
Bo Yu
Department of Animal Genetics and Breeding, College of Animal Science and Technology, China Agricultural University, Yuanmingyuan West Road No. 2, Haidian District, Beijing, 100193, People`s Republic of China
Huojun Cao
Department of Animal Genetics and Breeding, College of Animal Science and Technology, China Agricultural University, Yuanmingyuan West Road No. 2, Haidian District, Beijing, 100193, People`s Republic of China
Chunjiang Zhao
Department of Animal Genetics and Breeding, College of Animal Science and Technology, China Agricultural University, Yuanmingyuan West Road No. 2, Haidian District, Beijing, 100193, People`s Republic of China
Changxin Wu
Department of Animal Genetics and Breeding, College of Animal Science and Technology, China Agricultural University, Yuanmingyuan West Road No. 2, Haidian District, Beijing, 100193, People`s Republic of China
Xuemei Deng
Department of Animal Genetics and Breeding, College of Animal Science and Technology, China Agricultural University, Yuanmingyuan West Road No. 2, Haidian District, Beijing, 100193, People`s Republic of China
Asian Journal of Animal and Veterinary Advances
Year: 2011 | Volume: 6 | Issue: 3 | Page No.: 242-254







ABSTRACT
Map-based positional cloning of Drosophila melanogaster genes is hampered by both the time-consuming, labor-intensive, error-prone nature of traditional methods for genetic mapping-SNP mapping is costly and P element mapping is low-resolution for limit number of P elements. Here we used two strategies to locate a spontaneous mutant screened in our lab. One is P element mapping to map the mutant in a 33 k-interval on the right arm of the second chromosome; another is combination of P element mapping and a Single-Nucleotide Polymorphism (SNP) mapping together to map the mutant in a very small interval, about 5 kb, on the right arm of the second chromosome efficiently. At last, we guessed that the mutant phenotype maybe resulted from the overexpression of twist gene. Comparing the two strategies, the method combining the P insertion mapping and SNP mapping could ward off their drawbacks. Therefore, the strategy of combination of the two methods provides a rapid protocol for mapping mutations.
PDF Abstract XML References Citation
Received: March 21, 2010;
Accepted: May 06, 2010;
Published: July 10, 2010
How to cite this article
Bo Yu, Huojun Cao, Chunjiang Zhao, Changxin Wu and Xuemei Deng, 2011. A Rapid Way to Map Outheld-Wing Mutation in Drosophila melanogaster. Asian Journal of Animal and Veterinary Advances, 6: 242-254.
DOI: 10.3923/ajava.2011.242.254
URL: https://scialert.net/abstract/?doi=ajava.2011.242.254
DOI: 10.3923/ajava.2011.242.254
URL: https://scialert.net/abstract/?doi=ajava.2011.242.254
INTRODUCTION
Drosophila melanogaster plays an important role in the field of forward genetics over one hundred years, partially due to its advantages over other organisms, especially performing large-scale genetic screens of mutations by X- or γ-ray, chemical mutagen and spontaneousness that affect a given biological process. Genetic mapping of natural or induced genomic variation remains a powerful approach to understand the function of genes in a variety of biological processes (Baird et al., 2008). However, mapping and identification of the mutant with an interesting phenotype are often difficult and labor-intensive. So up to now, many methods have been developed and available to map a mutant locus, such as meiotic recombination mapping with dominant markers, male recombination mapping (Chen et al., 1998; Duttaroy, 2002), deficiency mapping (Littleton and Bellen, 1994; Zraly et al., 2002), SNP mapping (Wang et al., 1998; Berger et al., 2001) and P element mapping (Zhai et al., 2003). Moreover, both of SNP mapping and P element mapping are high-resolution and high-throughput methods and also can accelerate the mapping of given mutations.
With the completion of the fly genome (Adams et al., 2000), it is an easy work to cast about for more precisely mapped markers based on Single-Nucleotide Polymorphisms (SNPs). The SNPs are DNA sequence variations between paired chromosomes in an individual, including base-pair changes and single-nucleotide insertion or deletion. The SNP mapping is used to map a mutant locus in a two-step process (Martin et al., 2001; Nairz et al., 2002). First is establishing a SNP maps between two different genetic backgrounds. Second is genotyping with established SNP markers. Therefore£¬detecting SNPs in the mutant strains should be undergone meiotic recombination. SNPs can be detected by PCR-RFLP, PCR-PLP (Berger et al., 2001), PCR-SSCP, DHPLC (Hoskins et al., 2001; Underhill et al., 1997) or sequencing. Up until now, several publications have reported the successful application of the method in a few cases (Johnson et al., 2001; Martin et al., 2001; Wang and Todd, 2003; Macdonald et al., 2005), which demonstrate SNP mapping is a powerful and high-resolution method. However, SNP mapping requires large amounts of molecular biology reagents and analyses, which will be outstanding, if a team wants to construct a high-throughput SNP maps.An alternative method of SNP mapping is P element mapping. Since, first constructed as a tool for transgenesis in 1982 (Rubin et al., 1982), P element plays a more and more important role in fly genetics. A variety of systems based on the transposon have provided a range of tools for genetics and genomics application, especially in the reverse genetics, such as gene trapping (Sanyal, 2009), the GAL4-UAS system (Mondal et al., 2007), site-specific recombination using FRT sites (Theodosiou and Xu, 1998), gene targeting (Osamu et al., 2009) and so on. In the forward genetics, P element has an extensive application in mapping a mutation. Zhai et al. (2003) demonstrated that P element mapping, as SNP mapping, is also a high-resolution and high-throughput method if the large collection of molecularly defined P element insertions are available in a specific region and pointed out that theoretically, via using two molecularly mapped P elements, a mutation could be mapped to a single nucleotide. Two rounds should be required, when P element mapping is taken into account to map a mutation on the known chromosome (Zhai et al., 2003). The first round is rough mapping to around 500 kb and the second round is fine mapping to narrow the interval within 10-60 kb. Subsequently, various methods can be selected in the next step to further delimit the candidate region, for example candidate gene selection, complementary test using smaller deficiencies or TGCE to detect point mutation. The accuracy of the method lies in aligning the genetic and cytological maps with the genome sequence. However, owing to P element has a bias to fly genome (O’ Hare and Rubin, 1983), the density of P elements will be low as to cold spots for P element insertion. Therefore, P element’s preference is becoming the drawback of this mapping method.
Just as mentioned above, SNP mapping and P element mapping both have advantages and disadvantages, SNP mapping is high-resolution but costly, while P element mapping is cheap and easy but constricted by the limit numbers of P elements. Here, we combined SNP mapping and P element mapping together, warding off their drawbacks, to map the spontaneous mutation screened in our lab, beijing spreding wing (bsw), in a very narrow interval on the right arm of the second chromosome-about 5 kb, which proved that the strategy was cheap, efficient, reliable and high-resolution.
In the present study, we adopted two strategies to identify a mutant, bsw. One was P element mapping, the other was the combination of SNP mapping and P element mapping. Via comparing the two strategies, we found that the combination of the two methods is excelled to P element mapping only, which provided a rapid protocol for mapping mutation.
MATERIALS AND METHODS
Fly stocks: All fly stocks and crosses were maintained on standard yeasted cornmeal-agar food at 25°C and 50% relative humidity. Beijing spreading wings (bsw) were screened by Drosophila laboratory in College of Animal Science and Technology, China Agricultural University (CAU) in 2006. Its phenotype is as follows: the purple-eye mutant holds out its wings and the angle between the wing and the axis of the body is about 30∼90°. Meanwhile, the wings are elevated and the angle is about 30∼60°. This mutant is unable to fly, but there is no abnormality on its ability to crawl and jump. We named this mutant as beijing spreading wing (bsw). Balancer chromosomes, w; Adv/SM1; Sb/TM6, were supplied by Peking University. Drosophila Genetic Resource Center in Japan (DGRC) offered us with 37 P insertion stocks.
Five P element stocks used to do fine mapping and 33 deficiency stocks were provided by Bloomington Drosophila Stocks Center in America, a summary of the five P insertion stocks used in this study is shown in Table 1.
Detail information of all these Drosophila stocks ordered from Japan and the USA are described in Flybase (http://www.flybase.net).
Genetic methods
Chromosome mapping: The mutant (bsw) was crossed to wild-type strain, Canton Special (CS), to determine it’s a dominant or recessive gene. The females of F1 generation, bsw/+, were backcrossed to the mutant male flies and the progeny with different phenotypes were collected and counted (Fig. 1A).
In order to facilitate the next work, we first mapped the mutant site on the second chromosome using the balancer chromosome,
Female flies containing double balancer chromosomes,
were mated to the mutant male flies. In F1 generation, male flies with both phenotypes of abnormal wing vein (Adv) and short bristle (Sb) were collected and backcrossed to the mutant female flies. According to the phenotype of the progeny in F2 generation, we could confirm that which chromosome the mutant site located on (Fig. 1B).
| Table 1: | P element stocks used to fine map in this study |
 | |
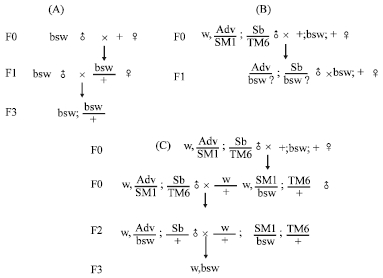 | |
| Fig. 1: | A summary of the crosses used to map the mutant loci on the chromosome level and synthesize w; bsw. Figure 1A shows the crosses between CS and bsw to determine the mutant gene is dominant or recessive. Shown in B is the process of balancer mapping. Shown in C are the process of synthesizing outheld wing and white-eye strain, w; bsw and isogenizing the second chromosome. In F2 generation, |
P element mapping: Thirty-seven P element stocks with white+ as a dominant marker were used to manage the rough mapping. Because taking mini-white as a dominant marker in this step, we had to synthesize outheld-wing and white-eye strain, w; bsw, firstly. Flies with double balancer chromosomes
were mated to the mutant flies. A virgin fly without both phenotypes of abnormal wing vein (Adv) and short bristle (Sb) collected in F1 generation backcrossed to a male fly with double balancer chromosomes and the progeny were random-mated to each other. In F3 generation, red-eye and outheld-wing flies (w; bsw) were collected (Fig. 1C). Meanwhile, this step was also an isogenizing process.
P element mapping has two rounds: the first round is rough mapping and the second is fine mapping. The schematic diagram of P element mapping principle is shown in Fig. 2, the homozygous mutant allele was crossed to a heterozygous w+-marked P insertion strain balanced by a balancer chromosome with Cy. In F1 generation, female flies backcrossed to the male mutant flies and the progeny’s phenotypes were observed (Fig. 2A). If no chromosome exchange occurred, white-eye, outheld-and elevated-wing flies and red-eye, normal wing flies were obtained (Fig. 2B); while if the recombination events happened, two recombinants were collected (Fig. 2C). The Projected Molecular Position (PMP) was calculated by using each pair of flanking P elements according to Zhai et al. (2003).
After using the P element mapping to rough map the mutant, we had two choices in the next step, one was P elements to fine map and the other was SNP mapping.
SNP mapping: We searched for sequencing polymorphisms in the interval around 59B region on the right arm of the second chromosome. We carried out PCR amplification with genomic DNA prepared from one P insertion stock with the insertion site being at 59B1-3 and the mutant, primers (0.4 μM), MgCl2 (1.5 mM), dNTPs (0.2 mM each) and Taq DNA polymerase (TakaRa, 1.25U). Cycle conditions were 94°C for 5 min, followed by 35 cycles at 94°C for 30 sec, 62°C for 30 sec and 72°C for 45 sec and a final extension at 72°C for 10 min. The products of PCR amplification were sent to SinoGenoMax Co., Ltd to carry out DNA sequencing after detecting on 1% agarose gel.
We then multiply re-aligned the sequences using the DNAMAN (Version 5.2.2) software with default parameters. We screened 55 SNPs for the presence of recognition site and selected 7 SNPs to analyzing by PCR-RFLP.
Primers used for SNPs identification and PCR-RFLP assay were designed using Primer 3 (Whitehead Institute/ MIT Center for Genome Research; http://www.genome.wi.mit.edu). We selected primers with melting temperatures of 60-70°C, maximum Tm difference of 1, length of 18-22nt, 40-60% GC content and giving a product of 700-900 bp. For RFLP assays, we digested 10 μL PCR product with the appropriate restriction enzyme in a 20 μL reaction and separated the resulting fragments on a 2% agarose gel.
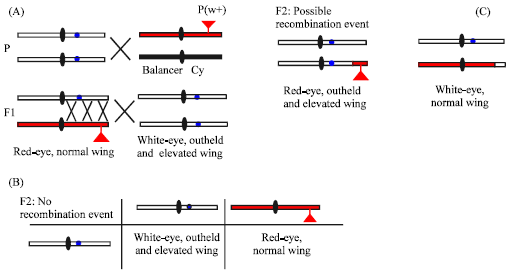 | |
| Fig. 2: | The schematic outline of the principle of P element mapping. Mutant chromosomes are indicated by open bars, P insertion-containing chromosomes are in red and the balancer chromosomes are in black. Mutant sites are marked with blue points. Note that the whole processes are under the |
We recovered recombinants between the mutant site and the P insertion site being at 59B1-3. We identified two types of recombinants by screening for red-eye, outheld wing and white-eye, normal wing. We recovered 81 recombinants from approximately 18,800 progeny.
RESULTS AND DISCUSSION
Experiment of bsw being crossed to wide type (CS) and chromosomal level mapping: Figure 1A showed that in the F1 generation, the phenotype of bsw/+ took on wild-type. And, in F2 generation, the numbers of bsw/bsw and bsw/+ were 300 and 310, respectively, which were formally non-significant (p>0.05). From the integrated process of crosses, we found there was no linkage between the mutant gene and the sex chromosome. In all, we concluded that the mutant gene, bsw, was a recessive mutant on autosome.
In Fig. 1B, the balancer chromosome stock,
were used to map the mutant gene. In F2 generation, among the flies we acquired, there was no one fly had the both phenotypes of abnormal wing vein (Adv) and outheld wing (the mutant phenotype, bsw), while flies with the phenotype of short bristle (Sb) and outheld wing could be collected. According to Mendelian inheritance, the mutant gene segregated with Adv forever, but recombined with Sb freely. The results concluded that the mutant gene was on the second chromosome.
P element mapping (the first round-rough mapping): Before managing P element mapping, we should change the eye-color background of the mutant (from purple to white), for every P element with a dominant marker, white+. Therefore, synthesizing white-eye, outheld-wing strain was necessary, which was also an isogenizing process (Fig. 1C).
Then, thirty-seven P element stocks that distributed averagely on the second chromosome were exploited to rough map the mutant gene on a known chromosome. The principle of P element mapping was outline in Fig. 2. In brief, the mutant allele was crossed to a heterozygous w+-marked P insertion strain balanced by a balancer chromosome with Cy. Recombination occurs in F1 females heterozygous for the mutant allele and the P insertion-bearing chromosome. These females were backcrossed to males homozygous for the mutant allele (Fig. 2A). Hence, in F2 generation, the progeny with white-eye, outheld wing and red-eye, normal wing would be acquired if no crossing-over occurred (Fig. 2B). Two types of phenotypes, red-eye, outheld-wing flies and white-eye, normal wing flies, were the result of recombination events between the P insertion and the mutant allele (Fig. 2C). Therefore, Recombination Rate (RR) could be calculated. Table 2 reported that when the physical sites of P element were in the interval of 21C; 38A, the Rrs between P insertion sites and the mutation were all over 30%, the minimum was 33% and the maximum was 50%. However, in the region of 43B; 60A, the trend of RRs change took on parabola curve and had a lowest point at the cytolocation of 59B, only 0.4%. While at two flanks of 59B-58D and 60A, RRs were higher, 2.7 and 2.5%, respectively. We know that each of major chromosome arms of Drosophila is divided into 20 numbered segments, 1-20 for X, 21-40 for 2L, 42-60 for 2R, 61-80 for 3L and 81-100 for 3R. Chromosome 4 in divided into regions 101-104 (Greenspan, 1997). Therefore, these data told us that the mutant site located around the region of 59B on the right arm of the second chromosome.
| Table 2: | Partial data of P element mapping |
 | |
Making a choice-the second round of p element mapping and SNP mapping
P element mapping (the second round-fine mapping): Here, we also favored P elements mapping to fine map the mutant lethal that used the same principle as for the rough mapping strategy (Fig. 2). Five P element stocks ordered from Bloomington Drosophila stocks center in America were selected to manage this strategy (Table 1). Table 1 showed that the cytolocation of the five insertion stocks distributed in the interval of 58F4; 59E2, which encompassed the mutant site. Therefore, we could locate the mutation from two flanks of 59B to further obtain the accurate results.
The five P insertion stocks used to fine mapping were mated to the mutant as the scheme outlined in Fig. 2. In F2 generation, the total numbers of the progeny and the recombinants were scored. The χ2-test indicated that the practical ratio of two types of recombinants was accorded with the theoretical ratio, 1:1. The percentage of two recombinant flies in the F2 progeny therefore represents the RD(RR%x100) in cM between the mutation and P insertion (Table 3). Because the P insertions are molecularly mapped, the molecular distance (MD in base pairs) between any two P insertions can easily be calculated from their insertion site. Therefore, for every pairs of P insertions, a Projected Molecular Position (PMP) of the mutation can be defined on the basis of the equations offered by Zhai et al. (2003) (Table 4). The PMP was calculated by using each pair of flanking P insertion. As shown in Table 1, we knew the cytolocations of the five P elements flanking the mapped-region, thus we could pair them as follows: (11217, 12194), (11217, 12195), (11217, 4399), (10471, 12194), (10471, 12195) and (10471, 4399). For the physical sites of five P insertions having been known, the Molecular Distance in base pair (MD), the Recombination Rate (RR), Projected Molecular Distance (PMD) and Projected Molecular Position (PMP) in base pair of each couple of P elements (Table 4). Table 4 showed that the PMPmax = 18895005 bp and PMPmin = 18862095 bp, so the location of the mutant could be mapped in the interval of 18862095-18895005 bp, about 33kb, which lies in the cytolocation map of 59B8;59C1-2 interval. If the RRs are similar across the region spanned by the P insertions, the PMP will be restricted to a smaller interval. As observed in our data, the RRs were similar (p>0.05), so the 33 kb-mapped interval was a very small interval and the results were accuracy.
| Table 3: | Number of recombinants in F2 generation |
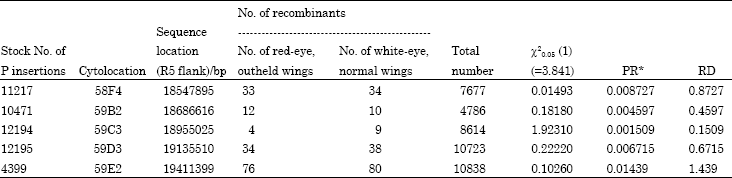 | |
| The χ2-test was used to test whether the practical ratio, No. of red-eye, outheld wings/No. of white-eye, normal wings, accorded with the theoretical ratio, 1:1. *RR = No. of recombinants/Total No. | |
| Table 4: | The results of P element mapping |
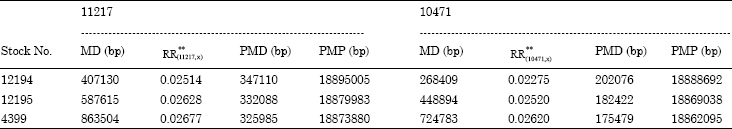 | |
| **RR indicates the recombination rate between two P elements, that is, | |
We then sequenced the region and compared it with the sequence of wild-type, however, no functional SNPs or deletions associated with the mutant phenotype were identified. We guessed that the mutant site might be located around the boundary of the region, which was confirmed by the SNP mapping. Maybe several reasons could explain this result. The first one is the number of F2 generation. According to the view of Zhai et al. (2003) if getting an accuracy of ≈ 0.1 cm, at least 10,000 F2 progeny should be collected. In present experiment, expect to F2 offspring of two stocks (stock numbers are 12195 and 4399, respectively) being over 10,000, the other three were all less 10,000. Another main reason is the translation of the recombinational map in cM into the physical map in base pair, which primarily bases on the colinearity of the two maps over short distances.
SNP mapping: Via using P element mapping to rough map the mutant, we got to know that when the cytolocation of P element was at 59B, the RR was the lowest and 81 recombinants were collected (Table 2). However, only 75 recombinants’ DNA then had been extracted for unexpected reasons, which contained 36 red-eye, outheld wings and 39 white-eye, normal wings. We amplified and sequenced PCR products approximately 800 bp in size from the P insertion stock and the mutant. We detected sequence polymorphisms and established PCR-RFLP assay for rapidly genotyping a subset of these polymorphisms that discriminate between the P insertion stock and the mutant. Seven SNPs for restriction-site polymorphisms were verified and analyzed by a restriction digest of PCR products from P insertion/mutant heterozygotes.
Table 5 showed the data of SNP mapping. Via genotyping the 75 recombinants using PCR-RFLP, we could confirm the number of progeny in which recombination events occurred before or after a given SNP. As shown in Table 5, the number of recombinants before SNP28 was 74 and only one recombinants after SNP28. Another SNP, SNP37, all of 75 recombinants were before the SNP37 and no one after the SNP37. Because SNP37 located in the site downstream of SNP28, we concluded that the mutant lied distal to SNP28 but probably not more than 5 kb from it (given the average of one recombination rate was 5 kb). Figure 3 showed that the mapped-region was upstream of twist gene (gray bar); therefore, the mutant was mapped in the region of 18927-18932 kb, which lied in 59C2-C3 interval.
The accuracy of the region then had been taken account of. As a matter of fact, with the exception to the stocks shown in Table 3, we had collected and counted the F2 progeny of another insertion stock whose stock number is 10,450 (Data were not shown in Table 3 and 4 for its physical site was unknown). The cytolocation of the stock is in 59C2-C3 interval. In F2 generation, 5,783 progeny were collected and only one of which was red-eye, outheld-wing. The data told us that the cytolocation of this stock was very near the lesion site. The result may be also as an evidence to prove the exact and precise of the strategy by combining the two methods.
| Table 5: | Data of SNP mapping |
 | |
| *As for SNP13, we had analyzed 33 recombinants, not all of 75 recombinants. NA: Not available data | |
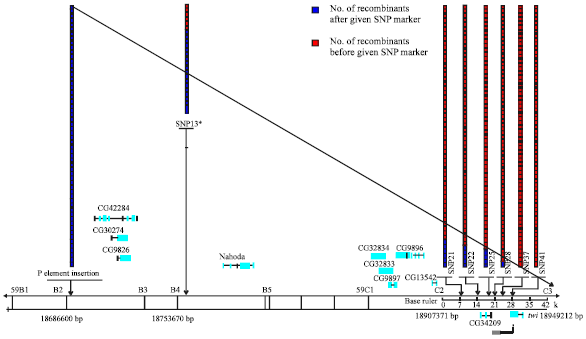 | |
| Fig. 3: | The results of high-resolution SNP mapping. Boxes indicate SNP genotypes determined for each recombinant (rows) at the indicated SNP marker (columns). The gray bar at the bottom indicates the mapped interval and the gray broken line shows the unknown distal breakpoint. *SNP13, as for this SNP marker, not all recombinants were analyzed, just only 33 recombinants |
Besides, using P insertions to rough map the mutation fist, we could narrow the interval and further low the cost of SNPs identification and genotyping. In present experiment, sufficient numbers of SNPs were identified (1 SNP per ≈ 5 kb), so mapping to either side of individual SNPs could provide the resolution necessary to map to a single gene.
When sequencing the 5 k-regions, we found that a 4,414 bp-sequence had inserted in the site of 18,929,626 bp. Via bioinformatics analysis, we found that the reverse complementary sequence of the inserted fragment had the retrotransposon features of gypsy/Ty3 subfamily: a pair of long terminal repeats (LTR, 97.8%) and two internal portion of Pifo_I retrovirus-like elements (ENV protein: 97.6%; portion of GAG protein: 99.6%), but POL protein was completely lost. The insertion site located in the intergenic spacer region and its two flanking genes were upstream CG34209 and downstream twist, respectively. Therefore, twist, which plays a very important role in the development of muscular progenitor cells (Bate et al., 1991; Baylies and Bate, 1996), was selected as a candidate gene. Another reason was that our mutant had the phenotype of outheld and elevated wing which maybe resulted from the defect of flight muscles during muscle development (Fernandes et al., 1991; Kozopas and Nusse, 2002). The cytogenetic map of twist gene is 59C2-59C2. In the process of deficiency mapping, we annotated whether all the deficiency stocks completely deleted or disrupted the gene. The results showed that some deficiency strains not deleting or disrupting twist gene could be fail to complement the mutant (mutant phenotype), while the strains that deleting or disrupting twist gene could complement the mutant (wild-type phenotype) (Data not shown). Therefore, we could get the conclusion that the function of twist was not disrupted in the mutant strain and the mutant gene was not a new allele of twist, which was confirmed by sequencing and comparing the gene sequence between the mutant strain and wild-type strain. Now, our main study then focused on the mRNA expression level of this gene. The results showed that the expression of twist gene was increased by 3.6- and 4.5-fold at age of third larval instar and 0-36 h APF, respectively, compared to wild-type at the two stages (data not shown). Therefore, we guessed high-level expression of twist at pupal stage was the main reason for the mutant phenotype.
As for the phenotype of outheld wing, many strains have been screened in fly research history by now. Flap wing (flp), located in 9B14-9C1 interval on the X chromosome, affects indirect flight muscles, which makes the wings hold out in the lateral position. The decapentaplegic gene (dpp), is also reported to relate with the phenotype of outheld wing in 1938 (Novitski and Rifenburgh, 1938). The dpp, encoded a growth factor-like protein (Padgett et al., 1987), is required for dorsal-ventral patterning of the Drosophila embryo (Irish and Gelbart, 1987). Another gene, Dichaete (D), located in 70D3 on the left arm of the third chromosome, got its name from a dominant phenotype discovered by Calvin Bridges in 1914 (Bridges and Morgan, 1923). Dichaete gene has 55 alleles, D1, D3, D5, D6, D7 among which show outheld wings, having the similar phenotype to bsw. The Dichaete protein is a member of the group B Sox class that is expressed in the neuroectoderm from the earliest stages of neurogenesis (Uwanogho et al., 1995; Collignon et al., 1996; Rex et al., 1997; Wood and Episkopou, 1999), closely related to vertebrate Sox1 and Sox2 (Pevny et al., 1998). Genetic analysis has uncovered requirement for Dichaete in segmentation, CNS midline and brain development and hindgut morphogenesis (Russell et al., 1998; Sanchez-Soriano and Russell, 1998, 2000). Here, we enumerate three examples above the outheld wing, which show that the interesting phenotype of outheld wing has a deep relationship with the pathway of wing development or flight muscles and nervous system development. Therefore, the mutant maybe give us some potential cues on the wing or muscle development. The twist gene plays an important role in the development of Drosophila, especially muscular development, so this mutant may be a model to research in the muscular development or muscle-neuron interaction.
CONCLUSION
Comparing the two approaches, P element mapping and the strategy that combining P element rough mapping and SNP mapping together, the latter has many advantages over the former, which prove the latter strategy is a low-cost, efficient and high-resolution method. We know that the reverse genetics plays an important role in the scientific research, however, as for a given phenotype we are interested in, identifying the mutant gene and positional cloning is a key step for the next work as it always a rate-limiting step in the forward genetic analysis. Therefore, both genetic and molecular tools are required to decipher genomic information and ultimately to understand gene function. With the rapid emergence of sequence data from the BDGP, it is important to find efficient methods to correlate sequences with gene functions (Chen et al., 1998). Here, P element mapping and SNP mapping provided a means for fast, inexpensive genetic mapping using existing laboratory strains of Drosophila melanogaster, which will be helpful and offer a rapid strategy to map a mutant of interested phenotype.
ACKNOWLEDGMENTS
We appreciate the assistance of Prof. Zhaohui Wang and Prof. Yongqing Zhang at the Key laboratory of Molecular and Developmental Biology, Institute of Genetics and Developmental Biology, Chinese Academy of Science (CAS), Beijing. We are grateful to Dr. Meiying Fang and Haigang Bao at College of Animal Genetics and Breeding in China Agricultural University (CAU), Beijing. We also thank Lihua Bai, Hongwei Li, Xinhai Li and Qin Wang in CAU for extensive help and excellent technical support.
REFERENCES
- Adams, M.D., S.E. Celniker, R.A. Holt, C.A. Evans and J.D. Gocayne et al., 2000. The genome sequence of Drosophila melanogaster. Science, 287: 2185-2195.
PubMedDirect Link - Baird, N.A., P.D. Etter, T.S. Atwood, M.C. Currey and A.L. Shiver et al., 2008. Rapid SNP discovery and genetic mapping using sequencing RAD markers. PLoS ONE, 3: e3376-e3376.
Direct Link - Bate, M., E. Rushton and D.A. Currie, 1991. Cells with persistent Twist expression are the embryonic precursors of adult muscles in Drosophila. Development, 113: 79-89.
PubMedDirect Link - Baylies, M.K. and M. Bate, 1996. Twist: A myogenic switch in Drosophila. Science, 272: 1481-1484.
PubMedDirect Link - Berger, J., T. Suzuki, K.A. Senti, J. Stubbs, G. Schaffner and B.J. Dickson, 2001. Genetic mapping with SNP markers in Drosophila. Nat. Genet., 29: 475-481.
Direct Link - Chen, B., T. Chu, E. Harms, J.P. Gergen and S. Strickland, 1998. Mapping of Drosophila mutation using site-specific male recombination. Genetics, 149: 157-163.
Direct Link - Collignon, J., S. Sockanathan, A. Hacker, M. Cohen-Tannoudji and D. Norriss et al., 1996. A comparison of the properties of Sox-3 with Sry and two related genes, Sox-1 and Sox-2. Development, 122: 509-520.
Direct Link - Duttaroy, A., 2002. Asymmetric exchange is associated with P element induced male recombination in Drosophila melanogaster. Heredity, 89: 114-119.
PubMedDirect Link - Fernandes, J., M. Bate and K. Vijayraghavan, 1991. Development of the indirect flight muscles of Drosophila. Development, 113: 67-77.
Direct Link - Hoskins, R.A., A.C. Phan, M. Naeemuddin, F.A. Mapa and D.A. Ruddy et al., 2001. Single nucleotide polymorphism markers for genetic mapping in Drosophila melanogaster. Genome Res., 11: 1100-1113.
Direct Link - Irish, V.F. and W.M. Gelbart, 1987. The decapentaplegic gene is required for dorsal-ventral patterning of the Drosophila embryo. Genes Dev., 1: 868-879.
PubMedDirect Link - Johnson, G.C., L. Esposito, B.J. Barratt, A.N. Smith and J. Heward et al., 2001. Haplotype tagging for the identification of common disease genes. Nat. Genet., 29: 233-237.
CrossRefDirect Link - Kozopas, K.M. and R. Nusse, 2002. Direct flight muscles in Drosophila develop from cells with characteristics of founders and depend on DWnt-2 for their correct patterning. Dev. Biol., 243: 312-325.
PubMedDirect Link - Littleton, J.T. and H.J. Bellen, 1994. Genetic and phenotypic analysis of thirteen essential genes in cytological interval 22F1-2: 23B1-2 reveals novel genes required for neural development in Drosophila. Genetics, 138: 111-123.
PubMedDirect Link - Macdonald, S.J., T. Pastinen, A. Genissel, T.W. Cornforth and A.D. Long, 2005. A low-cost open-source SNP genotyping platform for association mapping applications. Genome Biol., 6: R105-R105.
CrossRefDirect Link - Martin, S.G., K.C. Dobi and S.D.A. Johnston, 2001. A rapid method to map mutations in Drosophila. Genome Biol., 2: research0036.1-research0036.12.
Direct Link - Mondal, K., A.G. Dastidar, G. Singh, S. Madhusudhanan, S.L. Gande, K. VijayRaghavan and R. Varadarajan, 2007. Design and isolation of temperature-sensitive mutants of Gal4 in yeast and Drosophila. J. Mol. Biol., 370: 939-950.
CrossRef - Nairz, K., H. Stocker, B. Schindelholz and E. Hafen, 2002. High-resolution SNP mapping by denaturing HPLC. Proc. Natl. Acad. Sci. USA., 99: 10575-10580.
Direct Link - O`Hare, K. and G.M. Rubin, 1983. Structures of P transposable elements and their sites of insertion and excision in the Drosophila melanogaster genome. Cell, 34: 25-35.
PubMedDirect Link - Osamu, S., I. Hiroyuki, Y. Yasuhide, K. Yasuko, H. Reina and Y. Masamitsu, 2009. Identification of the Drosophila Mes4 gene as a novel target of the transcription factor DREF. Exp. Cell Res., 315: 1403-1414.
PubMedDirect Link - Padgett, R.W., S.R.D. Johnston and W.M. Gelbart, 1987. A transcript from a Drosophila pattern gene predicts a protein homologous to the transforming growth factor- gene family. Nature, 325: 81-84.
PubMedDirect Link - Pevny, L.H., S. Sockanathan, M. Placzek and R. Lovell-Badge, 1998. A role for SOX1 in neural determination. Development, 125: 1967-1978.
Direct Link - Rex, M., A. Orme, D. Uwanogho, K. Tointon, P.M. Wigmore, P.T. Sharpe and P.J. Scotting, 1997. Dynamic expression of chicken Sox-2 and Sox-3 genes in ectoderm induced to form neural tissue. Dev. Dyn., 209: 323-332.
PubMedDirect Link - Rubin, G.M., M.G. Kidwell and P.M. Bingham, 1982. The molecular basis of P-M hybrid dysgenesis: The nature of induced mutation. Cell, 29: 987-994.
PubMedDirect Link - Sanchez-Soriano, N. and S. Russell, 1998. The Drosophila SOX-domain protein Dichaete is required for the development of the central nervous system midline. Development, 125: 3989-3998.
Direct Link - Sanchez-Soriano, N. and S. Russell, 2000. Regulatory mutations of the Drosophila Sox gene Dichaete reveal new functions in embryonic brain and hindgut development. Dev. Biol., 220: 307-321.
PubMedDirect Link - Sanyal, S., 2009. Genomic mapping and expression patterns of C380, OK6 and D42 enhancer trap lines in the larval nervous system of Drosophila. Gene Expression Patterns, 9: 371-380.
CrossRef - Theodosiou, N.A. and T. Xu, 1998. Use of FLP/FRT system to study Drosophila development. Method, 14: 355-365.
PubMedDirect Link - Underhill, P.A., L. Jin, A.A. Lin, S.Q. Mehdi and T. Jenkins et al., 1997. Detection of numerous Y chromosome biallelic polymorphisms by denaturing high-performance liquid chromatography. Genome. Res., 7: 996-1005.
Direct Link - Uwanogho, D., M. Rex, E.J. Cartwright, G. Pearl, C. Healy, P.J. Scotting and P.T. Sharpe, 1995. Embryonic expression of the chicken Sox2, Sox3 and Sox11 genes suggests an interactive role in neuronal development. Mech. Dev., 49: 23-36.
PubMedDirect Link - Wang, D.G., J.B. Fan, C.J. Siao, A. Berno and P. Young et al., 1998. Large-scale identification, mapping and genotyping of single-nucleotide polymorphisms in the human genome. Science, 280: 1077-1082.
PubMedDirect Link - Wang, Y.S. and J.A. Todd, 2003. The usefulness of different density SNP maps for disease association studies of common variants. Hum. Mol. Genet., 12: 3145-3149.
Direct Link - Wood, H.B. and V. Episkopou, 1999. Comparative expression of the mouse Sox1,Sox2 and Sox3 genes from pre-gastrulation to early somite stages. Mechanisms Dev., 86: 197-201.
CrossRef - Zhai, R.G., P.R. Hiesinger, T.W. Koh, P. Verstreken and K.L. Schulze et al., 2003. Mapping Drosophila mutations with molecularly defined P element insertions. Proc. Natl. Acad. Sci. USA., 100: 10860-10865.
Direct Link - Zraly, C.B., Y. Feng and A.K. Dingwall, 2002. Genetic and molecular analysis of region 88E9: 88F2 in Drosophila melanogaster, including the ear gene related to human factors involved in lineage-specific leukemias. Genetics, 160: 1051-1065.
Direct Link