the TLR2 was amplificated among six breeds. Then, their amplificated fragments were sequenced for the SNPs identification. The experiment was performed in 2009.
Experiment 3. Polymorphisms of the coding region and untranslated regions (UTR) of the TLR2 gene in Chinese Holstein cattle: A total of 398 cows, aged 3-6 years, were selected randomly from 8 dairy farms (Shandong, China) including in the Dairy Herd Improvement test scheme for the case-control study. Animals were milked three times daily by machine in farms. The animals with 5526±145.2 kg (mean±SE) 305-day milk yield per lactation. The DNA was extracted from blood samples following the standard phenol-chloroform method with minor modifications (Moller et al., 2004). Blood samples were collected from 2006-2008. Genotyping and data analysis were carried out in 2008-2009, at the laboratory of the Center of Dairy Cattle Research, Shandong Academy of Agricultural Sciences.
Milk samples were collected three times for one day and one time per month of lactation. SCC was measured by the Fossomatic cell counter (Foss Electric, FOSSMATIC 5000, Denmark). The control group was defined as the SCC below 1x105 cells mL-1 and without the record of clinical mastitis. The subclinical mastitis group (Case) was considered that SCC above 3x106 cells mL-1 in the milk of the cows.
Gene expression analysis: Total RNA was extracted from approximate 150 mg of mammary tissue using a TRNzol Kit (Tiangen, China) and further purified with the RNase-free DNase I (Promega, Madison, WI). RNA purity and concentration were measured with the ND-1000 Spectrophotometer (NanoDrop, Wilmington, USA). Next, the ImProm-II™ Reverse Transcription (Promega, USA) was used to convert approximately 1 μg of RNA from each sample to cDNA according to the modified protocol. A volume of 5 μL each reaction, containing 1 μg RNA, 4 μM Oligo(dT)15 (Tiangen, China) and nuclease-free water, was incubated at 70°C for 5 min, then chilled quickly at 4°C for 5 min and held on ice. Above reaction component was added to the 15 μL RTMix and annealed at 25°C for 5 min, extended the first strand for 1 h at 42°C, incubated for 15 min at 70°C. RTMix combined the 3.1 μL nuclease-free water, 4 μL ImProm-II™ 5xbuffer, 3 mM MgCl 2,0.5 mMdNTP (Tiangen, China), 20U Rnasin (Tiangen, China) and 1 μL ImProm-II™ Reverse Transcription (Promega, USA). qPCR was carried out using cDNA templates and the SYBRGreen PCR Master Mix (Tiangen, China) in ABI PRISM 7900 Sequence Detector System (Applied Biosystem, Foster City, CA) according to manufacture’s protocol. Each sample was run in triplicate. qPCR primers of TLR2 (F:5’-TGCGTTGGTTTGGATAGTGA-3’ and R: 5’-AGACCAGAGGGGATGGAGTT-3’; Size = 231 bp) and the internal control GAPDH (F:5’-GGGTCATCATCTCTGCACCT -3’ and R:5’- GGTCATAAGTCCCTCCACGA-3’; Size = 176 bp) were used for qPCR. The protocol was as follows: initial hold at 50°C for 2 min, then denaturation at 94°C for 3 min followed by 40 cycles of 94°C for 30 sec denaturation, 60.0°C for 30 sec annealing and 68°C for 20 sec extension. The last stage for the dissociation curve was as follows, 95°C for 15 sec, 60°C for 15 sec and 95°C for 15 sec. The relative quantification of TLR2 gene expression was calculated by the standard curve-based method for relative real time PCR (Larionov et al., 2005). The relative quantity of TLR2 mRNA was expressed by 100 fold of the mean.
Cloning and bioinformatics of the 5’ flanking region of the TLR2 gene: Primers TLR2-r-F and LTR2-r-R (Table 1) were designed to amplify the 925 bp fragment of the 5’ upstream of the TLR2 gene.
| Table 1: | The primers used in the amplification of TLR2 gene in cows and other PCR conditions |
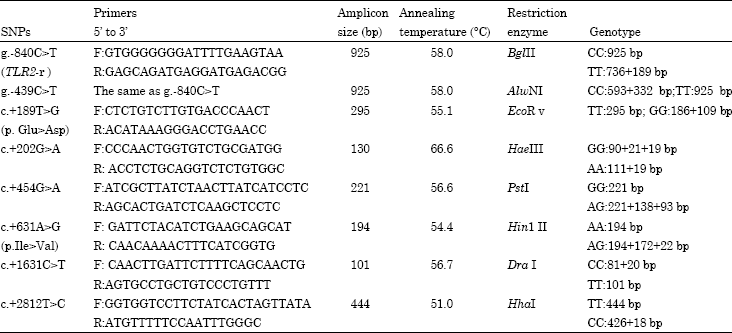 | |
The promoter and transcriptional factor binding sites were predicted with the MatInspector Professional (http://www.genomatix.de/).
Construction of DNA pools for SNP detection in the coding region and 3'-UTR: Ten cows with high SCS and 10 with low SCS were selected for creating DNA pools. Two DNA pools were constructed by aliquoting an equal amount of DNA from each selected cows in each group. These pools were used for the amplification of the promoter, exon2 and 3’-UTR of the TLR2 gene to detect SNPs. The PCR products were sequenced for SNPs searching using DNAMAN.
Genotyping: PCR-restriction fragment length polymorphism (PCR-RFLP) and Created Restriction Site PCR (CRS-PCR) methods (Huang et al., 2010) were used to genotype SNPs. Seven primer pairs (Table 1) were designed with primer 5.0 software based on reference sequence (GenBank No. NM.174197.2). PCR reaction was carried out in a total volume of 25 μL containing 50 ng DNA, 0.25 mM dNTPs, 2.0 mM MgCl2, 10 μM of each primer, 2 units Taq DNA polymerase (TaKaRa, China) and 10x Taq buffer. The PCR conditions were 95°C for 4 min, followed by 35 cycles of 95°C for 30 sec, at annealing temperature for 30 sec (Table 1), 72°C for 40 sec and a final elongation for 5 min at 72°C, with the PTC-200TM thermal cycler (MJ Research, Inc., Watertown, MA, USA). The PCR products were digested corresponding to the restriction enzyme (Table 1), following to the detection using 12% polyacryolamide gel electrophoresis and 3% agrose gel. The genotype was identified by the electrophoretic banding pattern.
Statistical analysis: The difference of the gene expressions between healthy and mastitis-infected groups was tested by paired T-test in SPSS version 11 (SPSS Inc, USA). The gene diversity, heterozygosity, polymorphism information content (PIC) and haplotype were analyzed by the powerMarker V3.25 software (Liu and Muse, 2005). Allele frequencies, haplotype and linkage disequilibrium (LD) analyses were determined by SNPAlyze version 3.2 software (http://www.dynacom.co.jp) in case-control study. Hardy-Weinberg equilibrium was assessed by Chi-Square analysis. The association analysis between the SNPs markers genotypes of the TLR2 gene and SCS were analyzed by the least squares method as applied in the GLM procedure of SAS(SAS Institute Inc,Cary, NC, USA). The applied linear model was as follows:
Yijklmn = μ + Fi+Gj +Sk+El + Hm + eijklmn |
where, Yijklmn was the observed value; μ was the overall mean; Fi was the fixed effect of farm; Gj was the fixed effect of genotype; Sk was the fixed effect of sire; El was the fixed effect of season; Hm was the fixed effect of parity and eijklmn was the random residual effect.
The p-value below 0.05 is regarded as significant.
RESULTS
Relative expression of TLR2 mRNA of the mammary glands in healthy and mastitis-infected cows: The relative expression of TLR2 mRNA in the infected mammary gland was significantly (p<0.05) higher than that in the healthy group (68.13±8.92 vs. 27.25±5.46).
Bioinformatics and polymorphism analysis of the promoter of TLR2 gene: The 925 bp (Simmental, Limousin, Luxi Yellow cattle, Bohai Black cattle, Holstein) and 921 bp (Hereford) fragments were obtained. The putative promoter of the bovine TLR2 gene locates at the position between -696 to -35 bp. Analysis of an approximately 661 sequence (-696 to -35) revealed 154 putative binding sites for various transcription factors (date not shown). No canonical TATA or CAAT boxes are found in the sequence analyzed. Seven novel SNPs were identified by the sequence multiple alignment. Four SNPs (g.-454 GAATdel, g.-439C>T, g.-221T>C and g.-211A>C) locates at the putative promoter region, whereas, another three SNPs (g.-840C>T, g.-760T>C, g.-758T>C) were not present in this region. In the 5’ regulatory region between the nucleotides -838 and -35, the putative SIXF [Sine oculis (SIX) homeodomain factors] binding site was found. Because of the mutation from C to T at the nucleotide -840, the SIXF was not found, whereas, GATA binding factors was appear. The 4bp deletion (GAAT) eliminated two SORY (SOX/SRY-sex/testis determining and related HMG box factors) and one HOXF (Paralog hox genes 1-8 from the four hox clusters A, B, C, D) transcriptional factor binding sites. However, in the locus g.-439C>T, the mutation results in an additional Zinc Finger Transcriptional Repressor (ZFTR) binding site.
Three SNPs in the 5’ flanking region of TLR2 gene were genotyped among six cattle breed populations. The genetic parameters of the three loci were as shown in Table 2 and 3. In the locus g.-840C>T, it was fixed in Limousin cattle population and was polymorphic among five breed populations (Table 3). In the locus g.-454 GAATdel, it was fixed among five cattle breeds and was polymorphic only in the Hereford cattle. All Hereford individuals are all heterozygote. The sequences alignment compared with the reference sequence (GenBank No. NM.174197.2) showed that 4 bp was deleted among five cattle breeds and the 4 bp (GAAT) of TLR2 is Hereford breed specific. In the locus g.-439C>T, it was polymorphic in five cattle breeds except for in the Simmental cattle. The number of haplotype of three SNPs ranged from 2 to 6 (Table 5). Haplotype analysis result revealed that there were two specific haplotypes (TAC and TAT) in Hereford cattle population.
| Table 2: | The genetic patterns of the 5’ flanking region of the bovine TLR2 gene among six cattle breeds* |
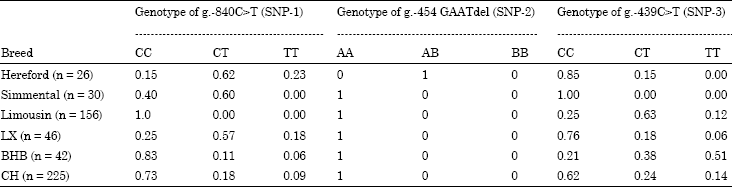 | |
| *LX: Luxi yellow cattle; BHB: Bohai black cattle; CH: Chinese holstein | |
| Table 3: | Diversity parameters of LTR2 gene among six cattle populations |
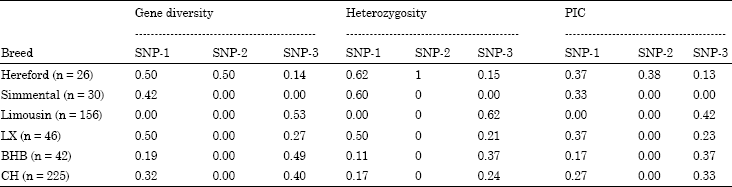 | |
| LX: Luxi yellow cattle; BHB: Bohai black cattle; CH: Chinese holstein | |
The genetic variants of the coding and 3’UTR regions of the bovine TLR2 gene in Case-control study: Table 4 shows the distribution of genotypic and allelic frequencies of the 6 SNPs in each group. Except for the c.+1631C>T (p = 0.001) in the case group, the observed and expected genotypic frequencies of the another five SNPs in the control and case groups were in good agreement with the predicted Hardy-Weinberg equilibrium values (data not shown). The genotypic and allelic frequency of c.+189T>G and the allelic frequency of c.+631A>G were significantly different between the control and case subjects (p<0.05). These data indicate that the distribution of c.+189T>G in cows can affect the overall analysis in the total subject group. Therefore, this could be used as a genetic marker of subclinical mastitis in cows, because the probability value for the allelic distribution of this SNP was significant.
In order to further assess the relationship of the two SNPs (c.+189T>G and c.+631A>G) and mastitis, the association analysis between two SNPs and SCS was performed. The results also revealed that genotype TT (3.15±0.52) in c.+189T>G has significant lower SCS (p<0.05) than that of genotypes TG (4.84±0.21) and GG (5.31±0.27). Because of the rarity of the T allele of the c.+189 locus in the present samples detected, a more large population is needed to confirm the result.
Possible Linkage Disequilibrium (LD) was evaluated by D’ and R2 values among six TLR2 gene polymorphisms (five in the exon2 and one in 3’-UTR) in subclinical and control cows (Table 6). LD analysis showed that c.+189T>G and c.+631A>G was not located in one haplotype block in the case and control groups. Therefore, the two SNPs were used to isolate the susceptibility haplotypes. Base on the results, the haplotype of the two SNPs were estimated and four haplotypes (GA, GG, TA, TG) were present. The distributions of 3 haplotypes (GA, GG and TA) showed no significant differences (p>0.05) in control and case groups.
| Table 4: | Genotype distribution in healthy and subclinical mastitis cows of screening analysis |
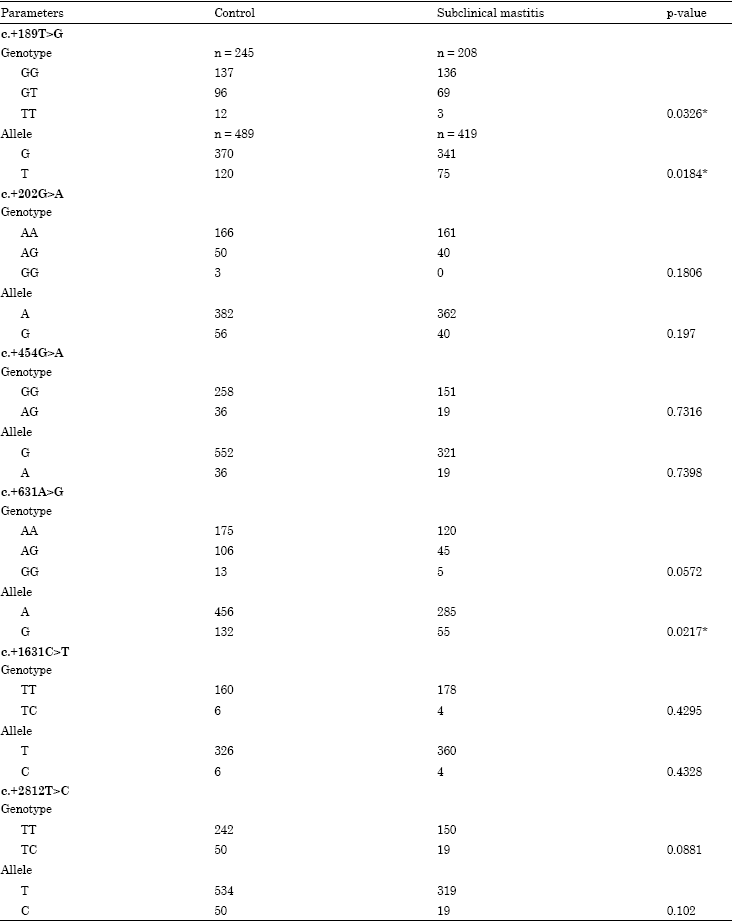 | |
| *Significant difference in distribution | |
| Table 5: | Haplotypical frequencies of three SNPs in the 5 flanking region of TLR2 gene among six breed populations |
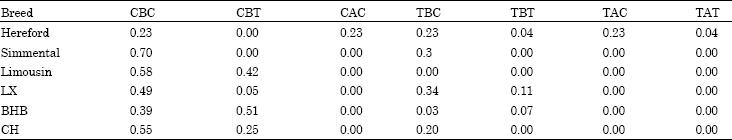 | |
| Table 6: | Linkage disequilibrium (LD) analyses of the control and case groups in Chinese Holstein cattle |
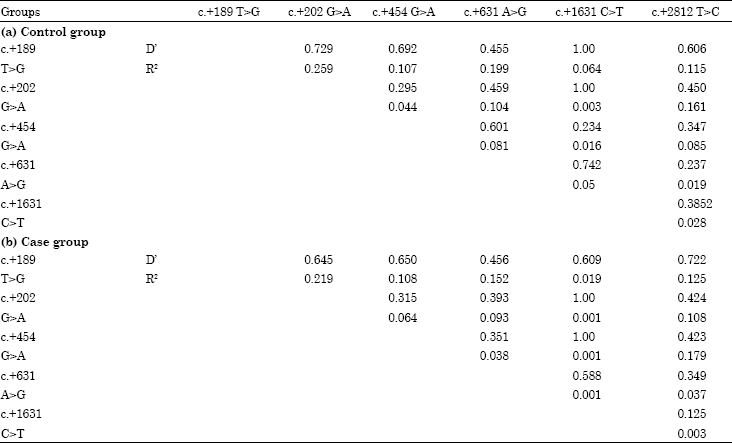 | |
However, the haplotype TG was significantly different (p<0.05). Therefore, the TG maybe used as a tolerance haplotype for the bovine mastitis.
DISCUSSION
Swanson et al. (2009) reported that TLR2 mRNA expressions were increased by the mastitis infected by the Strep. uberis, S. aureus and E. coil in udder tissues. However, no change was observed in the bovine mammary epithelial cells challenged by the same Strep.uberis. Our data is also consistent with the previous report by Goldammer et al. (2004). Heat-killed S. aureus and E. coli can activate equally well bovine TLR2 to induce NF-kappaB activation (Yang et al., 2008). The results suggest that mastitis increases the expression of TLR2 contributing to the innate immune defense system in the cow udder. It is suggested that the TLR2 gene may be considered as a candidate gene for mastitis resistance in Chinese Holstein cattle population.
The mutational SNPs in the 5’ regulatory region can alter the transcript factor binding sites, subsequently, resulting in the change of TLR2 gene expression. For example, SIX as a regulator of development and disease, is involved in the muscular development and can inactive the immune related gene Slc12a2 (Ando et al., 2005; Kumar, 2009). GATA binding factors was caused by the mutation from C to T at the nucleotide -840. It is interesting that GATA is a part of a multi-pathogen defense pathway that regulates innate immunity independently of the DAF-2/DAF-16 signaling pathway (Kerry et al., 2006). Furthermore, GATA is involved in the regulation of diverse processes including the development of the heart, haematopoietic system and sex gonads (He et al., 2007). HOXF can regulates numerous pathways during developmental and normal cellular processes (Svingen and Tonissen, 2006). Zinc Finger Transcriptional Repressor (ZFTR) binding site which promotes immortalization of human mammary epithelial cells (Nonet et al., 2001). These results can provide some basic information to better understanding the regulation of the TLR2 gene expression. To directly evaluate the functional relevance of the three SNPs polymorphisms, the transcriptional promoter activity using the luciferase reporter system are being conducted.
Haplotype analysis result showed that two haplotypes (TAC and TAT) were specific for Hereford cattle population. The difference may be caused by different selected purpose and their selected history of cattle.
TLR2 is critical in the immune response against gram positive bacteria. Three SNPs can alter the putative transcriptional factor binding sites and have different gene diversities and haplotypes among cattle breed populations, which suggests the polymorphisms in the promoter of TLR2 may affect the ability of the protein to recognise its related ligands. To our knowledge, this is the first report on the polymorphism of promoter of TLR2 gene in cattle.
Since, TLR2 as a major receptor can respond to a variety of peptides derived from gram-positive organisms, a mutation in TLR2 can affect the immune response to various bacterial stimuli (Brightbill et al., 1999). In human, the Arg753Gln mutation in TLR2 may predispose individuals to life-threatening staphylococcal infections. The mutation is located at the C terminus of human TLR2, it likely affects the signaling function of the molecule, rather than ligand binding (Lorenz et al., 2000). To date, several studies have identified some mutations in the coding and non-coding regions of TLR2 gene in Bos indicus and Bos taurus cattle breeds (Seabury and Womack, 2008; Mariotti et al., 2009; Zhang et al., 2009; Seabury et al., 2010). Furthermore, they demonstrated that they can enhance the ability of resistance for infection disease. Whereas, no significant association between the chromosomal regions surrounding TLR2 and mastitis in Norwegian Red cattle was found by a combined linkage and linkage disequilibrium methods (Opsal et al., 2008).
TLR2 have a common domain architecture: an extra-cellular domain containing 20 Leucine Rich Repeats ( AA54 -AA584), a transmembrane domain (AA585-AA607) and an intracellular TIR domain at AA633-AA783 (Jann et al., 2008). In the present study, the SNP (c.+189T>G) is located at the extra-cellular domain region, can alter amino acid (Glu to Asp), which further possibly affect the function of the TLR2 gene in part.
CONCLUSION
In conclusion, the present study reconfirmed the finding that the clinical mastitis caused by the S. aureus could significantly enhance TLR2 mRNA in mammary gland in Chinese Holstein cattle. The bovine TLR2 promoter was firstly cloned and characterized. Seven novel SNPs were revealed and two distinct hapolotypes (TAC and TAT)) of the g.-840C>T, g.-454 GAATdel and g.-439C>T loci in the 5’ regulatory region were found in Hereford cattle. The haplotype TG of two SNPs (c.+189T>G and c.+631A>G) in the coding region maybe used as a tolerance haplotype for the bovine mastitis.
ACKNOWLEDGMENTS
This research was supported by 863 Project of the Ministry of Science and Technology, Peoples Republic of China (No. 2006AA10Z1D9), Project in the National Science and Technology Pillar Program during the 11th Five-Year Plan of China (2008BADB2B09), Youth Science Foundation from Shandong Academy of Agriculture Science (No. 2006YQN030), Program of National Cow Industrial Technology System (No. nycytx-0107) and Projects of the Department of Science and Technology of Shandong Province (2007LZ10-04, Y2007D72).
REFERENCES
- Ando, Z., S. Sato, K. Ikeda and K. Kawakami, 2005. Slc12a2 is a direct target of two closely related homeobox proteins, Six1 and Six4. FEBS J., 272: 3026-3041.
PubMed - Bannerman, D.B., M.J. Paape, J.W. Lee, X. Zhao, J.C. Hope and P. Rainard, 2004. Escherichia coli and Staphylococcus aureus elicit different innate immune response following intra-mammary infection. Clin. Diag. Lab. Immun., 11: 463-472.
CrossRefDirect Link - Brightbill, H.D., D.H. Libraty, S.R. Krutzik, R.B. Yang and J.T. Belisle et al., 1999. Host defense mechanisms triggered by microbial lipoproteins through toll-like receptors. Science, 285: 732-736.
PubMed - Chaneton, L., L. Tirante, J. Maito, J. Chaves and L.E. Bussmann, 2008. Relationship between milk lactoferrin and etiological agent in the mastitic bovine mammary gland. J. Dairy Sci., 91: 1865-1873.
CrossRef - Goldammer, T., H. Zerbe, A. Molenaar, H.J. Schuberth, R.M. Brunner, S.R. Kata and H.M. Seyfert, 2004. Mastitis increases mammary mRNA abundance of β-defensin 5, Toll-Like-Receptor 2 (TLR2) and TLR4 but not TLR9 in cattle. Clin. Diagn. Lab. Immunol., 11: 174-185.
CrossRefDirect Link - He, C.J., H. Cheng and R. Zhou, 2007. GATA family of transcription factors of vertebrates: Phylogenetics and chromosomal synteny. J. Biosci., 32: 1273-1280.
Direct Link - Huang, J., H. Wang, C. Wang, J. Li, Q. Li, M. Hou and J. Zhong, 2010. Single nucleotide polymorphisms, haplotypes and combined genotypes of lactoferrin gene and their associations with mastitis in Chinese Holstein cattle. Mol. Biol. Rep., 37: 477-483.
CrossRefDirect Link - Jann, O.C., D. Werling, J.S. Chang, D. Haig and E.J. Glass, 2008. Molecular evolution of bovine Toll-like receptor 2 suggests substitutions of functional relevance. BMC Evol. Biol., 8: 288-288.
PubMed - Kerry, S., M. TeKippe, N.C. Gaddis and A. Aballay, 2006. GATA transcription factor required for immunity to bacterial and fungal pathogens. PLoS ONE, 1: e77-e77.
CrossRef - Kumar, J.P., 2009. The sine oculis homeobox (SIX) family of transcription factors as regulators of development and disease. Cell Mol. Life Sci., 66: 565-583.
PubMed - Larionov, A., A. Krauseand and W. Miller, 2005. A standard curve based method for relative real time PCR data processing. BMC Bioinform., 6: 62-78.
PubMed - Liu, K. and S.V. Muse, 2005. PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics, 21: 2128-2129.
CrossRefDirect Link - Lorenz, E., J.P. Mira, K.L. Cornish, N.C. Arbour and D.A. Schwartz, 2000. A novel polymorphism in the toll-like receptor 2 gene and its potential association with Staphylococcal infection. Infect. Immun., 68: 6398-6401.
Direct Link - Mariotti, M., J.L. Williams, S. Dunner, A Valentini and L. Pariset, 2009. Polymorphisms within the Toll-Like Receptor (TLR)-2, -4 and -6 genes in cattle. Diversity, 1: 7-18.
Direct Link - Moller, M., F. Berg, J. Riquet, D. Pomp and A. Archibald et al., 2004. High-resolution comparative mapping of pig Chromosome 4, emphasizing the FAT1 region. Mamm. Genome., 15: 717-731.
PubMed - Sharma, N., 2007. Alternative approach to control intramammary infection in dairy cows: A review. Asian J. Anim. Vet. Adv., 2: 50-62.
CrossRefDirect Link - Nonet, G.H., M.R. Stampfer, K. Chin, J.W. Gray, C.C. Collins and P. Yaswen, 2001. The ZNF217 gene amplified in breast cancers promotes immortalization of human mammary epithelial cells. Cancer Res., 61: 1250-1254.
Direct Link - Opsal, M.A., S. Lien, S. Brenna-Hansen, H.G. Olsen and D.I. Vage, 2008. Association analysis of the constructed linkage maps covering TLR2 and TLR4 with clinical mastitis in Norwegian Red cattle. J. Anim. Breed. Genet., 125: 110-118.
PubMed - Pant, S.D., F.S. Schenkel, I. Leyva-Baca, B.S. Sharma and N.A. Karrow, 2008. Identification of polymorphisms in bovine TLR2 and CARD15, associations between CARD15 polymorphisms and milk somatic cell score in Canadian Holsteins and functional relevance of SNP c.3020A>T. Dev. Biol., 132: 247-253.
PubMed - Seabury, C.M. and J.E. Womack, 2008. Analysis of sequence variability and protein domain architectures for bovine peptidoglycan recognition protein 1 and Toll-like receptors 2 and 6. Genomics, 92: 235-245.
PubMed - Seabury, C.M., P.M. Seabury, J.E. Decker, D.R. Schnabel, J.F. Taylor and J.E. Womack, 2010. Diversity and evolution of 11 innate immune genes in Bos taurus taurus and Bos taurus indicus cattle. Proc. Natl. Acad. Sci., 107: 151-156.
Direct Link - Shook, G.E. and M.M. Schutz, 1994. Selection on somatic cell score to improve resistance to mastitis in the United States. J. Dairy Sci., 77: 648-658.
CrossRef - Stevens, V.L., A.W. Hsing, J.T. Talbot, S.L. Zheng and J. Sun et al., 2008. Genetic variation in the toll-like receptor gene cluster (TLR10-TLR1-TLR6) and prostate cancer risk. Int. J. Cancer, 123: 2644-2650.
PubMed - Svingen, T. and K.F. Tonissen, 2006. Hox transcription factors and their elusive mammalian gene targets. Heredity, 97: 88-96.
Direct Link - Swanson, K.M., K. Stelwagen, J. Dobson, H.V. Henderson, S.R. Davis, V.C. Farr and K. Singh, 2009. Transcriptome profiling of Streptococcus uberis-induced mastitis reveals fundamental differences between immune gene expression in the mammary gland and in a primary cell culture model. J. Dairy Sci., 92: 117-129.
Direct Link - White, S.N., S.R. Kata and J.E. Womack, 2003. Comparative fine maps of bovine toll-like receptor 4 and toll-like receptor 2 regions. Mamm. Genome, 14: 149-155.
Direct Link - Yang, W., H. Zerbe, W. Petzl, R.M. Brunner and J. Gunther et al., 2008. Bovine TLR2 and TLR4 properly transduce signals from Staphylococcus aureus and E. coli, but S. aureus fails to both activate NF-κB in mammary epithelial cells and to quickly induce TNFα and interleukin-8 (CXCL8) expression in the udder. Mol. Immunol., 45: 1385-1397.
PubMed - Zhang, L.P., Q.F. Gan, T.H. Ma, H.D. Li and X.P. Wang et al., 2009. Toll-like receptor 2 gene polymorphism and its relationship with SCS in dairy cattle. Anim. Biotechnol., 20: 87-95.
CrossRefPubMedDirect Link