
Anru Wang
College of Veterinary Medicine, China Agricultural University, Beijing, People�s Republic of China
Fenghua Liu
Department of Animal Science and Technology, Beijing University of Agriculture, Beijing, People�s Republic of China
Zhepeng Wang
College of Animal Science and Technology, China Agricultural University, Beijing, People�s Republic of China
Xue Jiang
Beijing Da Bei Nong Technology Group Co. Ltd. (DBN), State Key Laboratory of Biological Feed Engineering, Beijing, People�s Republic of China
Wei Wang
Department of Animal Science and Technology, Beijing University of Agriculture, Beijing, People�s Republic of China
Kedao Teng
College of Veterinary Medicine, China Agricultural University, Beijing, People�s Republic of China
Jianqin Xu
College of Veterinary Medicine, China Agricultural University, Beijing, People�s Republic of China Key Laboratory of Development and Evaluation of the Chemical and Herbal Drugs for Animal Use, Ministry of Agriculture, Beijing, People�s Republic of China
Asian Journal of Animal and Veterinary Advances
Year: 2011 | Volume: 6 | Issue: 1 | Page No.: 36-50







ABSTRACT
The purpose of this study was to establish a model of avian viral disease for antiviral drug discovery and development. A Chinese IBDV strain BC6/85 was inoculated into 5-week-old specific-pathogen-free (SPF) male White Leghorn chicken and the clinicopathologic features of disease were compared with those reported before. Birds were monitored clinically and euthanized sequentially with collection of tissues. Tissues were examined by histopathology to observe microscopic lesions, by immunohistochemistry to detect viral protein, by electron microscope to observe ultrastructural changes of the plical epithelium of the bursa of Fabricius and by flow cytometry to detect apoptosis in bursal lymphocytes. Comparing of mean body weight gains and bursal weight indices between control group and virus inoculated groups, significant decreases are displayed. It was observed that the young chickens infected developed the typical histopathological changes of disease. The bursa of Fabricius revealed lesions characteristic of infectious bursal disease and virus particles identical to IBDV were found in the necrotic cells of the follicles. The mucosal surface of the bursal plica was erosion, ulceration and became smooth in the infected chicken groups. And the apoptotic cells were increased after 2 days of the virus inoculation. Morphologically, we clearly demonstrated that replication of IBDV in the bursa during the acute phase of infection and provided new information on the pathogenesis of IBDV.
PDF Abstract XML References Citation
Received: April 27, 2010;
Accepted: June 12, 2010;
Published: July 10, 2010
How to cite this article
Anru Wang, Fenghua Liu, Zhepeng Wang, Xue Jiang, Wei Wang, Kedao Teng and Jianqin Xu, 2011. Pathological Study of SPF Chickens Experimentally Infected with a Chinese IBDV Strain BC6/85. Asian Journal of Animal and Veterinary Advances, 6: 36-50.
DOI: 10.3923/ajava.2011.36.50
URL: https://scialert.net/abstract/?doi=ajava.2011.36.50
DOI: 10.3923/ajava.2011.36.50
URL: https://scialert.net/abstract/?doi=ajava.2011.36.50
INTRODUCTION
Infectious Bursal Disease (IBD) is an acute, highly contagious viral disease of young chickens often known as Gumboro disease (Lasher and Shane, 1994; Lukert and Saif, 2003). Infectious Bursal Disease Virus (IBDV) produces two distinctly different disease symptoms namely the clinical form and the subclinical form in susceptible chickens depending on age at infection. Clinical and subclinical infection with IBDV may cause immunosuppression (Sharma et al., 2000; Uddin and Hossain, 2006; Ahmed and Akhter, 2003). The clinical form is mostly observed in chickens of 5-8 weeks of age with typical lesions of IBD. The infectious disease causes considerable economic losses in the poultry industry worldwide. IBDV has a tropism to actively dividing pre-B lymphocytes, primarily in the bursa of Fabricius, but also in other organs of the immune system. The etiological agent is a member of the genus Avibirnavirus in the family Birnaviridae (Kibenge et al., 1988; Muller et al., 2003), whose genome consists of two double-stranded RNA segments designated A and B, which are enclosed within a nonenveloped icosahedral capsid. The genome segment B (2.9 kb) encodes VP1, the putative viral RNA polymerase. The larger segment A (3.2 kb) that contains two partially overlapping open reading frames (ORFs) encodes viral proteins VP2 (outer capsid), VP4 (protease), VP3 (inner capsid) and VP5 (non-structural protein). Among them, VP2 protein contains important neutralizing antigenic sites and elicits protective immune response and most of the Amino Acid (AA) changes between antigenically different IBDVs are clustered in the hypervariable region of VP2. Thus, this hypervariable region of VP2 is the obvious target for the molecular techniques applied for IBDV detection and strain variation studies. After infection IBDV multiplies rapidly in the B lymphocytes of the bursa of Fabricius, leading to immunosuppression, increased susceptibility to other diseases and reduced growth rate of surviving animals (Kibenge et al., 1988; Becht and Muller, 1991). The bursa is the principal reservoir of virus replication and peak virus titers in the bursa can be detected between 3 to 5 days after IBDV infection. In the bursae of chickens infected with IBDV, productive viral replication is often associated with necrosis, apoptosis of lymphoid cells, inflammatory change, atrophy and hemorrhages (Van den Berg, 2000; Kim et al., 2000; Taylor et al., 2008).
Results of previous studies indicated a correlation between IBDV replication and the proportion of apoptosis in bursal cells (Nieper et al., 1999; Jungmann et al., 2001). Remarkably, apoptotic cells were frequently located in the vicinity of antigen-expressing cells (Tanimura and Sharma, 1998; Jungmann et al., 2001; Taylor et al., 2008). This suggests the involvement of apoptotic processes in the pathogenesis of the disease. It is caspase dependent and activates caspases 3 and 9. Nuclear factor kappa B (NF-κB) is also activated and is required for IBDV-induced apoptosis during IBDV infection. The small, non-structural protein VP5 performs the function of inhibiting apoptosis initiated by viral replication and prevents infected cells from undergoing cell death before the virus finishes its life cycle (Liu and Vakharia, 2006). In order to understand the role of apoptosis in IBD and to elucidate the underlying molecular mechanisms, it is of importance to reliably detect and quantitate apoptotic cells (Schutte et al., 1998). The kinetics of Infectious Bursal Disease Virus (IBDV) replication and induction of apoptosis were investigated in vitro and in vivo. These data suggested that B cells may not be the sole target for the virus; macrophages and possibly other cells may serve as host for IBDV (Khatri et al., 2005). Extensive viral replication was accompanied by an infiltration of T cells in the bursa after virus infection. The results suggest that intrabursal T cells and T-cell-mediated responses may be important in viral clearance and promoting recovery from infection (Kim et al., 2000).
Although garlic was exalted throughout history, the quest to understand its action began only relatively recently (Harris et al., 2001). The main biological effects of garlic (Allium sativum) are attributed to its characteristic organosulfur compounds (Miron et al., 2000). The antimicrobial effects of aqueous garlic extracts are well established but those of garlic oil are little known (Ross et al., 2001). The purpose of the present study was to establish a model for infectious viral diseases with focus on IBD, so as to well understand the antiviral molecular mechanisms of garlic oil, providing a scientific evidence for modern drug development of a traditional medicine.
MATERIALS AND METHODS
Animals and viruses: 3-week-old specific-pathogen-free male White Leghorn chickens were used from Merial Vital Laboratory Animal Technology CO., Ltd. (Beijing, China) (2008) and reared in CC•JH-1-type positive pressure isolation units for the duration of the study. Birds for different experimental groups were housed in separate isolation units. Feed and water were provided ad libitum. A virulent strain of serotype 1 IBDV was purchased from China Institute of Veterinary Drug Control (Beijing, China). BC6/85 (CVCC AV7) strain is a conventional virulent type, which was shown to cause immunosuppression but little or no mortality. At 35-day-old, chickens were inoculated in each experiment with 100 bursal infective doses (BIDs/bird) of IBDV or phosphate-buffered saline (PBS) by eye drop.
Experimental design: In a first series of experiments one group of animals (15 chickens) were inoculated and euthanatized at 2, 4, 8 days postinoculation (days p.i.); control birds (totally 9 birds, 3 birds once) received the vehicle alone and were also euthanatized at the same day. The bursa of Fabricius, thymus, spleen, Harderian gland, caecal tonsils and liver were examined at 2, 4, 8 days p.i. for histopathological lesions. Tissue sections stained with haematoxylin and eosin were subjected to image analysis to quantify cellular depletion in the follicles of the BF.
In a second series of experiments, a total of 130 3-week-old SPF chickens were allotted randomly to three groups: Control group (40 chickens), Group I (45 chickens) and Group II (45 chickens). Group II chickens were intraorally administered with garlic oil (10 mg per kg BW) prior to IBDV inoculation, once per day, five consecutive days. Six hours after administration of garlic oil, each chicken from Group I and Group II was inoculated intraocularly with 0.1 mL of inoculum containing 100 bursal infective doses of BC6/85, respectively and reared separately. Control group was left as the mock-inoculation group, only given sterilized physiological saline by eye drop. At 2, 3, 5, 6, 9, 10, 15 and 18 days p.i., 3 chickens from each group were randomly selected and euthanatized for sample collection. Each bird was weighed and blood was drawn by heart or wing vein puncture at the times indicated. By the end of experiments, the bursa of Fabricius and spleen from killed birds (n = 19) were also weighed; and bursal and spleen weight indices were determined as follows:
All animal research was approved by China Agricultural University Animal Research Committee.
Histopathological examination: After establishing the IBD model of chicken, the animals were euthanatized for welfare reasons. For the detection of histopathological lesions, three chickens per group were examined at 2, 4 and 8 days postinoculation. Tissue samples were immediately collected, fixed in 10% phosphate-buffered formalin, dehydrated, embedded in paraplast embedding media X-tra (P3808, Sigma-Aldrich, St. Louis, MO, USA). Paraffin sections (5 μm) were de-waxed, routinely stained with hematoxylin and eosin (H and E, Sigma-Aldrich, USA), mounted with neutral balsam (Sinopharm Chemical Reagent Co., Ltd, Shanghai, China). Lesions were observed and photographed using an Olympus BX51 bright-field microscope (Olympus, Tokyo, Japan). Bursa lesion scores were determined and compared between groups.
Immunohistochemical detection of IBDV antigen in bursal sections: For immunolabelling of viral antigen, three chickens per group were also examined at 3, 6, 9, 15, 18 days postinoculation. A paraffin-embedded tissue slice of each bursa was also used for immunohistochemistry staining as previously described (Tanimura et al., 1995). Briefly, the sections were deparaffinized and hydrated. The endogenous peroxidase in sections was eliminated by 3% H2O2. Nonspecific reactions were blocked by normal rabbit serum. Mouse anti-IBDV mAb was used as primary antibody. Samples were then incubated overnight in a humidified chamber at 4°C with the primary antibody against IBDV diluted 1:1000. Mouse anti-VP2 monoclonal antibody was kindly provided by Dr Honglei Gao from Harbin Veterinary Research Institute, the Chinese Academy of Agricultural Sciences, Harbin, China. Whereas a 1:100 dilution of rabbit anti-Mouse IgG (whole molecule)-Peroxidase (Sigma-Aldrich Inc., Missouri, USA) was used as secondary antibody. Sections were then incubated for 1 h with above secondary antibody. 3,3'-Diaminobenzidine (DAB) substrate was used to detect the viral antigen in tissue sections, which yielded brown-black stain (Zhongshan Golden Bridge Biotechnology CO., Ltd, Beijing, China). This was followed by a light hematoxylin counterstain, after which the sections were washed in water, dehydrated and mounted with neutral balsam. The labeling procedure for negative control sections was the same as described above except above the primary antibody was substituted with PBS.
Scanning Electron Microscopy (SEM): The plicae of bursa of Fabricius were fixed overnight at 4°C with 2.5% glutaraldehyde in PBS, postfixed with 1% osmium tetroxide. After dehydration in a graded series of ethanol, samples were treated with isoamyl acetate overnight. Then specimens were dried in hexamethyl disilizane, sputter-coated with gold and examined and photographed with a Hitachi S-3400N Scanning Electron Microscope operated at 15 kV.
Transmission Electron Microscopy (TEM): The tissue samples (1 mm thick) of bursa of Fabricius at 3 and 6 days p.i. were fixed overnight at 4°C with 2.5% glutaraldehyde in PBS, gently rinsed in the 0.1 M phosphate buffer (pH 7.2), postfixed with 1% osmium tetroxide and rinsed thoroughly in the same buffer again. Subsequently they were dehydrated in a graded acetone series, embedded in SPURR resin after tissue processing and polymerized at 60°C for 2 days. Then ultrathin sections (70 nm) were prepared, stained with uranyl acetate and counterstained with lead citrate and examined under TEM (JEM-1230, Japan) operating at 80 KV and photographed.
Flow cytometric analysis of apoptosis in bursal lymphocytes: At 2, 5 and 10 days p.i., one pool containing 3 chickens per group were examined. Bursae were excised and single-cell suspensions of bursal tissues were separately prepared at the times indicated following IBDV infection. Briefly, the organs were overturned, the cells were gently scraped with a slide, suspended in RPMI 1640 supplemented with 2% FBS, filtered through 100-μm nylon mesh and centrifuged at 300 g for 5 min at 4°C. The cell pellet was resuspended in RPMI 1640 and separated in a discontinuous density gradient of culture medium and lymphocyte separation medium (Tianjin Haoyang Biological Manufactures CO., Ltd, Tianjin, China) and recentrifuged. The resulting cell pellet was resuspended in RPMI 1640 and the cell concentration was determined using a hemacytometer. To analyze cells for apoptosis the Annexin V-EGFP Apoptosis Detection Kit (KeyGEN, Nanjing, China) was employed and cell labeling was performed following the manufacturer’s instructions. The lymphocyte cell pellet was resuspended in 500 μL binding buffer. 1x106 cells were incubated with Annexin V-EGFP and Propidium Iodide (PI) at room temperature for 5 min. Positive cells stained were analyzed by EPICS ELITE (Coulter, America) and the software. Viable lymphocytes were gated on the basis of forward and side scatter characteristics and 10,000 events were analyzed for positive staining with EGFP and PI.
Statistical analysis: Statistical significance of differences among three groups in mean body weight gain and bursal weight index were calculated using Duncan’s multiple comparisons. The level of significance was set at p<0.05. Data were expressed as Mean±SD. SAS V.8.02 (SAS Institute, Cary, NC) was used to perform all calculations.
RESULTS AND DISCUSSION
Clinical signs and gross pathological changes: The groups of chickens inoculated with BC6/85 strain developed clinical symptoms including depression, anorexia, ruffled feathers, whitish or watery diarrhea and prostration, starting approximately at 4 days postinoculation. Indeed, the vIBDVs produce disease signs similar to conventional type 1 infection, with the same incubation period (4 days), but the acute phase is exacerbated and more generalized in the affected flock. As shown in Fig. 1a and b, the inoculation of chickens with BC6/85 strain caused reduced body weight gain in both the infected chickens (Group I) and the infected chickens after administration (Group II) as compared with the controls.
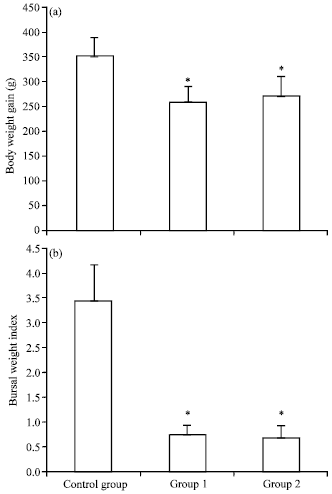 | |
| Fig. 1: | Comparison of (a) body weight gains and (b) bursal weight indices between groups by the end of experiments. Control group was a mock-inoculation group; Group I was an inoculated group; Group II was an inoculated group after administration. The results presented are the mean of nineteen chickens of each group (n = 19) ±SD. Asterisks indicate statistically significant differences between virus-free and virus-exposed groups (p<0.05) |
The body weight gain and bursal weight index of IBDV-exposed chickens were significantly different (p<0.05) from mock-inoculation controls by the end of experiments as indicated in Fig. 1. IBDV-exposed birds showed macroscopical lesions such as ecchymotic hemorrhage in the thigh muscles and the juncture of glandular and muscular stomach, branched hemorrhage in liver and bursal atrophy (Fig. 2a-d).
Histopathological changes: Experimental infection of susceptible chickens induced typical acute bursitis of various severity, and microscopic lesions in thymus, spleen, caecal tonsils, Harderian gland and liver (no shown). Histopathological changes were examined from all tissue sections after haematoxylin and eosin staining. Figure 3a is the microscopical picture of the bursa collected from the control bird. It revealed epithelial lining covering large polyhedral bursal follicles. The bursal follicle contained normal population of lymphoid cells in the cortex and medulla. Figure 3b revealed multifocal follicular atrophy of bursa from IBDV-exposed bird and extensive lymphoid damages and depletion of bursal follicles. The follicle was rounded, the interfollicular space was widened. A few follicles also revealed cellular debris in the medullary area. Besides the above histopathological changes, extensive necrosis and numerous apoptotic bodies of the BF follicles, eosinophilic cystic cavities in the medullary areas, the indistinct boundary between cortex and medulla, mild or severe increase in interfollicular stromal tissue were observed on Fig. 3c and d. But, Bursal follicles in the control bird were intact (Fig. 3a).
Immunolabelling of viral antigen: Bursal tissue sections at 3, 6, 9, 15 and 18 days postinoculation were examined. Immunohistochemistry (IHC) demonstrated abundant viral antigen in the BF at 3 and 6 days p.i. (Fig. 4a, b).
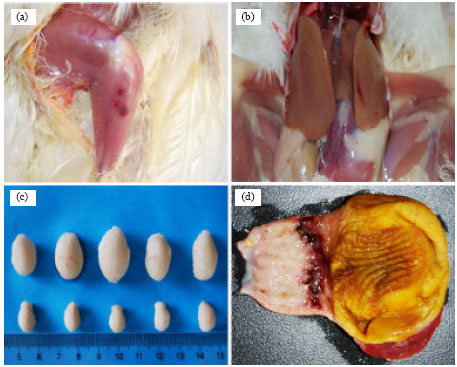 | |
| Fig. 2: | Histopathology of SPF chickens after vIBDV exposure. (a) Ecchymotic hemorrhage in the thigh muscles, (b) branched hemorrhage in the liver, (c) bursal atrophy in the exposed chickens as compared with the controls and (d) ecchymotic hemorrhage in the juncture of glandular and muscular stomach |
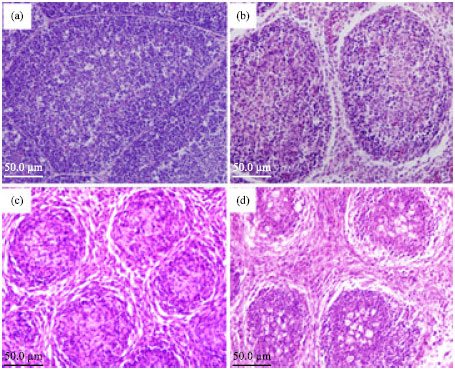 | |
| Fig. 3: | Effect of IBDV on bursae (H and E staining). At 2, 4, 8 days p.i., bursal sections from virus exposed chickens (b, c and d) and a control chicken (a) were examined. (b) The detectable changes were mild atrophy of lymphoid follicles and slight depletion of lymphocyte of the bursa of Fabricius at 2 days p.i. At 4 days p.i., the bursa revealed significant atrophy of lymphoid follicles with inflammatory lesions, severe lymphocyte depletion in almost all the follicles and the indistinct boundary between cortex and medulla. (c) There was lymphocytic necrosis, apoptosis and infiltration of heterophils in the bursal follicle and interfollicular stroma. At 8 days p.i., besides the above microscopic lesions, lymphocyte depletion became more pronounced, there were cellular debris and cystic cavities in the follicular medulla. (d) The bursal follicles got round, the interfollicular spaces were widened and filled up by fibrous tissue. (a) Bursal follicles in the control chicken were intact. 400x |
In contrast, less viral antigen was seen at 9 days p.i. (Fig. 4c); viral antigen could nearly not be viewed at 18 days p.i. (Fig. 4d). Many positive signals were shown in the cortical and medullary areas of bursal follicles (Fig. 4a-c). Various degrees of damage of the Follicle-Associated Epithelium (FAE) of the bursa of Fabricius could be observed, this damages made some bursal follicles connect directly with the bursal lumen and so many positive signals of viruses presented in these sites (Fig. 4a).
SEM observations of surface structure of bursal plicae: The mucosal surface of the bursal plica viewed through a SEM was erosion, ulceration and became smooth. The erosion of the plica mucosa at 6 days p.i. (Fig. 5a) was more serious than that at 3 days p.i. (data not shown). The reticular fibers were exposed after exuviation of epithelial cells and other cells (Fig. 5b). The normal mucosa of the plica was uneven, with button-like protruding and infolding follicular apices (Fig. 5c). There were numerous long, irregular microvilli on the epithelial surface of bursal plica after magnification (Fig. 5d).
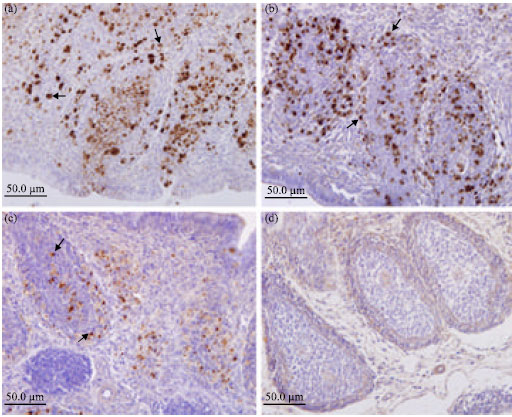 | |
| Fig. 4: | Immunohistochemical staining of IBDV-exposed bursae used to detect viral protein. At 3, 6, 9 and 18 days p.i., bursal sections from IBDV-exposed chickens were stained with anti-IBDV mAb and hematoxylin. The results revealed the presence of viral proteins (a, b and c) in the virus exposed chicken (brown-black areas shown by arrows). Viral proteins could nearly be viewed at 18 days p.i. (d). 400x |
TEM examination: The sequential morphological events of the bursa of Fabricius were examined by transmission electron microscope in SPF chickens experimentally infected with BC6/85. At 3 days p.i., the bursal epithelium and the underlying tunica propria could be observed; a few apoptotic stromal cells and necrotic cells were noted; many vacuolated inclusions were showed in the apoptotic cell (Fig. 6a, b). The striking ordered pattern of viral particles with cellular debris in the cytoplasm of a necrotic cell in the follicle could be seen, which is similar to the crystalline structure of various viruses seen intracellularly (Fig. 6c), the viral particle (virion) is spherical in shape, not surrounded by a membrane, the outer surface of virus is rough and uneven. Whereas a mature lymphocyte was not infected by IBDV, showing normal cytoplasmic organelles, for example, three mitochondria and a few rough endoplasmic reticula in Fig. 6e and f. cristae of mitochondria could be clearly found in higher magnification (Fig. 6f). At 6 days p.i., necrotic cells and apoptotic cells could be viewed and a macrophage was also found (Fig. 6d).
Time course of endogenous activation of bursal lymphocyte apoptosis: To quantitate the frequency of apoptotic cells, the lymphocyte cell suspensions were incubated with EGFP-labeled annexin V and PI. Using this flow cytometric technique, bursal lymphocyte apoptosis was examined after 2, 5 and 10 days of postinoculation. Fig. 7 shows bivariate annexin V-EGFP/PI analyses of such cell suspensions either not subjected to exposition (control, Fig. 7a) or exposed for 2, 5, or 10 days (Fig. 7b-d). Apoptotic cells appeared in the annexin V+/PI- fraction, whereas cells damaged by scraping appeared in the annexin V-/PI+ fraction.
 | |
| Fig. 5: | Mucosae of bursal plicae under SEM observation. (a) SEM shows smooth mucosal surface as compared with normal mucosa and erosion, ulceration in mucosa of bursal plica at 6 days p.i., larger than that at 3 days p.i. (not shown); (b) Exuviation of epithelial cells and other cells with the exposure of reticular fiber, specially that at 6 days p.i.; (c) The normal mucosa of the plica is intact. The plical surface with protruding follicular apices can be viewed; (d) Long, irregular microvilli of epithelial surface is clearly shown after magnification. Left lane 70x, right lane, higher magnification (5, 000x) of the same tissue sections |
The number of annexin V+/PI- cells increased with inoculation days, while the number of annexin V+/PI+ cells was the most at 5 days postinoculation. The latter fraction comprised the necrotic cells and the secondary necrotic cells, i.e., apoptotic cells, which lost their membrane integrity. As depicted in Fig. 7, apoptotic cells can clearly be visualized in the bivariate annexin V-EGFP/PI analysis. Undamaged cells remained negative for both parameters.
A Chinese IBDV strain BC6/85, a standard classic virulent IBDV strain, was chosen as the challenge virus in our experiments. BC6/85 strain is widely used in the evaluation of the efficacy of IBDV vaccines in China. In recent years, several techniques have been useful for detecting and identifying IBDV, including the reverse transcriptase-polymerase chain reaction (RT-PCR) for amplifying variable or hypervariable regions of VP2 gene and restriction fragment length polymorphism (RFLP) analysis of the amplified product (Gomes et al., 2005). The nucleic acid was extracted directly from virus particles in the vials by using the TRIzol Reagent in our experiments. This was done to eliminate the possibility of causing genetic mutations in the viruses that could occur with passage in a host (Jackwood and Sommer, 2002). RNA samples of BC6/85 strain were amplified using a modified version of a random PCR protocol (Lin et al., 1993). A 259-bp fragment and a 648-bp fragment in the VP2 and VP5 genes were amplified by RT-PCR. BC6/85 strain was found to be different from others through multiple nucleic acid or amino acid sequences alignment with the Genbank database.
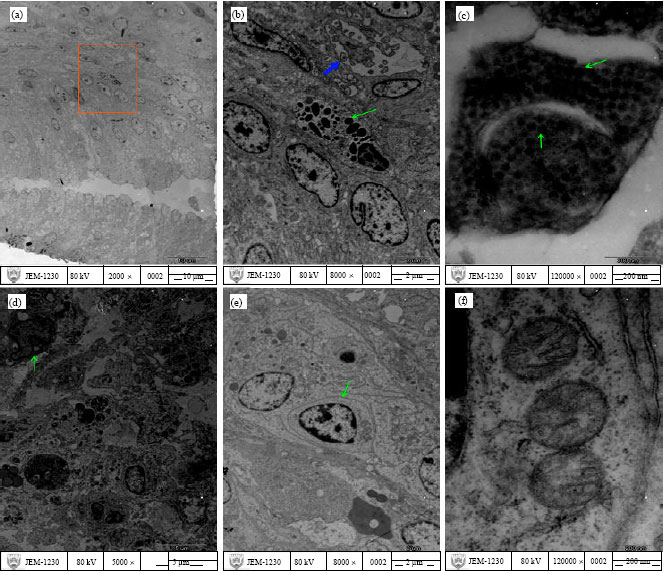 | |
| Fig. 6: | Transmission electron micrographs of the bursal plica. (a) The mucosal surface of the bursal plica and the plical cavity, (b) Higher magnification of (a), a necrotic cell (indicated by arrowheads) and a stromal cell in an early phase of apoptosis with condensed and peripheralized chromatin (indicated by arrow), many inclusions observed in the apoptotic cell, (c) IBD virus particles (arrow) showing a crystalline array in the cytoplasm of a necrotic cell in the follicle, (d) A macrophage (arrow) and many necrotic cells and apoptotic cells, (e) a mature lymphocyte (arrow) and (f) three normal mitochondria and a few rough endoplasmic reticula |
BC6/85 strain was pathogenic for SPF White Leghorn chickens when applied by the eye drop route. The clinical signs observed in IBDV-exposed chickens, macroscopic lesions and histopathologic changes in the bursa, are in agreement with findings by other authors (Sharma et al., 2000; Banda et al., 2003). BC6/85 strain was characterized by the induction of depression, anorexia, ruffled feathers, whitish or watery diarrhea, prostration and little or no mortality.
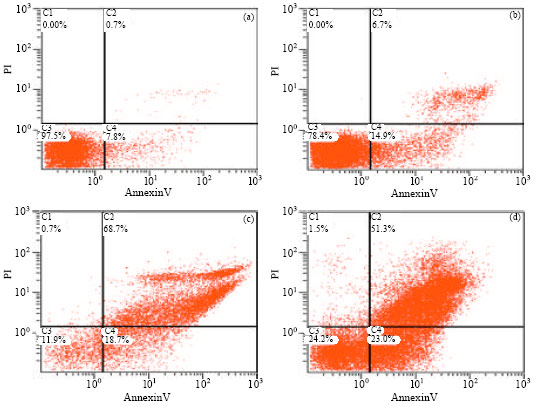 | |
| Fig. 7: | Flow cytometric analyses of single bursa cell suspensions at the times revealed that necroses and apoptosis at 2, 5 and 10 days after treatment of virus-exposed (b, c, d) or virus-free (a) bursal lymphocytes. The minimal apoptotic staining of lymphocytes in the bursas of unexposed birds represents normal cellular turnover during maturation of lymphocytes. (a) (F1)(A) 1.LMD: PMT2 log/PMT4 log, (b) (F1)(A) 2.LMD: PMT2 log/PMT4 log, (c) (F1)(A) 5.LMD: PMT2 log/PMT4 log and (d) (F1)(A) 8.LMD: PMT2 log/PMT4 log |
In addition, the exposed birds exhibited a reduction in body weight gain along with poor flock uniformity. The distinctive lesion was the presence of bursitis with further bursal atrophy. Upon necropsy, varying degrees of hemorrhages could be found in the thigh muscles and the juncture of glandular and muscular stomach, along with classic changes to the bursa: atrophy and gelatinous coating. But no hemorrhage could be found in pectoral muscles. Our experiment could not identify the causal effect of this. Histologically, considering the character of the infection and the affinity of the causative agent to immature precursors of B lymphocytes particularly in bursa and to a lesser extent also in the thymus, spleen, caecal tonsils and Harderian gland, the emphasis of our experiments was put on the bursa serving as targets for the virus. The detectable changes were atrophy of lymphoid follicles and depletion of lymphocyte of the bursa of Fabricius. As the infection advanced, the lesions showed more severe changes. Lymphocyte depletion with apoptosis and necrosis in lymphoid tissues became more pronounced, as cystic cavities and necrotic cells were observed in the follicular medulla and the structure of the follicles was dissolved, similar to the lesions described in chickens by Sharma et al. (1993). The combination of light microscopy and TEM showed that necrotic and apoptotic cells were seen predominantly in the medulla of the follicles in the early stages of viral infection, so we speculate that immature B lymphocytes in the medulla are first infected by IBDV, then reticulum cells or others are infected; Later, some cells in the cortex are infected. Whereas all mature lymphocytes in the cortex and medulla seem not be infected, as shown in Fig. 6e, the mature lymphocyte and cytoplasmic organelles were normal. Furthermore, the viruses may also replicate in other cells, immature B cells may not be the sole target for the virus; macrophages, reticulum cells and possibly other cells may serve as host for IBDV (Khatri et al., 2005). Figure 6b shows many inclusions in the cytoplasm of the stromal cell showing typical signs of apoptosis (margination of chromatin). These inclusions may be formed by virus particles, cytoplasmic membrane and other cellular components either when viruses replicate in host cells or after viruses are phagocytized by phagocytes. The damage of the cells observed in the bursa of Fabricius at 6 days p.i. was more severe than those at 3 days p.i. (Fig. 6b, d). In addition, probably there exists an inestimable possibility that the viruses may replicate in stromal cells within the medulla of lymphoid follicles, immature B lymphocytes may be affected simultaneously. As B lymphocyte development and differentiation depend upon the endogenous microenvironment, stromal cells may be involved in crucial interactions and close association with developing B cells, the progressive loss of the bursal stromal component could have resulted in the overall depletion of B cells. Therefore, we infer that the bursal stroma does not have the capacity to support normal lymphocyte development if bursal stromal cells are not available as a result of the stromal cell damages caused by IBDV, eventually, leading to the destruction of the general histologic structure of the bursa of Fabricius. Viral crystalline array in the cytoplasm of a necrotic cell in the follicle under transmission electron microscope was also observed (Fig. 6c).
Viral protein was detected by IHC in bursal tissues collected at 3, 6, 9 and 15d days postinoculation. Whereas viral protein could rarely be detected at 15 days p.i. (data not shown) and not nearly be found 18 days p.i. (Fig. 4d). The presence of viral antigen was confirmed by qualitative immunohistochemical analysis. A recent work of ours, using histopathological, immunohistochemical and SEM methods, clearly showed that the follicle-associated epithelial cells are affected by IBDV, while the surrounding epithelial cells do not show some visible signs of damages (Fig. 4a). After the viral antigen was positioned using immunohistochemistry, we noted that some follicles were directly connected with the bursal lumen as the Follicular Associated Epithelium (FAE) may be damaged. Therefore, we suspect that the erosion of the mucosal surface first occurred in these sites seriously damaged.
Because many authors having studied avian intraorbital glands (Ballantyne and Fourman, 1967; Burns, 1976; Payne, 1994; Shirama et al., 1996; Dimitrov and Nikiforov, 2005), report that the Harderian gland has a better developed and branched blood supply that could be a favorable factor for the spreading of the infectious agents in the organ, we speculate that IBDV may first reach the Harderian gland after intraocular inoculation, then the virus gets to the bursa of Fabricius and other organs via the blood stream. The virus may replicates primarily in the lymphocytes and macrophages of the BF, is then released into the blood stream, travels to other tissues or organs again, where replication will occur. After the second round of viral replication, stromal cell and possibly others cells in the bursa of Fabricius may be damaged, the necrosis and apoptosis of cells will occur.
The Annexin V/PI flow cytometry assay was used to detect cells population in viable, early apoptosis and late apoptosis (necrosis) stages. The cells were stained with EGFP conjugated Annexin V and PI and they were subjected to flow cytometry. Cells undergoing early and late apoptosis were detected by the emission of the fluorescence from both EGFP and PI, respectively. Our data showed inoculation with BC6/85 strain to SPF chickens results in apoptosis and necrosis of target cells, detected at 2 days p.i., with a maximum apoptosis + necrosis being observed at 5 days p.i., with a maximum apoptosis at 10 days postinoculation during the experiments. These results are consistent with earlier research by Kim et al. (2000), by 7 days p.i., the proportion of B cells in the bursa of IBDV-exposed chickens had dropped to the lowest; the numbers of both CD4+ and CD8+ T cells reached peak levels. We also noted that there was a minor difference in time, when viable lymphocytes in the bursa of IBDV-exposed chickens had dropped to the lowest. This may correlate with the virulence of IBDV, the age of chicken. Exposure to IBDV resulted in infiltration of T cells into the bursa and intrabursal T cells of IBDV-exposed chickens were activated. T cells may be involved in antiviral immune responses. The results shed new light on the importance of cell-mediated immunity in the pathogenesis of IBDV, a naturally occurring immunosuppressive virus of chickens that causes a lytic infection in B cells. Although T-cell immunity may be important for host defense in IBD, the possibility cannot be excluded that IBDV-induced T cells may exacerbate bursal lesions. Cytotoxic T cells may promote lysis of virus-exposed bursal cells. Alternatively, an influx of proinflammatory cytokines may enhance tissue destruction (Kim et al., 2000). Liu and Vakharia (2006) demonstrated that IBDV-induced apoptosis is caspase dependent and activates caspases 3 and 9. Nuclear factor kappa B (NF-κB) is also activated and is required for IBDV-induced apoptosis.
Further studies are needed to identify the cells involved in the pathogenesis of IBDV and their functions and establish the exact role of viral proteins in pathogenesis and immunosuppression, notably by comparing them in strains with different virulence.
ACKNOWLEDGMENTS
The authors would like to thank Dr. Qinfang Liu for his excellent technical assistance in our experiments and Dr. Honglei Gao for his generous present of monoclonal antibody.
REFERENCES
- Banda, A., P. Villegas and J. El-Attrache, 2003. Molecular characterization of infectious bursal disease virus from commercial poultry in the United States and Latin America. Avian Dis., 47: 87-95.
PubMedDirect Link - Becht, H. and H. Muller, 1991. Infectious bursal disease-B cell dependent immunodeficiency syndrome in chickens. Behring Inst. Mitteilungen, 89: 217-225.
PubMedDirect Link - Burns, R.B., 1976. The structure of the lacrimal glands of the domestic fowl and of the duck. Res. Vet. Sci., 21: 292-299.
PubMedDirect Link - Dimitrov, D.S. and I.P. Nikiforov, 2005. Histological and histochemical studies of Harderian gland, lacrimal gland and bursa of Fabricius in mulard ducks (Anas sterilis) with chlamydial infection. Bulgarial J. Vet. Med., 8: 119-127.
Direct Link - Gomes, A.D., J.T. Abreu, R.A.F. Redondo, N.R.S. Martins, J.S. Resende and M. Resende, 2005. Genotyping of infectious bursal disease virus strains by restriction fragment length polymorphism analysis of the VP1, VP2 and VP3 genes. Avian Dis., 49: 500-506.
CrossRefDirect Link - Harris, J.C., S.L. Cottrell, S. Plummer and D. Lloyd, 2001. Antimicrobial properties of Allium sativum (garlic). Applied Microbiol. Biotechnol., 57: 282-286.
CrossRefDirect Link - Jackwood, D.J. and S.E. Sommer, 2002. Identification of infectious bursal disease virus quasispecies in commercial vaccines and field isolates of this double-stranded RNA virus. Virology, 304: 105-113.
CrossRef - Jungmann, A., H. Nieper and H. Muller, 2001. Apoptosis is induced by infectious bursal disease virus replication in productively infected cells as well as in antigen-negative cells in their vicinity. J. Gen. Virol., 82: 1107-1115.
Direct Link - Khatri, M., J.M. Palmquist, R.M. Cha and J.M. Sharma, 2005. Infection and activation of bursal macrophages by virulent infectious bursal disease virus. Virus Res., 113: 44-50.
PubMedDirect Link - Kibenge, F.S., A.S. Dhillon and R.G. Russell, 1988. Biochemistry and immunology of infectious bursal disease virus. J. Gen. Virol., 69: 1757-1775.
Direct Link - Kim, I.J., S.K. You, H. Kim, H.Y. Yeh and J.M. Sharma, 2000. Characteristics of bursal T lymphocytes induced by infectious bursal disease virus. J. Virol., 74: 8884-8892.
Direct Link - Lin, Z., A. Kato, Y. Otaki, T. Nakamura, E. Sasmaz and S. Ueda, 1993. Sequence comparisons of a highly virulent infectious bursal disease virus prevalent in Japan. Avian Dis., 37: 315-323.
PubMedDirect Link - Liu, M.H. and V.N. Vakharia, 2006. Nonstructural protein of infectious bursal disease virus inhibits apoptosis at the early stage of virus infection. J. Virol., 80: 3369-3377.
CrossRefPubMedDirect Link - Muller, H., M.R. Islam and R. Raue, 2003. Research on infectious bursal disease-The past, the present and the future. Vet. Microbiol., 97: 153-165.
CrossRefPubMedDirect Link - Nieper, H., J.P. Teifke, A. Jungmann, C.V. Lohr and H. Muller, 1999. Infected and apoptotic cells in the IBDV-infected bursa of Fabricius, studied by double-labelling techniques. Avian Pathol., 28: 279-285.
CrossRefDirect Link - Payne, A.P., 1994. The harderian gland: A tercentennial review. J. Anat., 185: 1-49.
PubMedDirect Link - Ross, Z.M., E.A. O'Gara, D.J. Hill, H.V. Sleightholme and D.J. Maslin, 2001. Antimicrobial properties of garlic oil against human enteric bacteria: Evaluation of methodologies and comparisons with garlic oil sulfides and garlic powder. Applied Environ. Microbiol., 67: 475-480.
CrossRefDirect Link - Uddin, S.N. and S.A. Hossain, 2006. Effect of infectious bursal disease virus on in vitro propagation of chicken embryo fibroblast cells. AJAVA 1: 55-59.
CrossRefDirect Link - Schutte, B., R. Nuydens, H. Geerts and F. Ramaekers, 1998. Annexin V binding assay as a tool to measure apoptosis in differentiated neuronal cells. J. Neurosci. Methods, 86: 63-69.
CrossRefDirect Link - Sharma, J.M., J. Dohms, M. Walser and D.B. Snyder, 1993. Presence of lesions without virus replication in the thymus of chickens exposed to infectious bursal disease virus. Avian Dis., 37: 741-748.
PubMedDirect Link - Sharma, J.M., I.J. Kim, S. Rautenschlein and H.Y. Yeh, 2000. Infectious bursal disease virus of chickens: Pathogenesis and immunosuppression. Dev. Comp. Immunol., 24: 223-235.
CrossRefPubMedDirect Link - Shirama, K., T. Satoh, T. Kitamura and J. Yamada, 1996. The avian harderian gland: Morphology and immunology. Microscopy Res. Tech., 34: 16-27.
PubMedDirect Link - Tanimura, N. and J.M. Sharma, 1998. In-situ apoptosis in chickens infected with infectious bursal disease virus. J. Comp. Pathol., 118: 15-27.
PubMedDirect Link - Tanimura, N., K. Tsukamoto, K. Nakamura, M. Narita and M. Maeda, 1995. Association between pathogenicity of infectious bursal disease virus and viral antigen distribution detected by immunohistochemistry. Avian Dis., 39: 9-20.
CrossRefPubMedDirect Link - Taylor, R.C., S.P. Cullen and S.J. Martin, 2008. Apoptosis: Controlled demolition at the cellular level. Nat. Rev. Mol. Cell Biol., 9: 231-241.
CrossRefPubMedDirect Link - Van den Berg, T.P., 2000. Acute infectious bursal disease in poultry: A review. Avian Pathol., 29: 175-194.
CrossRefDirect Link - Ahmed, Z. and S. Akhter, 2003. Role of maternal antibodies in protection against infectious bursal disease in commercial broilers. Int. J. Poult. Sci., 2: 251-255.
CrossRefDirect Link